Нефтяное загрязнение Каспийского моря как один из факторов инициирования оксидативного стресса у осетровых

Автор: Гераскин Птр Петрович, Пономарева Елена Николаевна, Металлов Геннадий Фдорович, Галактионова Майя Львовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Проблемы прикладной экологии
Статья в выпуске: 5-1 т.14, 2012 года.
Бесплатный доступ
На основании многолетних материалов проанализированы зависимости интенсивности перекисного окисления липидов мышц и печени от уровня загрязнения вод Северного Каспия различными видами токсических веществ и их влияние на активность цитохромоксидазы - фермента, контролирующего интенсивность тканевого дыхания.
Пол, регрессионный анализ, активность цитохромоксидазы, поллютанты
Короткий адрес: https://sciup.org/148201449
IDR: 148201449 | УДК: 597.442:628.394.12
Petrolium pollution of Caspain Sea as one of the factors initiated the oxidative stress in sturgeon
The dependence of rate of lipid peroxidation in muscles and liver from the water pollution by different pollutants in North Caspian Sea was analyzed on the basis of long term materials. It is also shown their influence on the cytochrome-oxidase activity – the enzyme controlling the rate of tissue respiration.
Текст научной статьи Нефтяное загрязнение Каспийского моря как один из факторов инициирования оксидативного стресса у осетровых
активность реакции переокисления липидов у карпа, карася и линя [17]. Изучение воздействия пестицидов группы синтетических пиретроидов на различные виды промысловых рыб Азовского моря выявило у них интенсификацию свободнорадикальных процессов в крови [10]. Двухнедельный контакт со стоками целлюлознобумажного производства 2-х видов Oreochromis niloticus приводил к повышению уровня перекисей липидов в печени, как и у сомиков ( Genidens genidens ), выловленных в загрязненном участке моря [20]. Такой же эффект вызывало воздействие нефти на сеголеток русского осетра в концентрациях 10-20 ПДК [7]. Замечена также активизация процессов ПОЛ у карпа при повышенных концентрациях аммиака в водной среде [2].
Таким образом, увеличение интенсивности перекисных процессов в организме рыб и, прежде всего, в печени, возникает под воздействием разных токсических веществ. Присутствие в водах Каспия большого разнообразия токсикантов затрудняет задачу выделения из них веществ вызывающих у осетровых рыб оксидативный стресс. Однако применение методов регрессионного анализа даёт такую возможность -выделить из всего разнообразия, ведущие факторы.
МАТЕРИАЛ И МЕТОДИКА
Для изучения оксидативного стресса у осетровых рыб исследовалась интенсивность перекисного окисления липидов (ПОЛ) у русского осетра, севрюги и белуги в печени и белых мышцах по реакции малонового диальдегида (МДА) с тиобарби-туровой кислотой, выделяя спонтанную (СпПОЛ) и аскорбатзависимую (АсПОЛ) составляющие, а также накопление МДА в исследуемых тканях [15]. Пробы печени и белых мышц у рыб отбирали сразу же после их вылова тралом. Отлов осетровых рыб вели весной, летом и осенью в Северном Каспии. Анализ проводили по среднегодовым величинам многолетних материалов, полученных в 1993-2003 гг., сравнивая их с данными по среднегодовому содержанию в Северном Каспии различных токсических веществ, любезно предоставленными сотрудниками лаборатории водных проблем и токсикологии КаспНИРХ. Активность ферментов энергетического обмена – цитохромоксидазы и лактатдегидрогеназы определяли также в печени и мышцах. Циохромоксидазы – по скорости окисления восстановленного цитохрома С, лактатдегидрогеназы – окисления лактата в пируват [5, 13].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Окислительный стресс у животных возникает в результате нарушения равновесия в системе прооксиданты-антиоксиданты в сторону накопления молекул с прооксидантными свойствами. Разнообразные по своей природе внешние раздражители, при достаточной силе и продолжительности действия, вызывая активизацию перекисного окисления липидов [12], могут сдвинуть это равновесие и вызвать окислительный стресс.
У разновозрастных особей русского осетра, севрюги и белуги, выловленных в Северном Каспии, также зафиксирована высокая интенсивность пероксидации липидов в печени и мышцах. В одних случаях она была очень высокой, в других опускалась до относительно низких величин. Такие колебания отмечены как для спонтанной реакции переокисления липидов (СпПОЛ), так и протекающей по аскорбатзависимому типу (АсПОЛ), а также в накоплении продукта этой реакции – малонового диальдегида (МДА).
