Нейромедиаторные биогенные амины в структурах костного мозга при аллопересадке

Автор: Воробьева Ольга Васильевна
Журнал: Морфологические ведомости @morpholetter
Рубрика: Краткие сообщения
Статья в выпуске: 4 т.24, 2016 года.
Бесплатный доступ
Изучено влияние аллопересадки костного мозга на биоаминсодержащие структуры костного мозга во временном аспекте. C помощью методов люминесцентной микроскопии и цитоспектрофлюориметрии в костном мозгє дана количественная характеристика содержания гранулярных люминесцирующих и тучных клеток, а также содержание нейроаминов. Установлено, что до 40 мин аллопересадки у мышей наблюдается увеличение числа биоаминпродуцирующих структур (гранулярных люминесцирующих и тучных клеток) в костном мозге. Через 2 часа происходит истощение данных структур, со сниженным содержанием нейроаминов. Разрушается регуляторное звено в костном мозге, осуществляющее контроль за размножением клеток. Нарушаются процессы пролиферации и эпоптоза в костном мозге.
Аллотрансплантация костного мозга, катехоламины, серотонин, гранулярные люминесцирующие клетки, тучные клетки
Короткий адрес: https://sciup.org/143177153
IDR: 143177153
Neurotransmitters biogenic amines in the bone marrow structures at the allografting
The effect of allografting of bone marrow on bioamine - containing structures in the temporal aspect was studied with the use of fluorescent mieroscopy and cytospectrofluorimetry in the bone marrow and quantitative description of the content of granular luminescent and mast cells, as well as cell which content neyroamins. It is found that up to 40 min of allografting observed increase of the number of bioamin containing structures (granular luminescent and mast eells)in the bone marrow. Through 2 hours depletion occurs numbers of this structures and reducing of neyroamins. Is destroyed regulatory link in the bone marrow which exercised control over the reproduction of cells. Violated the processes of proliferation and apoptosis in the bone marrow.
Текст краткого сообщения Нейромедиаторные биогенные амины в структурах костного мозга при аллопересадке
Введение. В организме содержатся биоа- минпродуцирующих клеток, которые оказывают регулирующее действие на клетки и ткани. Среди этих клеток выявляют гранулярные люминесциру-ющие и тучные клетки, содержащие наибольшее количество нейроаминов. Кроме них нейроамины содержатся в адренергических нервных волокнах, мегакариоцитах, липоцитах. Гранулярные люминесцирующие и тучные клетки являются продуцентами нейроаминов. Они способны продуцировать, накапливать и выделять нейроамины [1, 2, 3, 4]. Адренергические нервные волокна содержат катехоламины и серотонин, которые являются результатом синтеза нервных клеток. Липоциты также содержат в большой концентрации нейроамины, но в основном в цитоплазме и их ядре. Данные клетки скорее всего являются адсорбентами и накопителями для этих веществ [2].
В связи с этим, нами в основном изучались данные структуры в костном мозге после аллотрансплантации костного мозга.
Цель исследования - изучение нейроамин-ного обмена в костном мозге при аллопересадке во временном аспекте.
Материал и методы исследования. Опытных животных разделили на 2 серии: 1 - интактные мыши без введения (n=15); 2 - аллогенная пересадка (n=15) – 0,1 мл костного мозга, взятый из бедренной кости мыши помещали в 1 мл 0,9% физиологического раствора и тщательно размешивали. 1 мл суспензии костного мозга вводили в хвостовую вену мыши другой линии. Число клеток в 1 мл суспензии было равно 2,1* 108.
Животные содержались в стандартных условиях вивария со свободным доступом к корму. Мышей выводили из эксперимента через 40 мин и 2 часа после аллопересадки костного мозга, брали костный мозг и изготавливали срезы 5 мкм с последующей обработкой по методу Фалька–Хилларпа в модификации Е.М. Крохиной для выявления катехоламинов и серотонина [5]. Концентрация катехоламинов, серотонина в структурах костного мозга оценивались с помощью ми-крофлуориметрической насадки к люминесцентному микроскопу ФМЭЛ-6 (ЛОМО, Россия). Для определения катехоламинов служил интерферен- ционный фильтр 6 (48О нм), серотонина – 8 (525 нм). Hаблюдение велось при напряжении 8ОО вольт, с диаметром зонда О,5 (условные единицы). Подсчет гранулярных люминесцирующих и тучных клеток производили в 5 полях зрения микроскопа Микромед-2 (ЛОМО, Россия) при увеличении об. 40, ок. 10, диаметр зонда 0,5 мм. Использовался иммуногистохимический метод для выявления антиапоптического белка Bcl2 в структурах костного мозга. Полученные данные обрабатывались с использованием стандартного программного пакета Statistica 6.0 на персональном компьютере Pentium III. Оценку значимости цифровых данных проводили по t-критерию Стьюдента. Различия считали значимыми при P<0,05.
