Нейровазальные отношения в энтеральной нервной системе человека

Автор: Марков И.И., Низаметдинова Д.Р.
Журнал: Морфологические ведомости @morpholetter
Рубрика: Оригинальные исследования
Статья в выпуске: 4 т.33, 2025 года.
Бесплатный доступ
Цель: доказать существование в энтеральной нервной системе человека различных морфологических вариантов нейровазальных отношений. Материалы и методы. Изучены аутопсийные фрагменты тонкой и толстой кишки новорожденных (n=9) и людей первого зрелого возраста (n=12) классическими гистологическими методами (окраска гематоксилином и эозином, парарозанилином с толуидиновым синим, азур II-эозином) и модифицированным универсальным методом импрегнации аргирофильных структур. Изучали топографию, морфологию и кровоснабжение ганглиев межмышечного и подслизистого нервных сплетений. Результаты. Установлен модульный принцип кровоснабжения ганглиев и межузловых тяжей энтеральной нервной системы человека. У новорожденных 79% ганглиев находятся в просвете лимфатических микрососудов, трофика нейроцитов осуществляется за счёт лимфы (нейролимфатический вариант). У 21% ганглиев выявлен нейролимфогематический вариант с наличием кровеносных микрососудов. У людей первого зрелого возраста функционирует экстраневральное микрососудистое русло с модулем сетевого типа (нейрогематический вариант). Микрососудистые модули межузловых нервных трактов построены по магистральному типу с интра- и экстраневральными микрососудами. Обнаружены артериовенулярные анастомозы и противоточные газовые обменники. Заключение. На всех этапах дифференцировки нейроцитов энтеральной нервной системы человека (стадии нейробласта, роста и созревания) функционируют соответствующие морфологические варианты нейровазальных отношений: интерстициальный, нейролимфатический, нейролимфогематический и нейрогематический, что обеспечивает оптимальные условия для дифференцировки и длительного устойчивого выживания гетерогенной популяции нейроцитов.
Энтеральная нервная система, нейровазальные отношения, нервные ганглии, кровоснабжение, лимфатические сосуды, кишечник, новорожденные, человек
Короткий адрес: https://sciup.org/143185414
IDR: 143185414 | УДК: 611.018.7:611.34 053.2/.6 | DOI: 10.20340/mv-mn.2025.33(4).993
Neurovascular relationships in the human enteric nervous system
Objective: To demonstrate the existence of various morphological variants of neurovascular relationships in the human enteric nervous system. Materials and Methods. Autopsy fragments of the small and large intestines from newborns (n=9) and adults of first mature age (n=12) were studied using classical histological methods (hematoxylin and eosin staining, pararosaniline with toluidine blue, Azure II-eosin) and a modified universal method for impregnation of argyrophilic structures. The topography, morphology, and blood supply of ganglia of the myenteric and submucosal nerve plexuses were examined. Results. A modular principle of blood supply to ganglia and interganglionic cords of the human enteric nervous system was established. In newborns, 79% of ganglia are located within the lumen of lymphatic microvessels, and neurocyte nutrition is provided by lymph (neurolymphatic variant). In 21% of ganglia, a neurolymphohematic variant with blood microvessels was identified. In adults of first mature age, an extraneural microvascular bed with a network-type module functions (neurohematic variant). Microvascular modules of interganglionic nerve tracts are organized according to a magistral type with intra- and extraneural microvessels. Arteriovenular anastomoses and countercurrent gas exchangers were detected. Conclusion. At all stages of differentiation of enteric nervous system neurocytes in humans (neuroblast stage, growth stage, and maturation stage), corresponding morphological variants of neurovascular relationships function: interstitial, neurolymphatic, neurolymphohematic, and neurohematic, which ensures optimal conditions for differentiation and long-term stable survival of a heterogeneous population of neurocytes.
Текст научной статьи Нейровазальные отношения в энтеральной нервной системе человека
(4).993
Источники кровоснабжения энтеральной нервной системы изучены недостаточно полно, они дают информацию лишь об ее ангиоархитектонике и нейрокопиллярных отношениях [1–3]. При этом, отдельные кровеносные капилляры определяются в периневральных пространствах вегетативных нервных сплетений [2]. Они рассматриваются как интраганглионарные капилляры, подобные капиллярам головного мозга, способные формировать гистогематический барьер, идентичный гематоэнцефалическому барьеру [3]. И в то же время, данные о периневральных пространствах нервных сплетений кишечника представлены и в работах по морфологии лимфатической системы [4–6]. Они, далеко не всегда, но все же заполняются цветной массой Герота при интерстициальном ее введе- нии в стенку кишечника. Подобное представление о существовании гистогематического барьера в энтеральных ганглиях развивают и другие авторы [7–10]. Так, авторы [7] проанализировали полученные результаты с позиции теории о ги-стогематических барьерах и пришли к заключению о существовании в вегетативных ганглиях гистогематического и межтканевого барьера. Их назначение – создание изолированной постоянной внутренней среды для нервных структур, а функцию барьеров выполняют эндоневральные микрососуды и периневральный эндотелий. Однако, считается, что структуры этих барьеров не только недостаточно изучены [9], но и признаются не всеми исследователями [10]. Наиболее категоричен в этом [11], считающий, что в ганглиях энтеральной нервной системы нет ни соединительной ткани, ни кровеносных сосудов.