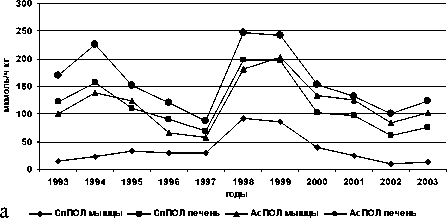
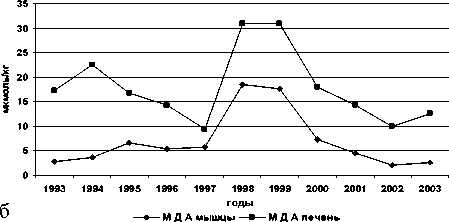
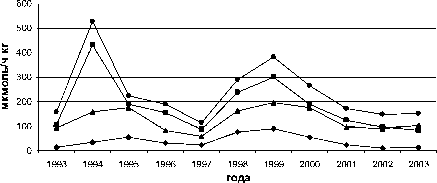
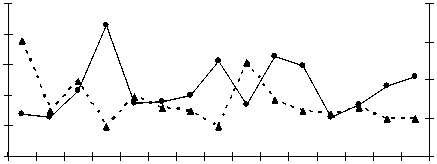
Межгодовые изменения интенсивности обоих типов реакций переокисления и накопления МДА в печени и мышцах у всех исследованных видов однотипны, при имеющихся видовых особенностях (рис. 1).
—♦— СпПОЛ мышцы —■— СпПОЛ печень A АсПОЛ мышцы —•— АсПОЛ печень
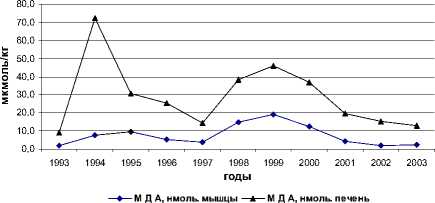
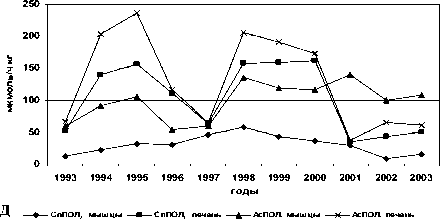
Рис. 1. Межгодовая динамика интенсивности перекисных процессов (АсПОЛ и СпПОЛ) и накопления малонового диальдегида в печени и мышцах русского осетра (а, б), севрюги (в, г) и белуги (д, е)
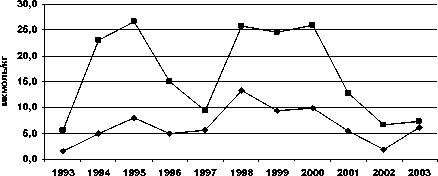
годы е —♦— М Д А. мышцы —■— М Д А. печень
В динамике межгодовых изменений интенсивности перекисных процессов, как в печени, так и мышцах русского осетра, севрюги и белуги четко прослеживаются два пика. Первый пик приходится на 1994 г., а у белуги он пролонгируется и на 1995 г., второй – на 1998-1999 гг. При этом хорошо просматриваются видовые особенности исследованных рыб, выражающиеся в интенсивности роста изучаемых параметров ПОЛ в эти годы. Такие резкие изменения в межгодовых колебаниях параметров перекисного окисления липидов возможны лишь при появлении во внешней среде факторов, воздействия которых на организм рыб носят чрезвычайный характер. Это высокий уровень загрязнения морских вод полютантами, органического и неорганического характера.
Сопоставление межгодовой динамики уровня ПОЛ в печени у исследованных видов осетровых, с колебаниями содержания нефтяных углеводородов в воде Северного Каспии в этот период, выявило достаточно высокое их сходство. Естественно в условиях большого водоёма и высокой миграционной способности осетровых и их биологических особенностей эти совпадения не были идеальными. Однако общие черты динамики изменений совпадали. Минимальному содержанию в воде этих веществ, в 1997 г. соответствовал и минимальный уровень ПОЛ у всех трех (русский осетр, севрюга, белуга) видов осетровых. В тоже время рост концентрации нефтеуглеводородов в воде в 1998 г. в 4,3 раза, по сравнению с 1987 г. [8], вызвал соответственно увеличение интенсивности ПОЛ, как в печени, так и в мышцах. То же самое наблюдалось и 1994 г. – увеличение содержания нефтяных углеводородов в водах Северного Каспия повлекло за собой и подъем скоростей ПОЛ в исследуемых тканях осетровых. Соответствие и направленность изменений концентрации в воде нефтяных углеводородов и уровня ПОЛ отмечается и в 1995 и 1996 гг.: больше НУ – выше ПОЛ в 1995 г., в сравнении с 1996 г. Заметим при этом, что повышенные концентрации нефтеуглеродов в период 1999 г. по 2002 г. [9, 14] отразились и на динамике параметров ПОЛ в эти же годы. При этом необходимо учесть и то обстоятельство, что в это же время в водах Северного Каспия регистрировались и другие виды токсикантов, как органического, так и неорганического характера: фенолы, СПАВ, ХОП, цинк, медь, свинец, кадмий и марганец.