Результаты исследования и их обсуждение. Через 40 минут в гранулярных люминес-цирующих клетках костного мозга происходит снижение содержания катехоламинов до 8,8±0,1 у.е. (у интактных мышей - 15,6±0,1 у.е.) и серотонина до 9,7±0,1 у.е. (у интактных - 14,1± 0,1 у.е.), наблюдается незначительное увеличение данных нейроаминов в межклеточном пространстве. Аналогичная картина отмечается в тучных клетках. со сниженным содержанием катехоламинов до 10,1±0,2 у.е. (у интактных мышей - 19,6±0,2 у.е.) и серотонина до 11,3±0,4 у.е. (у интактных мышей -21,3±0,4 у.е.). Однако число тучных и гранулярных люминесцирующих клеток остается повышенным до 3 – 4 на поле зрения (у интактных – 1-2 клетки).
Выявляются группы клеток, расположенные в центре размножения, состоящие из макрофага, гранулярной люминесцирующей, тучной клетки, в редких случаях можно выявить липоцит. Вокруг этих клеток-регуляторов располагаются клетки эритроидного ряда, или смешанные: эритроидного и нейтрофильного рядов. И макрофаг, и гранулярная люминесцирующая клетка похожи друг на друга, но отличаются содержанием моноаминов и формой гранул. Нервные волокна слабо визуализируются. Тучные клетки чаще всего выявляются частично дегранулироваными.
Через 2 часа люминесценция межклеточного вещества была увеличена вследствие тотальной дегрануляции тучных клеток. Число тучных и гранулярных люминесцирующих клеток снижается до 1 – 2 клеток на несколько полей зрения. Выявляется распад гранул в гранулярных люминесциру-ющих клетках, имеющие 1-2 люминесцирующие гранулы, а остальные не люминесцировали вследствие отсутствия нейроаминов в них. Содержание катехоламинов в люминесцирующих гранулах было повышенным. Гранулярные люминесциру-ющие и тучные клетки располагаются диффузно, в то время, как у интактных мышей данные клетки располагаются около островков размножения. Отмечается сниженное число гранулярных люми- несцирующих и тучных клеток в костном мозге по сравнению с интактными мышами.
Выявляемость адренергических нервных волокон, при исследовании на катехоламины и серотонин, увеличилась, но они не имели четких очертаний, т.е. произошел выброс нейроаминов из этих структур. Выявлялись, в повышенном числе, округлые клетки со светящимися мелкими гранулами и несветящимся бобовидными ядрами (макрофаги).
Кроме того, наблюдается распад групп клеток, расположенных в центре размножения, которые осуществляют синтез и поддерживают оптимальное содержание нейроаминов в костном мозге.
Экспрессия белка Bcl 2 выявляется в тучных клетках, одни из которых имеют темное ядро, в других оно не определяется. Появляются единичные компактные тучные клетки. Позитивных гранулярных клеток выявляется до 3 (у интактных 5-6), иногда образующие скопления.
Кроме того, экспрессия белка Bcl 2 определяется в некоторых ядрах сегментоядерных нейтрофилов и митотически делящихся клетках (рис. 1). Отмечается сниженное число позитивно экспрессирующих белок мегакариоцитов.
Исходя из вышеизложенных данных можно заключить, что аллопересадка костного мозга существенно влияет на содержание нейроаминов в костном мозге и изменяет внутриклеточные связи. Через 40 минут в гранулярных люминесцирующих и тучных клетках костного мозга отмечается численное снижение нейроаминов с последующей частичной дегрануляцией и накоплением их в межклеточном пространстве. Однако функциональная активность биоаминпродуцирующих клеток (гранулярных люминесцирующих, тучных клеток) остается на высоких значениях по отношению к интактным мышам, т.е. продукция и секреция нейроаминов сохраняется. До 40 минут физиологическая тенденция клеток продуцентов: гранулярных люминесцирующих и тучных клеток однонаправленная. Однако уже к 2-м часам от начала эксперимента имеются резкие различия как в расположении этих клеток, так и в содержании в них нейроаминов. Происходит дегрануляция тучных клеток и исчезновение нейроаминов в некоторых гранулах гранулярных люминесциру-ющих клетках. Число нейроаминпродуцирующих клеток снижается, происходит выход нейроаминов в межклеточное пространство из некоторых гранул, результатом чего является нарушение соотношения процессов регенерации гемопоэтических клеток.
