Некоторые аспекты гнездовой биологии и экологии Motacilla tschutschensis plexa и Motacilla flava beema в Западной Сибири

Автор: Емцев А.А., Попов С.В., Ван Остин Х.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 3, 2016 года.
Бесплатный доступ
Таксономический комплекс жёлтых трясогузок Motacilla flava sensu lato имеет сложную внутреннюю структуру, до сих пор не разрешенную в полной мере по некоторым аспектам. В настоящей публикации рассматриваются отдельные вопросы гнездовой биологии и экологии двух подвидов - Motacilla tschutschensis plexa и Motacilla flava beema. Птицы изучались в юго-западной и центральной частях Западной Сибири. Производился учет особей на пробных площадках и маршрутах, осуществлялось описание гнезд и яиц, отмечалась окраска взрослых трясогузок. Выявлено, что рассматриваемые подвиды для устройства гнезд выбирали разные микростации и отличались по основным оологическим показателям. На техногенно трансформированных участках комплексных верховых болот плотность гнездования M. t. plexa увеличивалась по сравнению с относительно менее нарушенными местообитаниями. Некоторые взрослые особи этого подвида характеризовались серой окраской шапочки и маски, выраженной бровью.
Гнездовая биология, экология, западная сибирь
Короткий адрес: https://sciup.org/147204777
IDR: 147204777 | УДК: 598.829:591.521:591.9(5
Some aspects of nesting biology and ecology of Motacilla tschutschensis plexa and Motacilla flava beema in Western Siberia
The taxonomy of the Yellow Wagtail Motacilla flava sensu lato is very complicated and not fully resolved as yet. Here, we report on the nesting biology and ecology of two subspecies - Motacilla tschutschensis plexa and Motacilla flava beema. The birds were studied in southwest and central parts of Western Siberia. The accounting of animal unit on trial area and routes was made, the nests and eggs were described and the plumage characteristics of adult Wagtails were noted. It appeared that both subspecies chose different microhabitats where they constructed their nests and they differed with respect to the main oological indicators. The nesting density of M. t. plexa increased in technogenic transformed sites of complex raised bogs, compared to relatively less violated habitats. Some adult individuals of M. t. plexa were characterized by a gray coloring of the hat and the mask expressed by the eyebrow.
Текст научной статьи Некоторые аспекты гнездовой биологии и экологии Motacilla tschutschensis plexa и Motacilla flava beema в Западной Сибири
Жёлтая трясогузка Motacilla jlava sensu lato — один из наиболее «проблемных» в плане индивидуальной и географической изменчивости таксономических комплексов птиц, распространенных в Палеарктике [Маут, 19561 Редькин и др., 2015]. Среди специалистов долгое время не существовало единого мнения относительно точного числа таксономических единиц данной группы, области их гнездования и данной группы, области их гнездования и пространственных взаимоотношений различных форм. Вопросы систематического плана и детали распространения жёлтых трясогузок не теряют своей актуальности и в настоящее время.
В Западной Сибири преимущественно встречается две четко различимые по внешним морфологическим
(С Емцев А. А., Попов С. В., ван Остин X. Герман, 2016
признакам формы трясогузок, по современным представлениям относимые к двум подвидам разных видов (берингийской (жёлтой) и жёлтой), - MotadUa tschatscheHsk plexa (Tliayer el Bangs. 1914) и Mo-CadUa flava beema (Sykes, 1832) [Коблик и др.. 2006; Коблик, Архипов, 2014; Рябичев, 2014]. Наряду с другими более-менее выраженными признаками, первый во взрослом наряде имеет темно-серую шапочку и темную маску (у самок иногда проявляется бровь), населяет центральную и северную части Западной Сибири [Рябицев, 2014]. У второго в брачном наряде серая или голубовато-серая окраска головы и белая бровь (может отсутствовать), распространение главным образом в центральной и южной частях рассматриваемой территории. Возможны встречи Мо-tadila flava flava Linnaeus, 1758, но площадь гнездового ареала этого подвида в Западной Сибири очень незначительна и охватывает приграничные районы на востоке от Урала [Там же]. Также вероятны регистрации других форм трясогузок, в том числе гибридных, в южных районах. Следует отметить, что ранее XL t. pkxa относили к форме «thunbergi», выделяя ее в самостоятельный подвид желтой трясогузки [Степанян. 2003; Рябицев, 2008].