Для уточнения степени влияния загрязняющих веществ на интенсивность перекисных процессов в мышцах и печени осетровых рыб был применён регрессионный анализ. Зависимость интенсивности перекисных процессов от уровня общего загрязнения вод Северного Каспия оказалась очень высокой: коэффициент множественной корреляции колебался от 0,84 до 1,00 (табл. 1).
Таблица 1. Уравнения множественной регрессии параметров перекисного окисления липидов тканей русского осетра от уровня загрязнения различными видами токсикантов
|
Показатели |
R |
R 2 |
R 2* |
p |
Уравнения множественной регрессии |
|
|
X
|
АсПОЛ мкмоль/ч на кг |
0,844 |
0,713 |
-1,87 |
> 0,05 |
статистически неподтверждаемая зависимость |
|
СпПОЛ мкмоль/ч на кг |
0,907 |
0,823 |
-0,77 |
> 0,05 |
статистически неподтверждаемая зависимость |
|
|
МДА мкмоль/кг |
0,909 |
0,826 |
-0,74 |
> 0,05 |
статистически неподтверждаемая зависимость |
|
|
3 G" 3 2 |
АсПОЛ мкмоль/ч на кг |
0,934 |
0,872 |
-0,28 |
> 0,05 |
статистически неподтверждаемая зависимость |
|
СпПОЛ мкмоль/ч на кг |
1,000 |
1,000 |
1,000 |
< 0,01 |
У= - 0,20Х 1 + 1,37Х 2 + 0,416Х 3 - 0,35Х 4 + 0,44Х 5 – 0,88Х 6 + 0,21Х 7 + 1,05Х 8 + 0,249Х 9 – 31,59 |
|
|
МДА мкмоль/кг |
1,000 |
1,000 |
0,997 |
< 0,05 |
У= - 0,29Х 1 + 1,49Х 2 + 044Х 3 – 0,31Х 4 + 0,088Х 5 – 0,98Х 6 + 0,26Х 7 + 1,07Х 8 + 0,28Х 9 -7,64 |
|
Х1 – Нефтеуглеводороды; Х2 – Фенолы; Х3 – СПАВ; Х4 – ХОП; Х5 – Zn; Х6 – Cu; Х7 – Pb; Х8 – Cd; Х9 - Mn
R – множественный коэффициент корреляции; R 2 – коэффициент детерминации; R 2* - скорректированный коэффициент детерминации; р – уровень значимости
Однако достоверными являются лишь регрессионные линейные уравнения зависимости скорости СпПОЛ в мышечной ткани и накопление в ней МДА от концентраций в воде исследованных токсикантов. В этом случае коэффициент множественной корреляции достигает 1,00, показывая прямую линейную зависимость этих параметров ПОЛ от уровня загрязнения токсическими веществами. Приведенные в таблице модели работают практические в 100% варианте (коэффициент детерминации R2=1,00, как и скорректированный его вариант R2*). Более тесная связь интенсивности спонтанного переокисления липидов и накопления МДА в мышцах объясняется меньшей лабильность данных показателей, в сравнении с параметрами ПОЛ в печени, что даёт возможность им более адекватно реагировать на изменения уровня загрязнения среды обитания.
Более детальный анализ зависимостей исследуемых параметров ПОЛ от концентраций отдельно взятых полютантов показал, что достоверную корреляционная связь из большого набора загрязняющих веществ демонстрируют: содержание в воде нефтеуглеводородов, фенолов, а также меди и кадмия (табл. 2).
В первом случае регрессионные модели, подобранные для отдельных параметров ПОЛ, имеют коэффициенты корреляции от 0,61 до 0,81, во втором – от 0,66 до 0,84. В случае с кадмием они также высоки – 0,69-084. Достоверные модели корреляционных отношений меди и параметров ПОЛ найдены лишь для таких показателей как спонтанное переокисление липидов в мышцах и содержание в них малонового диальдегида при примерно близких коэффициентах корреляции. Если рассмотреть вариант множественной корреляции параметров ПОЛ печени с выявляемыми в воде Северного Каспия токсичными веществами, то достоверный характер обнаружен лишь для регрессионных уравнений, связанных с содержанием в этом органе малонового диальдегида и интенсивностью спонтанного переокисления (табл. 3). В первом случае при варианте с тремя (НУ, фенолы, Cd) и четырьмя (НУ, фенолы, Cd, Cu) токсичными веществами, во втором - только с тремя токсикантами. Из этого следует, что основу негативного влияния загрязнения вод Северного Каспия на русского осетра составляют нефтеуглеводороды, фенолы и кадмий, а также медь.