Таким образом, в литературе, посвященный микрососудистому руслу энтеральной нервной системы накопилось много спорных и нерешенных вопросов, требующих проведения детальных исследований для их решения. Более того, особые затруднения возникают при сопоставлении результатов, полученных морфологическими методами с результатами физиологических исследований. Часто они находятся в полном противоречии друг с другом. Причина тому – многочисленные артефакты как из-за несовершенства методов исследования, так и из-за чрезвычайной сложности структурной организации микрососудистого русла стенки кишечника.
Эта ситуация усложняется еще и наличием в ней энтеральной нервной системы и дополнительных структур, обеспечивающих ее гомеостаз.
ЦЕЛЬ ИССЛЕДОВАНИЯ: доказать существование в энтеральной нервной системе человека различных морфологических вариантов нейро-вазальных отношений.
МАТЕРИАЛЫ И МЕТОДЫ
Объект исследования – аутопсийные фрагменты тонкой и толстой кишки новорожденных (n = 9) и людей I зрелого возраста (n = 12). Материал получен из патологоанатомических отделений лечебных учреждений г. Самары в соответствии с Законодательством РФ.
Фрагменты кишечника после предварительной подготовки подвергались гидропрепарированию под стереомикроскопом и разделялись на слизистую и мышечную оболочки и подслизистую основу. Каждая из выделенных оболочек фиксировалось в 10% нейтральном аметаноль-ном формалине в отдельных флаконах. Гистологическая обработка для классических методов окраски осуществлялась по стандартным прописям [12, 13]. Готовились парафиновые блоки, из них получали срезы толщиной 5–7 мкм, которые после окраски, просветления, проведения через спирты заключались в канадский бальзам.
Для одновременного выявления нервных и сосудистых элементов энтеральный нервной системы человека был модифицирован универсальный метод импрегнации аргирофильных структур [14]. Фиксация расслоенных фрагментов стенки кишки (слизистой и мышечной оболочки и подслизистой оболочки) проводится без смены раствора в течение 1,5 месяцев. Увеличение сродства серебра к сосудистой стенке кровеносных и лимфатических микрососудов, к нейроцитам и их отросткам, и к сателлитной глии осуществляется предварительным осаждением на них гидрооксида магния, кальция и бария. В первоначальной прописи метода – это достигается перфузией через брюшную аорту – воротную вену их слабых растворов. В модифицированной прописи небольшие фрагменты оболочек стенки кишечника в 0,2% погружаются в растворы гидроокисей бария, кальция, магния на строго определенное время.
Концентрация первого раствора серебра и его экспозиция на препарате определяется опытным путем в каждом конкретном случае. На ранних сроках импрегнации более высокое средство к серебру проявляется у стенки кровеносных и лимфатических микрососудов, а на поздних сроках – у структурных элементов нервной ткани. После стандартных манипуляций (проведения через батарею спиртов и просветления в ксилоле) препараты толщиной до 40,0–80,0 мкм и площадью до 15 см2 (3,0×5,0 см) заключаются в канадский бальзам по методу Я. Кларка.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Дефинитивная энтеральная система включает в себя пять вегетативных нервных сплетений: 1) субсерозное (Шабадаша); 2) межмышечное внутреннее (Ауэбаха); 3) мышечное глубокое (Кахаля); 4) внутреннее кишечное (Генле); 5) подслизистое (Мейсснера) [3, 15]. Все сплетения состоят из ганглиев, связанных между собой межузловыми нервными трактами. Межмышечное нервное сплетение находится между мышечными слоями мышечной оболочки и опре- деляется во всех изученных отделах кишечника. В его составе выделены, кроме главного сплетения (Ауэбаха), еще и глубокое мышечное сплетение (Кахаля) и внутреннее кишечное (Генле). Эти сплетения образованы меньшими по размерам ганглиями и тонкими межганглионарными трактами. Подслизистое сплетение расположено на подслизистой основе и в его глубине как в тонкой, так и в толстой кишке. Его ганглии значительно меньше по размерам ганглиев основного межмышечного сплетения.