Предпочтения строго определенных типов местообитаний X/. /. pkxu (XLf. ihunbergd и М. / Ытаъ по-видимому. определяющие их симпатрию отмечались некоторыми исследователями [Бойко, 1998а, б; Редькин, 2001] и уже были рассмотрены нами ранее [х ал Oosten, Emtsev; 2013]. В данном сообщении, не пре тендуя на какие-либо выводы систематического характера, мы нашли интересным отразить отдельные аспекты гнездовой биологии и экологии трясогузок анализируемого комплекса в Западной Сибири их распространение и «нетипичную» окраску’ некоторых взрослых особей.
Методы исследований
Исследовались центральная, северная и южная части Западной Сибири, согласно административному делению принадлежащие Ханты-Мансийскому — Югре и Ямало-Ненецкому автономным округам, Свердловской и Челябинской областям (рис. 1). Сроки работ — с 2000 по 2015 гт. Работы велись в нескольких геоботанических подзонах — северной и средней тайге, редколесьях и лесостепи [Растительный покров..., 1985], Наши исследования, прежде всего, были ориентированы на изучение населения птиц на этой территории. Данные по трясогузкам М. I. ркха и М. / Ьета (белоухим жёлтым трясогузкам) собраны попутно.
В каждом районе мы определяли по нескольку ключевых участков, которые посещали в отдельные годы обозначенного периода. Общее число обследованных участков составило 35 (рис. 1, табл. 1). На участках устанавливались основные типы местообитаний, где закладывались учетные площадки или проводились маршрутные учеты.
Таблица 1
Распространение подвидов берингийской (жёлтой) и жёлтой трясогузок^ занимаемые местообитания и годы регистраций
|
Таксон, номер и геелрафическая привязка ключевого участка |
Координаты |
Год |
Местообитание |
|
MotadUa tschtuschensis pkxa |
|||
|
2. Район г. Радужный |
6242.6Н 78°9.4Е |
2004 |
ПБВГМОБ |
|
3. Район г. Радужный |
62е 16.5Н 784 1.5Б |
2004 |
НБВ1 МОБ, ВР? |
|
4, Район г. Радужный |
62°9.1Н 77°54.5Е |
2004 |
ПБВГМОБ |
|
6. Окрестности г. Лянтор |
6Р35.8Н 7241.ЗБ |
2005 |
Окраина ПБВГМОБ |
|
7, Район оз. Антон юр |
61°52.0Н 7Р21.5Б |
2005 |
ПБВГМОБ, ПД ПБВГМОБ |
|
8. Район оз. Вилинглор |
61М4.6Н 7149.9Б |
2005, 2006 |
ПБВГМОБ, ПД ПБВГМОБ |
|
9, Окрестности д. Русски гская |
624.6N, 73°28.1Е |
2006 |
ПБВГМОБ |
|
10. Район оз. Пильтлор |
62620.4Н 72°58.6Б |
2006 |
ПД ПБВГМОБ |
|
11. Район оз. Куглопъяунлор |
62°56,5Н724 5.6Б |
2006 |
ПБВГМОБ, ПД ПБВГМОБ |
|
12. Район оз. Нанканкиллор |
6245.3’N, 72°5.8'Н |
2006 |
НД ПБВГМОБ |
|
13. Окрестности вахтового пос. Ьарсуков-ский |
64°20.бН 75°36.9Б |
2007 |
ТППБВБМО, окраина ПБВГМБ |
|
14, Междуречье р. Нгарка-Варкьяха и Апо-куяха |
64°7.7Ч 7547.8Е |
2007 |
ТППБВБМО |
|
15. Район р. Хасуйяха (приток р. Пурпе) |
64625.6Н 75°22.4Б |
2007 |
ТППБВБМО, окраина ЗРЛКСМ, окраина ЗРСПКБ |
|
16, Окрестности нежилого пос. Воет по |
644.1N, 7841.5Е |
2008 |
ТППБВБМО |
|
18. Окрестности пос. Уренгой |
65°58.0Н 78°34.0Б |
2010 |
ТППБВБМО |
|
19. Район нижнего течения р. Вынга |