Таблица 2. Уравнения регрессии параметров перекисного окисления липидов тканей русского осетра от уровня загрязнения различными видами токсикантов
|
Показатели |
Коэффициент |
Уровень |
Уравнения регрессии |
|
значимости |
|||
|
корреляции детерминации |
Нефтеуглеводороды (мг/л)
|
X
|
АсПОЛ |
r = 0,6162 |
R2= 0,38 |
p < 0,05 |
У = 164,240 Х + 42,289 |
|
СпПОЛ |
r = 0,6634 |
R 2 = 0,44 |
p < 0,05 |
У = 142,681 Х + 25,261 |
|
|
МДА |
r = 0,7420 |
R 2 = 0,55 |
p < 0,01 |
У = 24,485 Х + 3,333 |
|
|
3 G" 3 X 2 |
АсПОЛ |
r = 0,6385 |
R 2 = 0,40 |
p < 0,05 |
У = 122,299 Х + 30,740 |
|
СпПОЛ |
r = 0,8099 |
R 2 = 0,66 |
p < 0,01 |
У = 97,423 Х - 4,332 |
|
|
МДА |
r = 0,8005 |
R 2 = 0,64 |
p < 0,01 |
У = 19,618 Х - 0,998 |
Фенолы (мкг/л)
|
G
|
АсПОЛ |
r = 0,7225 |
R 2 = 0,52 |
p < 0,01 |
У = 30,487 X + 15,497 |
|
СпПОЛ |
r = 0,7943 |
R 2 = 0,63 |
p < 0,01 |
У = 28,323Х - 1,530 |
|
|
МДА |
r = 0,8000 |
R 2 = 0,64 |
p < 0,01 |
У = 4,472 Х - 0,4744 |
|
|
3 G" 3 s |
АсПОЛ |
r = 0,6642 |
R 2 = 0,44 |
p < 0,05 |
У = 22,332Х + 12,651 |
|
СпПОЛ |
r = 0,8278 |
R 2 = 0,69 |
p < 0,01 |
У = 17,066Х -18,088 |
|
|
МДА |
r = 0,8373 |
R 2 = 0,70 |
p < 0,001 |
У = 3,597Х - 4,069 |
Медь (мг/л)
|
3 G" |
СпПОЛ |
r = 0,7060 |
R 2 = 0,50 |
p <0,05 |
У = 1,560Х + 2,704 |
|
3 3 2 |
МДА |
r = 0,6684 |
R 2 = 0,45 |
p < 0,05 |
У = 0,282Х + 0,596 |
Кадмий (мг/л)
|
X G G |
АсПОЛ |
r = 0,6870 |
R 2 = 0,47 |
p < 0,05 |
У = 226,405Х + 15,239 |
|
СпПОЛ |
r = 0,7532 |
R 2 = 0,57 |
p < 0,05 |
У = 209,764Х - 1,515 |
|
|
МДА |
r = 0,7746 |
R 2 = 0,68 |
p < 0,01 |
У = 33,870Х - 0,780 |
|
|
3 G" 3 s |
АсПОЛ |
r = 0,8384 |
R 2 = 0,70 |
p < 0,01 |
У= 222,062Х - 6,443 |
|
СпПОЛ |
r = 0,8067 |
R 2 = 0,65 |
p < 0,01 |
У = 130,623Х - 19,963 |
|
|
МДА |
r = 0,8216 |
R 2 = 0,60 |
p < 0,01 |
У = 27,689Х - 4,488 |
АсПОЛ, СпПОЛ – мкмоль/ч на кг сырой ткани; МДА – мкмоль/кг сырой ткани
Таблица 3. Уравнения множественной регрессии параметров ПОЛ печени русского осетра от содержания в воде некоторых видов токсических веществ
|
Показатели ПОЛ |
R |
R2 |
R2* |
p |
Уравнения множественной регрессии |
|
СпПОЛ мкмоль/ч на кг |
0,84 |
0,71 |
0,58 |
< 0,05 |
У= -0,12Х 1 + 0,85Х 2 + 0,59Х 8 – 14,12 |
|
МДА |
0,88 |
0,77 |
0,62 |
< 0,05 |
У= - 0,08Х 1 + 0,96Х 2 + 0,40Х 8 – 3,51 |
|
мкмоль/кг |
0,88 |
0,77 |
0,62 |
< 0,05 |
У= - 0,08Х 1 + 0,94Х 2 + 0,073Х 6 + 0,34Х 8 – 3,70 |
Х1 - Нефтеуглеводороды; Х 2 - Фенолы; Х 6 - Cu; Х 8 - Cd ;
R – множественный коэффициент корреляции; R 2 – коэффициент детерминации;
R 2* - скорректированный коэффициент детерминации; р – уровень значимости
Анализ в этом отношении параметров ПОЛ двух других видов осетровых дал результаты, свидетельствующие о видовых особенностях реакции этих рыб на загрязнение среды обитания. Так, у белуги достоверная множественная корреляция обнаружена с изученными компонентами загрязнения вод северного Каспия только для скоростей аскорбатзависимого переокисления липидов в мышцах, несмотря на высокие значения корреляционных отношений (от 0,94 до 1,00) по другим параметрам ПОЛ (табл. 4).