Подслизистое сосудистое сплетение также лежит в подслизистой основе и тонкой, и в толстой кишке, но значительно глубже, чем сплетение нервное. Кроме того, все ганглии подслизистого нервного сплетения непосредственно связаны с внутриорганными стволами блуждающих нервов.
Поверхностное же подслизистое нервное сплетение располагается в самом наружном слое подслизистой основы, в непосредственной близости к циркулярному слою мышечной оболочки. Оно образовано небольшими по размеру ганглиями, связанными между собой тонкими межузловыми трактами. Размеры ганглиев основного межмышечного и основного подслизистого нервных сплетений значительно отличаются: средняя площадь ганглия межмышечного сплетения в 1,75 раза превышает среднюю площадь ганглия подслизистого сплетения. Ганглии этих сплетений отличаются и по форме: ганглии межмышечного сплетения имеют удлиненную и округлую форму, а в подслизистом сплетении доминируют ганглии треугольной формы. Плотность ганглиев межмышечного и подслизистого сплетений в тонкой и в толстой кишке различны: количество ганглиев межмышечного сплетения в тонкой кишке в 2,0 раза (p<0,001) больше, чем в толстой кишке.
Энтеральная нервная система (ЭНС) развивается из клеток вагального и сакрального участков нервного гребня [17], нейробласты которых затем мигрируют вдоль блуждающих нервов и колонизируют кишечную стенку. При этом, дифференцирующиеся нейробласты формируют ганглии межмышечного сплетения, а остальные нейробласты мигрируют дальше [18]. Терминальные отделы толстой кишки колонизируются нейробластами из сакрального участка нервного гребня. Весь процесс колонизации нейробластами кишечника эмбриона человека продолжается 10 недель. Миграция нейробластов начинается на 4-й неделе развития, а уже на 8-9 неделе формируется межмышечное сплетение тонкой кишки, а на 12-14 неделе - организация ЭНС достигает дефинитивного строения [19]. Подслизистое нервное сплетение является производным межмышечного сплетения и формируется после миграции нейробластов из него в подслизистую основу кишечника.
У новорожденных значительная часть ганглиев (до 79%) находится в просвете лимфатических микрососудов, и трофика нейроцитов осуществляется за счет лимфы с более низким содержанием белка, чем в плазме крови. Это соответствует филогенетически более раннему диффузному способу трофики нервных структур, характерному для рептилий и земноводных.
Для этих ганглиев характерны: диффузный тип питания, отсутствие кровеносных сосудов, нейропиля и соединительнотканной капсулы. Большая часть нейробластов имеет крупное, эксцентрично расположенное ядро, у них нет отростков и глиоцитов. На этой стадии дифференцировки они находятся в просвете лимфатических микрососудов (рис. 1). Аналогичным образом осуществляется трофика нейроцитов желудочного ганглия речного рака Astacus astacus, находящегося в просвете крупного кровеносного сосуда [20]. Посредством соединительнотканных тяжей ганглий фиксируется к стенке сосуда и омывается циркулирующей гемолимфой. У черепах же интрамуральные нервные ганглии и хромаффинные элементы находятся уже в просвете крупных лимфатических коллекторов, и их трофика осуществляется только лимфой [21]. И только меньшая часть ганглиев (21%) новорожденных, формирующих основные сплетения ЭНС: межмышечное и подслизистое, уже имеют дополнительный источник трофики – кровеносные микрососуды (рис. 2).
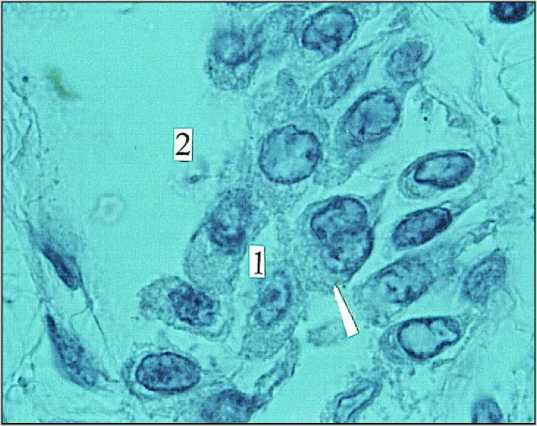
Рис. 1 . Ганглий (1) межмышечного нервного сплетения тонкой кишки новорожденного; 2) лимфатический микрососуд; ↑) дикарион. Окраска парарозанилином и толуидиновым синим. Ув. 900
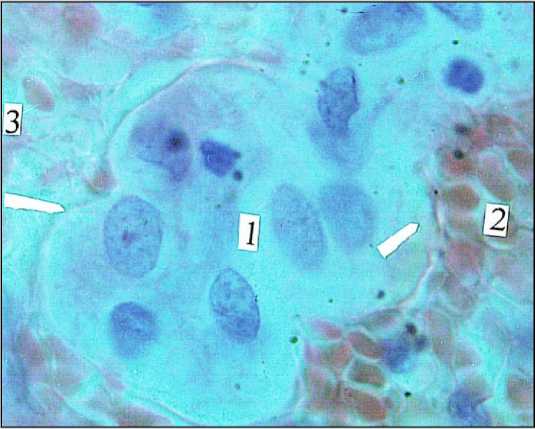
Рис. 2. Ганглий (1) межмышечного сплетения тонкой кишки (3) новорожденного; 2) венула; ↑) стенка лимфатического микрососуда. Окраска азур II-эозином. Ув. 900