6149.9Н72°31.2Б |
2011 |
ПОЛИ |
|
20. Окрестности г. Надым |
6543.4^, 724 1. ГН |
2012 |
11 П1БВБМО |
|
22. Окрестности г. Ноябрьск |
634 0.5Н 7544.9Б |
2013 |
ТППБВБМО |
|
23, Окрестности д, Юган |
6О°52.8Н 7341,ЗБ |
2015 |
ПБВГМБ |
|
34. Район полевого стационара Мухрино |
6043.2Н 6842.1Б |
2009 |
ПБВГМБ |
Окончание табл. 1
|
Таксон, номер и географическая привязка ключевого участка |
Координаты |
Год |
Местообитание |
Motacilla flava beema
|
1. Окрестности д. Сай гати на |
61°15.7Н 72°53.4*Е |
2002^ 2008. 2009, 2011 |
СРТЛ, ПОЛИ |
|
5. Окрестносги д. Тренька |
61°13.0'N, 69°3.8'Е |
2004 |
ИЧЗЛ |
|
17. Окрестности г. Сургут |
61°15.7Н 73°20.ГЕ |
2009, 2012, 2013 |
ПЗБ. ПОЛИ |
|
19. Район нижнего течения р. Вынга |
61°19.9Н 72°31.2'Е |
2011 |
ПОЛИ |
|
21. Окрестности г. Сургут |
61°20.7'N, 73°25.8'Е |
2013 |
РТЗЛП |
|
23. Окрестности д. Юган |
60°52.8'N, 73°41.3'Е |
2014, 2015 |
СРТЛ, ПД РТЗЛ |
|
24. Окрестности с. Сипавскос |
56О14.4Н61°53.3'Е |
2000-2011 |
РТЗЛИК |
|
25. Район оз. Б. Сун гуль |
56°22.0Н61°37.8,Е |
2000-2012 |
ОБПРТЗЛЗ, ОКЛ |
|
26. Окрестности г. Каменск-Уральский |
56°19.8'N,61°57.6,E |
2000-2013 |
РТЗЛИК |
|
27. Окрестности пос. Синарский |
56°8.Ж61°59.ГЕ |
2000-2014 |
РТЗЛИК |
|
28. Район оз. Куракпи-Маян |
56О5.5Н6Г54.4*Е |
2000-2014 |
РТЗЛИК, ОКЛ |
|
29. Район оз. Маян |
55°58.4'N,61°49.5,E |
2000-2014 |
РТЗЛИК, ОКЛ |
|
30. Район оз. Маян |
55O57.1*N, 62°0.3'Е |
2000-2014 |
РТЗЛИК, ОКЛ |
|
31 Район оз. Уелги |
55°47.6Н61О29.9'Е |
2000-2014 |
РТЗЛИК, ОКЛ |
|
32. Окрестносги д. Шанша |
61°05.3*N, 69°27.6'Е |
2009 |
ПЗЛ |
|
33 Район полевого стационара Мухрино |
60°56.3'N, 68°42.3'Е |
2009 |
ПЗЛ |
|
35. Западнее г. Ханты-Мансийск, р. Иртыш |
60°59.ГМ, 68°57.2'Е |
2009 |
ПЗЛ |
Примечание. Условные обозначения: ВР — высохший рям; ЗРЛКСМ — заболоченный разреженный лишайниково-кустарничковый сосняк с мелколесьем; ЗРСГТКБ — заболоченный редкостойный пойменный кедрач с наличием березы; ИЧЗЛ — иван-чайный злаковый луг; ОБПРТЗЛЗ — осиново-березовые перелески среди разнотравнозлаковых лугов и залежей; ОКЛ — осоково-кочкарпый луг; ПБВГМБ — плоскобугристое верховое грядовомочажинное болото; ПБВГМОБ — плоскобугристое верховое грядово-мочажинно-озерковое болото; ПД — придорожный участок; ПЗБ — пойменное закустареннос (с ивами) болото; ПЗЛ — пойменный злаковый луг; ПОЛИ — пойменный осоковый луг с зарослями ив; РТЗЛ — разнотравно-злаковый луг; РТЗЛИК — разнотравно-злаковый луг с ивняками и редкими колками; РТЗЛП — разно травно-злаковый луг с покосом; СРТЛ — суходольный разнотравный луг; ТППБВБМО — тундроподобное плоскобутристое верховое болото с мочажинами и озерками.
? — подвидовая (видовая) принадлежность птиц в первоисточнике нс уточняется, форма гипотетически выделена на основании других наблюдений.
Использованы данные А.В. Сесина (3-й ключевой участок, высохший рям) и А.В. Поргунёва (1-й ключевой участок, 2008 г., суходольный разнотравный луг).