Кроме того, в костном мозге выявлено регуляторное звено, состоящее из комплекса клеток – из гранулярной люминесцирующей, тучной клет-
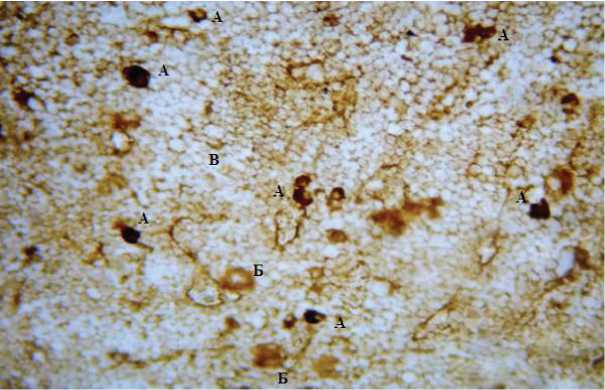
Рис. 1. Срез костного мозга при аллопересадке костного мозга. А. Тучные клетки; Б. Мегакариоциты; В. Клетки эритроидного ряда. Реакция на маркер Bcl 2. Ув. 00 Микроскоп Leica DM 4000B.
ки, ретикулярной клетки и макрофага, которые содержат все изучаемые нейроамины и участвуют в размножении, дифференцировке, старении и умирании клеток. Начиная с ранних сроков после подсадки чужеродного костного мозга, нарушается структурная организация комплекса клеток продуцентов. Гранулярные люминесцирующие и тучные клетки располагаются неупорядоченно, теряют локализацию расположения около гемопоэтических островков. Таким образом, группы гемопоэтических клеток не получают достаточного количества нейроаминов, что приводит к нарушению процессов возобновления клеточных форм костного мозга. Через 2 часа данный комплекс распадается, и изменяется функционирование кроветворной ткани в костном мозге.
При анализе иммуногистохимии выявлено, что до 40 мин появляются единичные компактные тучные клетки. Можно предположить, что это образуются молодые клетки, позволяющие сохранять оптимальное содержание нейроаминов в костном мозге. Через 2 часа данные клетки разрушаются. Уменьшается число позитивных на белок Bcl 2 мегакариоцитов, что указывает о нарушенном тромбоцитопоэзе. Можно предположить, что аллопересадка костного мозга угнетает процессы апоптоза и пролиферации.
Следуя вышеизложенным данным, можно заключить, что аллогенная пересадка костного мозга приводит к численному снижению биоамин-продуцирующих клеток, с нарушенным синтезом нейроаминов и в последующем, к разрушению группы клеток-регуляторов, расположенных около островков размножения.
Выводы:
-
1. При аллопересадке в костном мозге до 40 мин наблюдается увеличение числа нейроа-минпродуцирующих клеток, но в поздние сроки происходит уменьшение данных клеток и выход нейроаминов в межклеточное
-
2. Происходит распад группы клеток, осуществляющих регуляцию содержания нейроаминов в костном мозге.
-
3. Угнетаются процессы пролиферации и апоптоза в костном мозге после аллопересадки.
пространство с последующим распадом.
Список литературы Нейромедиаторные биогенные амины в структурах костного мозга при аллопересадке
- Агафонкин, С.А. Исследование биогенных аминов и биаминосодержащих структурах костного мозга человека при нарушении гемопоэза/С.А. Агафонкин//автореф.дис..канд.мед.наук. -М., 2006. -25 с.
- Любовцева, Л.А., Любовцева, Е. В. Биоаминсодержащие структуры костного мозга при системных заболеваниях крови/Л.А. Любовцева, Е.В. Любовцева//Морфология. -2012. -№3. -С. 95-96.
- Ставинская, О.А. Роль гистамина и серотонина в поддержании иммунного гомеостаза/О.А. Ставинская//Национальная конференция «Аллергология и клиническая иммунология -междисциплинарные проблемы». -российский аллергологический журнал. -Москва. -2008. -№1. -С. 283-284.
- Boyce, J. A. The biology of the mast cell/J.A. Boyce//Allergy and Asthma Proc. 2004. -Vol. 25, №1. -P.27-30.
- Falck, B. Fluorescence of catecholamines and related compounds condensed with formaldehyde/B. Falck, N. A. Hillarp, G. Thieme, A. Torp//J. Histochem. &Cytochem. -1962. -Vol. 10. -P. 348-354.