Небольшое количество зрелых нейроцитов при огромном количестве нейробластов в ЭНС новорожденных, вероятно, является морфологической основой развития у них младенческих колик. В 90–95% случаев они имеют функциональный характер и даже рассматриваются как «условно» физиологическое состояние, как период адаптации желудочно-кишечного тракта ребенка грудного возраста [22, 23]. Причина в том, что «созревание нервной системы кишечника продолжается до 12–18-месячного возраста ребенка» [22, с. 162]. Нарушения же ее формирования ведут к развитию у новорожденных клинической картины некротического энтероколита, морфологической основой которого является аганглиоз или нейрональная интестинальная дисплазия [24–27]. Выделено два типа интестинальной дисплазии: типа А – аплазия и гипоплазия межмышечного нервного сплетения [24] и тип В – гиперплазия межмышечного нервного сплетения и нарушение развития подслизистого нервного сплетения [24, 25, 27]. Клинические проявления этой патологии варьируют от легких запоров до некротического энтероколита.
Морфологическая основа нейрональной интестинальной дисплазии типа А, – аплазии и гипоплазии межмышечного сплетения, не вызывает сомнений. Но, данные о морфологической основе дисплазии типа В, очевидно, нуждаются в тщательной проверке. Во-первых, нельзя признать «гигантскими» ганглии подслизистого нервного сплетения, содержащие более восьми нейроцитов. При этом, авторы [24, 26] неоправданно считают нормальными ганглии с двумя – четырьмя нейроцитами. Во-вторых, в этих «гигантских» ганглиях дифференцированные нейроциты могут полностью отсутствовать. Тогда идентифицированные авторами [24, 26] нервные клетки – это нейробласты, еще не участвующие в иннервации кишечника. Исходя из этого, клиническая симптоматика при инте-стициальной дисплазии действительно может нормализоваться [27, 28]. Однако, только тогда, когда нейробласты «гигантских» ганглиев пройдут все стадии дифференцировки, получат статус «зрелых» нейроцитов и станут выполнять свою функцию. В-третьих, в нарушение закономерностей естественной миграции пронейробластов: в начале миграция в межмышечное, а только затем – в подслизистое нервное сплетение, – «гигантские» ганглии почему-то формируются в подслизистом, а не в межмышечном сплетении [25].
ЭНС людей I зрелого возраста формируют ганглии двух типов: плоскостные, с диффузным расположением нейробластов и нейроцитов, и локальные, с их компактной упаковкой (рис. 3, 4).
Основа структурной организации микрососу-дистого русла плоскостных ганглиев – модульный принцип с тремя различными вариантами ее специализации: ганглионарным, межганглионарным и стволовым. В ганглиях функционирует экстраневральное микрососудистое русло с модулем сетевого типа с короткими, но широкими артериолами и венулами (рис. 5). Морфометрические параметры микрососудов модуля: артериол (диаметр – 14,0–17,0 мкм), прекапиллярных артериол (диаметр – 7,0–10,0 мкм), капилляров (диаметр – 5,5–6,1 мкм), посткапиллярных венул (15,0–18,0 мкм) и венул (диаметр 20,0–24,0 мкм) свидетельствуют о его низком сосудистом сопротивлении. Значительно более низком, чем сопротивление сосудистых конструкций других морфологических структур кишечника [29]. Эта особенность модуля обеспечивает эффективный кровоток в ганглии в различные периоды его функциональной активности. При этом, нейроциты в ганглиях не имеют непосредственного контакта с обменными микрососудами, поскольку удалены от них на расстояние не менее 15,0–20,0 мкм.
Локальная регуляция кровотока в микрососудах модуля обеспечивается гладкомышечными и эндотелиальными сфинктерами и внекапил-лярными путями кровотока, относящимися к пре- и посткапиллярных полушунтов. Локализация их в микрососудистом русле плоскостных ганглиев позволяет считать, что их гемодинамическая роль не столь велика, как это принято считать. Однако, обладая минимальным сосудистым сопротивлением, полушунты функционируют независимо от колебаний перфузионного давления, и тем самым способствуют стабилизации трансорганного кровотока [30].