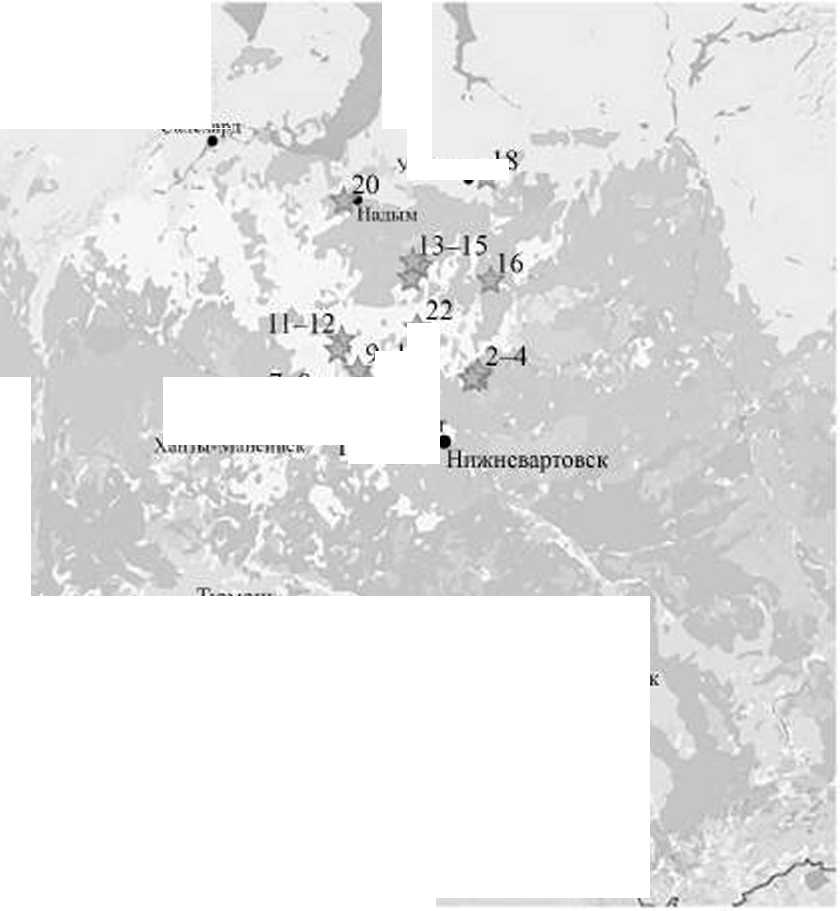
Салсхпр,!
32-35.5
Д урит
XatUN-Mad^k* П7-О1 •
Нижневартовск
Уренгой 8
9-10
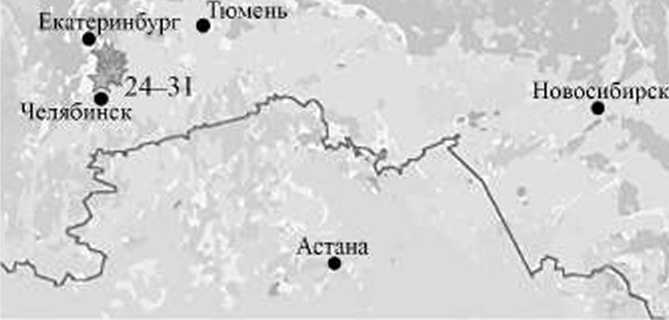
Рис. 1. Места проведения исследований в Западной Сибири:
1-35 — ключевые участки
На комплексных верховых болотах, преобладающих в северной части обозначенного выдела, на каждом участке осуществлялась закладка 2-4
контрольных площадок общей площадью 2-5 км2. Ввиду преобладания здесь открытых и полуоткры-тых пространств, по мере возможности, площадки брались с как можно большим размером учетной площади — 1-3 км2. Когда распределение вида (подвида) в местообитании было более-менее равномерным. учетную площадь сокращали до 1-0.5 км2. Если обследованная площадь местообитания была недостаточной для оценки плотности гнездования или обилия птиц, такой пересчет для них нс производился. Большие площади обрабатывались нами в течение нескольких дней, при многократном посещении. Территориальные самцы или пары. беспокоящиеся или кормящие птицы принимались за гнездящихся. Пролетные, негнездящиеся птицы или птицы с невыясненным характером пребывания отмечались отдельно. Особенность трясогузок окрикивать наблюдателя группой из обитающих по соседству особей, позволила оценивать их число па определенном участке более точно.