В отличие от русского осетра, у белуги выявлены лишь регрессионные зависимости (табл. 5) для двух пар: НУ : МДА (мышцы) и Zn : АсПОЛ (мышцы).
У севрюги регрессионный анализ даёт достоверные результаты лишь в случае уменьшения видового количества токсикантов, участвующих в моделировании зависимости перекисных процессов от содержания полютантов в воде (табл. 6).
Таблица 4. Уравнение множественной регрессии интенсивности АсПОЛ печени белуги от уровня загрязнения различными видами токсикантов
|
Показатели ПОЛ |
R |
R2 |
R2* |
p |
Уравнение множественной регрессии |
|
АсПОЛ мкмоль/ч на кг |
1,0 0 |
1,00 |
1,00 |
<0,05 |
У=0,30Х 1 +0,35Х 2 +0,09Х 3 –0,26Х 4 +0,63Х 5 -0,78Х 6 -0,02Х 7 +0,85Х 8 - 0,21Х 9 +23,63 |
Х1 – Нефтеуглеводороды; Х2 – Фенолы; Х3 – СПАВ; Х4 – ХОП; Х5 – Zn; Х6 – Cu; Х7 – Pb; Х8 – Cd; Х9 - Mn.
R – множественный коэффициент корреляции; R 2 – коэффициент детерминации; R 2* - скорректированный коэффициент детерминации; р – уровень значимости
Таблица 5. Уравнения регрессии некоторых показателей ПОЛ в мышцах белуги от уровня загрязнения различными видами токсикантов
Таблица 6. Уравнение множественной регрессии параметров ПОЛ печени и мышц севрюги от уровня загрязнения различными видами токсикантов
|
Показатели |
R |
R 2 |
R 2* |
p |
Уравнение множественной регрессии |
|
|
3 3 S |
СпПОЛ мкмоль/ч на кг |
1,00 |
1,00 |
1,00 |
<0,05 |
У=0,32Х 1 +0,52Х 2 -0,24Х 3 +0,25Х 6 +0,53Х 9 -56,84 |
|
МДА мкмоль/кг |
1,00 |
1,00 |
0,99 |
<0,001 |
У=0,83Х 1 -0,24Х 3 +0,64Х 9 |
|
|
X К |
СпПОЛ мкмоль/ч на к |
1,00 |
1,00 |
1,00 |
<0,01 |
У=0,85Х 1 +0,06Х 2 -0,09Х 7 +0,15Х 8 +0,29Х 9 -38,04 |
|
АсПОЛ мкмоль/ч на кг |
1,00 |
1,00 |
1,00 |
<0,01 |
У=1,10Х 1 -0,89Х 6 +0,45Х 7 +0,51Х 8 +0,52Х 9 -8,30 |
|
|
МДА мкмоль/кг |
1,00 |
1,00 |
1,00 |
<0,01 |
У=0,78Х 1 +0,07Х 4 +0,14Х 6 +0,25Х 8 +0,30Х 9 -10,15 |
Х1 – Нефтеуглеводороды; Х2– Фенолы; Х3 – СПАВ; Х4 – ХОП; Х6 – Cu; Х7 – Pb; Х8 – Cd;
Х9 – Mn
R – множественный коэффициент корреляции; R 2 – коэффициент детерминации;
R 2* – скорректированный коэффициент детерминации; р – уровень значимости
Однако и в этом случае во всех вариантах регрессионного анализа, достоверно отражающих влияние компонентного состава загрязнения вод Северного Каспия, на первом месте присутствуют нефтеуглеводороды и лишь как дополнение к ним тяжелые металлы и один или два вещества органического происхождения.
Для мышечной ткани найдены модели, отражающие влияние на интенсивность спонтанного переокисления липидов комплекса нефтеуглеводороды, фенолы, СПАВ, медь и марганец, а малонового диальдегида – регрессионное уравнение, элементами которого являются содержание в воде нефтеуглеводородов, СПАВ и марганца.