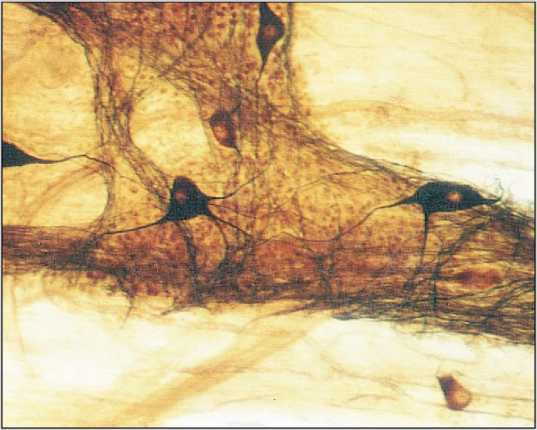
Рис. 3. Плоскостной ганглий с диффузным расположением нейроцитов. Подслизистое нервное сплетение толстой кишки мужчины I зрелого возраста. Универсальный метод импрегнации. Ув. 400
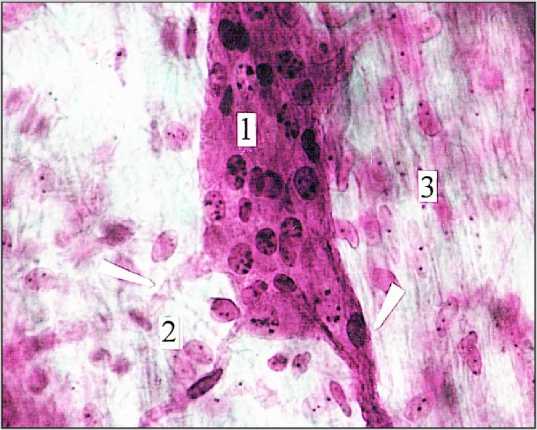
Рис. 4. Локальные нервный ганглий (1) с компактным расположением нейробластов в мышечной оболочке тонкой кишки. Мужчина I зрелого возраста. 2↑) отростки нейробластов. Универсальный метод импрегнации. Ув. 400
Микрососудистые модули межузловых нервных трактов построены по магистральному типу. В этом случае артериола, венула, посткапиллярные венулы и капилляры следуют параллельно, имея значительную протяженность, но очень мало поперечных связей. В таких модулях взаимодействуют экстра- и интраневральные микрососуды. Интраневральные капилляры есть в межузловых трактах диаметром от 30,0 до 150,0 мкм. Просвет их заполнен эритроцитами, а это свидетельствует о высоком капиллярном гематокрите (рис. 6). Стволовые микрососудистые модули образованы артериолами диаметром 25,0–35,0 мкм, вступающими под углом в периневральные пространства интраорганных нервов. Кроме подобных сосудисто-нервных взаимоотношений, существуют, очевидно, и другие, в которых взаимодействуют нервные стволы и лимфатические микрососуды. Так, и в межмышечном, и в подслизистом нервных сплетениях нервные стволы постоянно идентифицируются в лимфатических микрососудах (рис. 7). О том, что периневральные пространства межмышечного нервного сплетения связаны с лимфатическим руслом стенки кишечника значительно ранее указывали [4, 5]. Лимфатические сосуды, как продолжение периневральных пространств, заполнялись массой Герота, позже и значительно труднее, чем периневральные пространства [5]. Очевидно, что связи периневральных пространств с лимфатическим руслом существует не только в кишечнике. Это, вероятно, общая закономерность, поскольку они обнаружены также и в надкостнице [31], и в симпатических ганглиях [32].
Особое значение в гемоциркуляции в плоскостных ганглиях имеют артерио-венулярные анастомозы (АВА). Скорость кровотока в них и в капиллярах резко отличается. Так, через капилляр диаметром 10,0 мкм 1,0 мм3 крови проходит в течение 6 часов. Такой же объем крови проходит через АВА диаметром 100,0 мкм всего лишь за 2,0 сек. Эти расчеты сделаны на основе формулы Пуазейля и важны, в первую очередь, для сравнения [33]. В последнее время, кроме уста- новленных еще в 1961 году восьми функциональных назначений АВА [34], открыты их четыре новых роли: 1) в оксигенации портальной крови [35]; 2) в запуске восходящей дилатации магистральных артерий [36]; 3) в циркуляции нейтрофильных гранулоцитов [37]; 4) в увеличении сосудистой проницаемости [38].
И все-таки, несмотря на органную специфичность ЭНС, ее внекапиллярные пути кровотока и, прежде всего, АВА – типичные классические варианты АВА (рис. 8). Это – морфологически и функционально специализированные конструкции брадитрофных тканей, выполняющие гидродинамические, реологические и другие, изложенные выше, функции. Именно они и обеспечивают, несмотря на функциональные и экстремальные колебания общего перфузионного давления, стабильный кровоток в нервных ганглиях, межузловых трактах и нервных стволах. Кроме того, в сосудистых конструкциях межузловых трактов и нервных стволов обнаружены также и противоточные газовые обменники. Их функцию выполняют параллельно идущие в составе этих конструкций артериолы и венулы, находящиеся друг от друга на расстоянии не более 12,0–18,0 мкм. Такие теснейшие взаимоотношения между двумя противоположными потоками крови позволяют значительно повышать содержание кислорода в венозной крови. Чем интенсивнее кровоток, тем медленнее происходит обмен газов, тем менее эффективна работа газового шунта. Но, при замедлении артериального кровотока в условиях редуцированного кровотока, кровь насыщается углекислотой, поступающей из венозной крови. В связи с этим, противоточный обменник начинает функционировать как противоточный умножитель – развивается гиперкапния, следствием которой является максимальная дилатация микро-сосудистого русла межузловых трактов и нервных стволов. Более того, микрососуды перестают реагировать на различные вазоактивные вещества [39]. По всей вероятности, этим объясняется достаточно высокая устойчивость ЭНС к острой и хронической ишемии [40, 41].
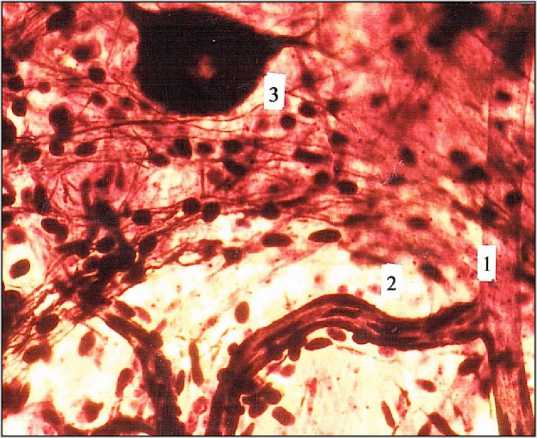
Рис. 5. Кровеносные микрососуды (1, 2) плоскостного ганглия (3) межмышечного нервного сплетения тонкой кишки. Женщина I зрелого возраста. Универсальный метод импрегнации. Ув. 900
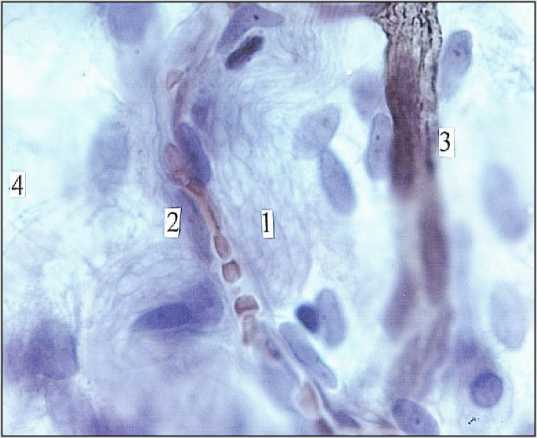
Рис. 6. Микрососудистое русло межузлового тракта (1) подслизистого нервного сплетения (4) тонкой кишки мужчины I зрелого возраста. 2) капилляр; 3) посткапиллярная венула. Импрегнация по Ранвье. Докраска гематоксилином и эозином. Ув. 600
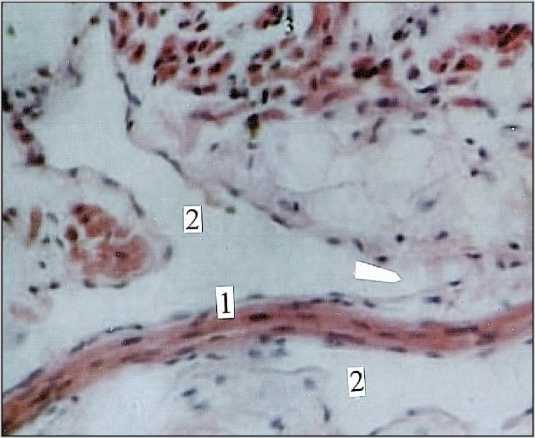
Рис. 7. Нервный ствол (1) в лимфатическом микрососуде (2) мышечной оболочки (3) стенки тонкой кишки. Мужчина I зрелого возраста. Окраска железных гематоксилином. Ув. 200
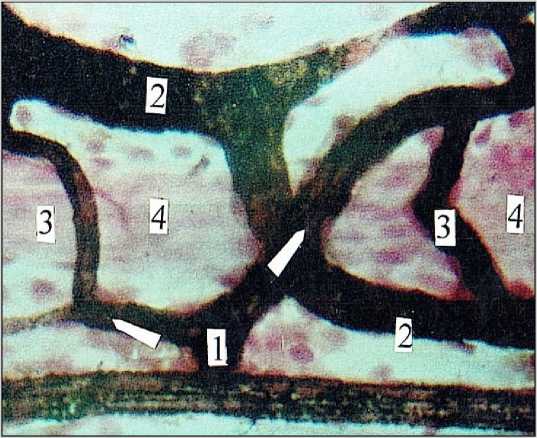
Рис. 8. Артериоло-венулярный анастомоз (3↑) в подслизистом нервном сплетении толстой кишки. Женщина I зрелого возраста. 1) артериола; 2) венула; 4) нервный ствол. Импрегнация по Ранвье. Докраска гематоксилином. Ув. 200