В печени интенсивность обоих типов реакций моделируют регрессионные уравнения, в состав которых входят: для спонтанной реакции – нефтеуглеводороды, фенолы и три тяжёлых металла (свинец, кадмий и марганец), а для аскорбатзависимой – нефтеуглеводороды и 4 металла (медь, свинец, кадмий и марганец). Накопление в печени МДА в наибольшей степени отражает другой комплекс токсических веществ – нефтеуглеводороды, ХОП и тяжёлые металлы (медь, кадмий, марганец). В этих сочетаниях практически все модели, представленные в таблице 6 работают в 100% варианте (коэффициент детерминации R2 и его скорректированная величина R2* равны 1,00).
В случае регрессионного анализа по отдельным видам токсических веществ, у севрюги достоверные связи выявлены для двух параметров ПОЛ в мышцах: интенсивности спонтанного переокисления липидов и содержания малонового диальдегида, величина которых коррелирует с содержанием в воде фенолов и меди (табл. 7).
Коэффициент корреляции в этом случае не столь высокий – 0,58-0,68, из чего следует, что на показатели ПОЛ у севрюги оказывает влияние не отдельные токсические вещества, а их комплекс, индуцируя состояние окислительного стресса у исследованных рыб. То же самое можно сказать и про другие виды осетровых рыб. Это демонстрирует высокая степень множественной корреляции и коэффициент детерминации, свидетельствующий о полном совпадении величин параметров ПОЛ от уровня загрязнения токсичными веществами вод Северного Каспия, описываемых уравнениями множественной регрессии. При этом отметим, что во всех вариантах множественной регрессии одним из элементов модели выступают нефтеуглеводороды. Вторыми в этом ряду стоят фенолы, а уже затем, различные виды тяжёлых металлов. Видовые особенности выражаются в проявлениях связи между параметрами ПОЛ и отдельными видами токсикантов или их сочетанием. Среди тяжёлых металлов можно выделить кадмий, который, видимо, в сочетании с нефтеуглеводородами и фенолами увеличивает токсичность последних. Это следует из того, что его содержание в воде Северного Каспия составляет сотые доли ПДК, в то время как по меди и цинку отмечаются превышения предельно допустимых концентраций.
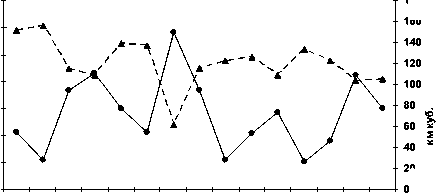
Реагирует на изменения загрязнения вод Северного Каспия и энергетическая система осетровых рыб. Ответ этой физиологической системы на ухудшение экологической ситуации в море проявляется изменениями активностей ферментов аэробного и анаэробного пути получения энергии в печени и мышцах – цитохромоксидазы (ЦО) и лактатдегидрогеназы (ЛДГ). Наиболее чутко на воздействие токсикантов отзывается ЦО печени (рис. 2.).
Таблица 7. Уравнения регрессии некоторых показателей ПОЛ в мышцах белуги от уровня загрязнения различными видами токсикантов
|
Показатели ПОЛ в мышцах |
Токсиканты |
r |
R 2 |
p |
Уравнение регрессии |
|
СпПОЛ |
Фенолы |
0,68 |
0,46 |
<0,05 |
У = 0,017Х + 1,448 |
|
мкмоль/ч на кг |
Медь |
0,58 |
0,33 |
<0,05 |
У = 0,048Х + 4,511 |
|
МДА |
Фенолы |
0,64 |
0,41 |
<0,05 |
У = 0,075Х + 1,540 |
|
мкмоль/кг |
Медь |
0,63 |
0,40 |
<0,05 |
У = 0,244Х + 4,524 |
|
r – коэффициент корреляции; R 2 – коэффициент детерминации; р – уровень значимости |
|||||
4,5
I 3,5
1 2,5
i 1,5
0,5 0
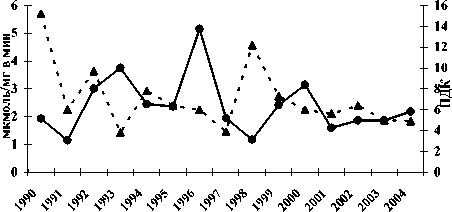
ЦО печень - - *- - ∑ПДК НУ+Фен
а
б
ЦО печень - - - ∑ПДК НУ+Фен
s
4 i
s
i
10 I
1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004
годы
ЦО печень - - -* - - ■ ∑ПДК НУ+Фен
в
Рис. 2. Межгодовая динамика активности цитохромоксидазы в печени русского осетра (а), севрюги (б), белуги (в) и суммарного уровня загрязнения нефтеуглеводородами и фенолами
Вследствие этого её активность существенно колеблется на протяжении исследуемого периода. Как было показано выше, на примере межгодовой динамики интенсивности перекисных процессов, организм осетровых в большей степени реагирует на те загрязнения, уровень которых в воде превышает рыбохозяйственные ПДК, а именно нефтепродукты и фенолы. В годы роста суммарной загрязнённости этими токсикантами водной среды
Северного Каспия в большинстве случаев имеет место снижение активности ЦО в печени – как в 1994 и 1998 гг., при однозначном повышении активности фермента в годы снижения загрязнённости водной среды, как в 1993 и 19961997 гг. Так, у русского осетра и севрюги в 1994 г. среднегодовая величина активности ЦО в печени снижалась соответственно до 2,47±0,26 и 2,45±0,3 мкмоль/мг в мин, при ещё большем понижении в
1998 г. – до 1,02±0,18 и 1,18±0,33 мкмоль/мг в мин соответственно.