Локальные ганглии с компактным расположением нейробластов обнаружены в мышечной оболочке, в собственной пластинке слизистой оболочки и в подслизистой основе стенки кишечника людей I зрелого возраста. Их площадь в 2–3 раза меньше площади плоскостных ганглиев, и в них нет ни нейропиля, ни глиоцитов. Число нейробла- стов варьирует в них от 12 до 56. Это крупные клетки с большим, интенсивно импрегнирован-ным ядром овальной формы, содержащим многочисленные (от 5 до 10) крупные глыбки хроматина. У них светлая нейроплазма, лишенная нейрофибрилл, но граница между ней и ядром – четкая. Нейробласты упакованы настолько компактно, что между ними формируются плотные контакты и щелевые контакты с последующим образованием дикарионов. Другая особенность локальных ганглиев - это источники их питания (рис. 9). В отличии от плоскостных ганглиев с диффузным расположением нейроцитов и собственным мик-рососудистым руслом, локальные ганглии или отдельные нейробласты и нейроциты находятся в просвете лимфатических микрососудов или в ин-терстиции. То есть, их источники питания - лимфа или интерстициальная жидкость.
Между ганглиями и окружающими их мышечными и соединительно-тканными структурами всегда определяется тонкая соединительнотканная прослойка. Однако, полностью сформированные капсулы у локальных ганглиев, все-таки, отсутствуют. При этом каждый из них находится на определенной стадии дифференцировки. Так, нейробласты одних ганглиев не принимают участия в иннервации кишечника: для выполнения этой функции у них нет морфологических «инструментов» - ни фибриллярного аппарата, ни нейритов, ни дендритов. Известно, что морфологические структуры, подобные локальным ганглиям, есть и в головном мозге. В них нейроциты также компактно сгруппиро- ваны и плотно прилежат друг к другу. Это зубчатая извилина гиппокампа, поля СА1, СА2 и гранулярный слой коры мозжечка [42, 43, 44]. И в первом, и во втором случае, т.е. в локальных ганглиях и в указанных структурах головного мозга отсутствует выраженная глиальная защита. А это значит, что в них нет препятствий для формирования межнейронных синцитиальных связей, а, в последующем, и для образования дикарионов [43, 44]. При этом, если в первом случае трофика нейробластов и нейроцитов осуществляется лимфой, то во втором случае – ликвором [45, 46].
Отдельные локальные ганглии обладают нейритами, сформированными униполярными нейробластами. И они вступают в окружающие ганглии ткани. Их число незначительное: от 1 до 9. При этом, они значительно отличаются от нейритов плоскостных нервных сплетений: они слабо импрегнированы, имеют широкий равномерный диаметр и содержат одно безмиелиновое волокно (рис. 10). Более того, они объединяют близко расположенные локальные ганглии, выполняя для них роль синцитиальных комиссур.
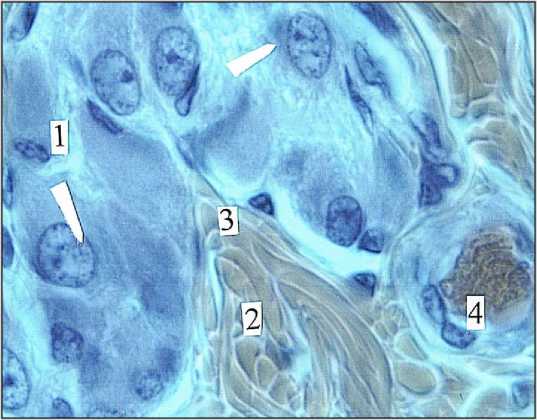
Рис. 9. Локальный ганглий с компактной упаковкой нейро-цитов (1|) в мышечной оболочке (2) толстой кишки. Мужчина I зрелого возраста. 3) оболочка ганглия; 4) венула.
Окраска гематоксилином и эозином. Ув. 400
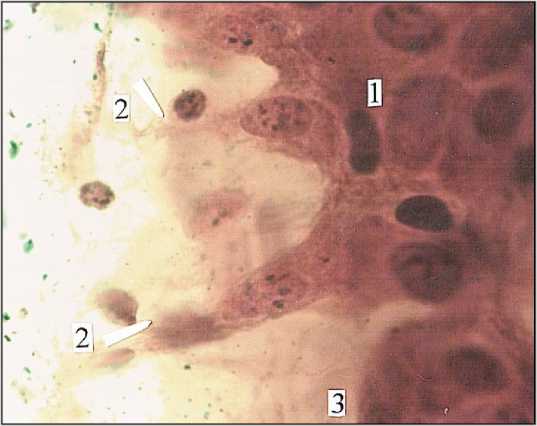
Рис. 10. Локальный ганглий (3) с компактной упаковкой нейроцитов (1) в собственной пластинке слизистой оболочки толстой кишки. Женщина I зрелого возраста. Универсальный метод импрегнации. Ув. 900
Соотношение числа нейробластов к числу отходящих от локальных ганглиев нейритов находится в пределах 15:1. Так, 9 нейритов идентифицировано у ганглия, содержащего 53 нейробласта. У ганглиев даже с одинаковым количеством нейробластов всегда определяется разное количество отростков, но эти вариации чрезвычайно малы – не более 2–3 отростков. Это означает, что даже у людей I зрелого возраста лишь незначительная часть нейробластов имеет нейриты. Они находятся на определенной стадии специфической дифференцировки и только приобретают статус дефинитивных униполярных нейроцитов. Для них характерна типичная локализация – на периферии ганглиев. В глубине же ганглиев у нейробластов нейриты отсутствуют, и они продолжают сохранять между собой плотные контакты, образуя единый синцитиальный кластер. Поскольку нейропептиды синтезируются и в головном мозге, и в ЭНС, то не исключено, что их продуцентами в ЭНС являются нейробласты и нейроциты локальных ганглиев. Так, установлено наличие субстанции и энкефалинов в нейроцитах ганглионарных сплетений ЭНС [47], доказано, что большие запасы тиролиберина и нейротензина сосредоточены в желудочно-кишечном тракте [48, 49]. Но необходимо отметить, что синтезируемые головным мозгом нейропептиды секретируются не в кровь, а в ликвор [50–53]. Исходя из этого положения, очевидно, что и расположенные в просвете лимфатических микрососудов локальные ганглии ЭНС, также синтезирующие нейропептиды, тоже секретируют их не в кровь, а в лимфу.
ЗАКЛЮЧЕНИЕ
Быстрое накопление новых данных о морфологии и физиологии нейроцитов привело не только к пересмотру многих общепринятых постулатов, но и к признанию известных, но скептически воспринимаемых фактов [54]. Так, доказано, что представления о чрезвычайной уяз- вимости нейроцитов оказались значительно преувеличены [55, 56, 57]. Высокая резистентность нейроцитов к ишемии была подтверждена в экспериментах, и она значительно повышалась в условиях гипотермии. В итоге, результаты экспериментов дали основание авторам [58, 59, 60] констатировать о «невероятном продлении жизни нейронов» гиппокампа морских свинок при их ишемии или полной апоксии, но в условиях гипотермии.
Но информации о парадоксальной реакции нейроцитов на высокое парциальное напряжение кислорода была известна значительно раньше [61, 62, 63].
В норме и головной мозг, и ЭНС потребляют меньше кислорода, чем почки (7,0 см3 / 100 г / мин) и сердце (10,0 см³ / 100 г / мин). ЭНС – 3,6 см³ / 100 г / мин, головной мозг – 3,2 см³ / 100 г / мин [64, 65]. Более того, и головной мозг, и ЭНС образуют значительно меньше тепла, чем почки (34,5 кал / 100 г / мин) и сердце (48,0 кал / 100 г / мин). ЭНС – 17,4 кал / 100 г / мин, а головной мозг – 15,4 кал / 100 г / мин [64, 65]. И головной мозг, и ЭНС постоянно находятся в жидкой среде: головной мозг перфузируется ликвором с чрезвычайно низким содержанием белка (0,028%), ЭНС – интерстициальной жидкостью и лимфой с содержанием белка не более 2,61% [65].
Эти параметры являются оптимальными как для нейроцитов головного мозга, так и нейро-цитов ЭНС. Они создают благоприятные условия для дифференцировки и длительного устойчивого выживания чрезвычайной гетерогенной и избыточной популяции нейроцитов. На всех этапах дифференцировки: стадии нейробласта, стадии роста нейроцита и стадии созревания нейроцита [66, 67], они обеспечены соответствующим морфологическим вариантом нейро-вазальных отношений: интерстициальным, нейролимфатическим, нейролимфогематиче-ским и нейрогематическим.