У белуги активность этого фермента в печени в эти годы была ещё ниже и не различалась по величине – 1,7±0,14 мкмоль/мг в мин. В то же время 1993 г. она составляла у осетра и севрюги 3,75 мкмоль/мг в мин при ошибке ±0,37 и ±0,11 соответственно, а в 1996 г. ещё выше – 4,73±0,48 и 5,15±0,26 мкмоль/мг в мин. У белуги среднегодовая активность цитохромоксидазы в 1993 г. была также достаточно высокой – 4,29±0,11
мкмоль/мг в мин, как и в 1997 г. – 3,09±0,39.
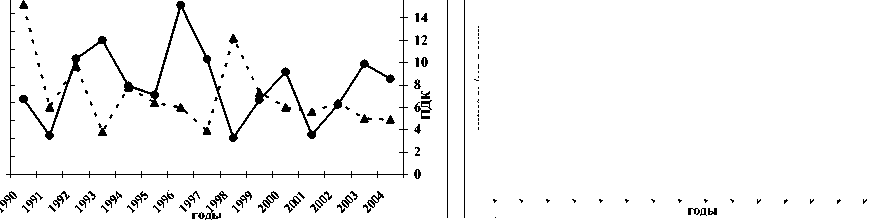
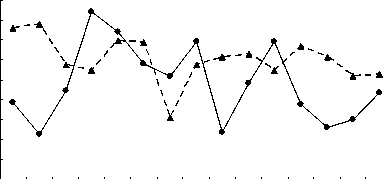
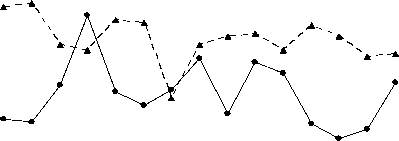
Однако если учесть, что активность энергетических ферментов определялась в летний период, то точнее вести сравнительный анализ не со среднегодовым загрязнением вод Северного Каспия, а уровнем загрязнённости в летний период обусловленного выносом различных токсических веществ паводком. Считается [1], что основной объём (около 90%) загрязняющих веществ в Северный Каспий поступает с речным стоком, при сохранении примерного соотношения загрязняющих веществ. Это иллюстрирует рисунок 3, на котором прослеживается обратная зависимость степени активности цитохромоксидазы в печени в летний период от величины паводка. Чем больше вынос волжскими водами токсических веществ в Северный Каспий, тем ниже активность в печени цитохромоксидазы.
1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004
годы
—• ЦО печень - -*- - Паводок
4,5
4,0
3,5
3,0
2,5
2,0
1,5
1,0
0,5
0,0
1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004
годы
—е— ЦО печень - - Паводок
а
б
2 2
60 1
годы
ЦО печень —*— Паводок
1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004
в
Рис. 3. Межгодовая динамика активности ЦО печени русского осетра (а), севрюги (б) и белуги (в) в летний период при различных объёмах паводка
Таким образом, индуцирование оксидативного стресса со снижением тканевого дыхания у русского осетра, севрюги и белуги, нагуливающихся в Северном Каспии, возникает в летний период вследствие выноса половодьем токсических веществ. Его уровень напрямую зависит от общего загрязнения вод. Однако основу негативного влияния на исследованные виды осетровых составляют нефтеуглеводороды, фенолы и кадмий, усиливающий, по всей видимости, токсичность первых двух, а также, в некоторой степени, медь.
Список литературы Нефтяное загрязнение Каспийского моря как один из факторов инициирования оксидативного стресса у осетровых
- Глумов И.Ф. Современное состояние экосистем и хозяйственное освоение природных ресурсов Каспийского моря.//Бюллетень «Использование и охрана природных ресурсов в России. 2001. № 7. С. 64-70.
- Грубинко В.В., Леус Ю.В., Арсан О.М. Перекисное окисление липидов в тканях карпа при действии аммиака//Гидробиол. ж. 1996. 32, № 4. С. 52-57.
- Грубинко В.В., Смольский О.С., Леус Ю.В., Явоненко О.Ф., Арсан О.М. Взаимосвязь функционирования системы гемоглобина и перекисного окисления липидов в крови карпа при интоксикации.//Док. Нац. АН Украины. 1997. № 2. С. 146-150.
- Гумидова Г.П., Сорокина Н.Н. Некоторые условия спектрофотометрического определения активности СДГ и ЦХО в митохондриях мозга.//Бюлл. эксперимент. биол. и медиц. 1967, 1. С.41.
- Гумидова Г.П., Сорокина Н.Н. Некоторые условия спектрофотометрического определения активности СДГ и ЦХО в митоходриях мозга. Бюлл.эксперимент. биол. и медиц. 1967. с. 41.
- Есина О.И. Действие органических соединений олова на молодь осетровых рыб.//Автореф. дис. канд. биол. наук. Астрахань. 2006. 23с.
- Исуев А.Р., Габибов М.М., Гусейнова С.А. Состояние перекисного окисления липидов и антиокислительной активности сеголеток русского осетра Acipenser gueldenstaedtii как показатель загрязнения нефтью окружающей водной среды///Вопр. ихтиол. 2000. 40, № 4. С. 551-559.
- Катунин Д.Н., Гераскин П.П., Курочкина Т.Ф., Костров Б.Л., Андреев В.В. Современное углеводородное загрязнение Каспийского моря и возможные последствия для экосистемы при широкомасштабном развертывании нефтедобычи на шельфе.//Вестник МАНЭБ. С.-Пб. 1999. № 9 (21) С. 33-39.
- Катунин Д.Н., Курочкина Т.Ф., Попова О.В., Рылина О.Н., Карыгина Н.В., Теркулова А.А. Содержание загрязняющих веществ в водоёмах Волго-Каспийского бассейна.//Рыбохозяйственные исследования на Каспии. Астрахань. 2002. С. 37-41.
- Кесельман М.Л., Гвозденко С.И., Кузнецов Л.Я. Свободнорадикальные процессы, активность антиоксидантных механизмов и структурно-функциональное состояние мембран в тканях рыб при воздействии пиретроидных пестицидов//В сб.: 1 Конгр. ихтиологов России. Астрахань. сент. 1997. Тез. докл. Астрахань. 1997. С. 220-221.
- Лукьяненко В.И. Общая ихтиотоксикология. М.: Легкая и пищевая промышленность. 1983. 320 с.
- Меерсон Ф.З. Адаптация, стресс и профилактика. М.: Наука. 1981. 278 с.
- Методы биологии развития. М.: Наука. 1974. С. 323-363.
- Рылина О.Н., Попова О.В., Попов О.П., Карыгина Н.В., Теркулова А.А., Чуйко Е.В., Кобзева Л.П., Попова Е.С. Эколого-токсикологический мониторинг Волго-Каспийского бассейна//В сб.: Рыбохозяйственные исследования на Каспии. Астрахань. 2003. С. 54-74.
- Строев Е.А., Макарова В.Г. Практикум по биологической химии. М.: Высшая школа, 1986. С. 211-214.
- Bano Y. and Hasan M. Mercury induced time-dependent alteration in lipid profiles and lipid peroxidation in different body organs of catfish Heteropneustes Fossilis.//J. Environ. Sci. Health. 1989. B24. P.145-166.
- Gabryelak T. and Klekot J. The effekt of paraquat on the peroxide metabolism enzymes in erythrocytes of freshwater fish species.//Comp. Biochem. Physiol. 1985, B 77. Р. 583-587.
- Radi A.A.R. and Matkovics B. Effekts of metal ions on the antiokxidant enzyme activities, protein contents and lipid peroxidation of carp tissues.//Comp. Biochem. Physiol. 1988. № 90. P. 69-72.
- Thomas P, Wofford H.W. and Neff J.M. Effekt of cadmium on glutatione content of mullet (Mugil cephalus) tissues. In Physiological Mechanisms of Marine Pollutant Toxicity (Edited by Vernberg W. B., Calabrese A., Thurberg F. H. and Vernberg F.P.). Academic Press. New York. 1982. Р. 109-125
- Wilhelm Filho D., Soares C.H.L., Pedrosa R.C., Baptista I.E., Tribess T.B., Gaspari G. The effect of pulp mill effluents on fish antioxidants. II. Final effluents.//Abstr. 9th Bieen. Meet. Int. Soc. Free Radic. Res. “Free Radic. Res. 21st Century”, Sao Paulo, 7-11 Sept., 1998. Rev. farm. e bioquim. Univ. Sao Paulo. 1998. 34, Supl. N 1. Р. 136.