Дия выявления распределения птиц в местообитаниях. подверженных более сильному антропогенному воздействию, учеты проводились в непосредственной близости от дорог, промышленных сооружений и вахтовых поселков. У дорог посреди комплексов вер- ховых болот закладывались площадки длиной 3-7 км и шириной 140 м, Ширина площадки являлась оптимальной с учетом дальности обнаружения птиц и степени трансформации местообитаний. На нефтепромыслах в 10-30 м или более от обочин дорог, как правило, размещались трубопроводы и ЛЭП, В удалении 100-150 м от дорог их наличие отмечалось редко. Следовательно, в учетную полосу попадали существенно трансформированные местообитания.
Также нами исследовались наиболее характерные лесные, лесостепные и пойменные местообитания. В пойменных лесах и сосняках закладывались площадки до 0.35-0.55 кмЗ Когда их плошддь составляла менее 0.2 км', показатель плотности гнездования не рассчитывался.
Кроме учетов, производились поиски и обследования гнезд изучалась гнездовая биология обсуждаемых подвидов. Степень насиженности яиц оценивали по признаку плавучести (флотации) их в воде [Rahn, Аг, 1974; Медине, Блум, 1976; Dunn et al, 1979; Меднис, 2002 и др ]. За время работ найдено 25 гнезд (16 Н. t. plexa и 9 М. / bemaY
Объем яиц находили по формуле А.Л. Романова и А.И. Романовой (1959), модифицированной D.F. Hoyt [1979]; V = 0,51 *L*B2. где V — объем (мм\ L — длина (мм ). В — ширина (мм) яйца. Индекс удлиненности (%) вычисляли по формуле Sph = 100>BZL [Мянд. 1988]. Статистическую обработку данных выполняли с помощью программы StatPliis 2007. Окраску отдельных особей М. t. plexa описывали по фотографиям.
Результаты и их обсуждение
Типологизация размещения гнезд в местообитаниях
Все особи М. I. plexa и М. / Ьета с гнездовым поведением наблюдались в открытых или полуоткрытых местообитаниях (табл. 1). При этом М. г. plexa отмечены практически исключительно на комплексных верховых болотах (19 ключевых участков; 2-4, 6-16, 18, 20, 22. 23. 34). Главным образом они представляли собой олиготрофные болотные системы с преобладанием грядово-мочажинно-озерковых комплексов. багульниково-ерниково-сфагновые на буграх и осоково-сфагновые в мочажинах, характерные для средней тайги и южной части северной тайги Западной Сибири, а также тутцфоподобные открытые плос-кобутристые верховые болота. багульниково-ерникюво-лишайниковые на буграх и осоково-сфагновые в мочажинах, распространенные в северной части северной тайги и редколесьях Западной Сибири. По границам комплексных верховых болот, где также были встречены М Г. plexa. распространены низкорослые сосновые рямы с мочажинами, озерками, примыкающие к различным соснякам или пойменным лесам.
М. / Ьета регистрировались на пойменных лутах с ивами, суходольных разнотравно-злаковых лутах средней тайги (9 ключевых участков; 1, 5. 17. 19, 2 L 23. 32 3 3, 3 5). а также на разнотравно-злаковых лугах с ивняками и отдельными колками, осоково-кочкарных лутах, пойменных разнотравно-злаковых лутах по долинам средних рек лесостепи (8 ключевых участков: 24-31). Лишь на пойменном осоковом лугу с отдельными зарослями ив ключевого участка 19 наблюдали птиц (по-вцдимому, гнездящихся) с внешними признаками как М / Ьета, так и М. t. plexa. Использование одного местообитания двумя формами наблюдается достаточно редко и. по всей вероятности, также происходит в условиях симпатрии [Жуков, 2011].
Размещение гнезд М. L plexa нами подразделено на следующие типы:
-
- под прикрытием пушицы у основания ос кочки, примыкающей к сфагновому бугру среди подобных травяно-моховых образований с кустарничками и пушицей в отдалении от открытой воды, площадь межбутровых понижений около гнезда незначительна (и = 2);
-
- под прикрытием сфагнума (нависает над гнездом) на склоне бугра с редкими хамедафне и морошкой, соседствуют другие бугры с кустарничково-сфагновым покровом и сильно обводненная мочажина с пушицей (п = 1);
-
- под прикрытием кладонии (нависает над гнездом) на склоне бугра с лишайншюво-кустарничкоБым покровом среди подобных бугров в 12 м от озера (п = 1):
-
- под прикрытием кладонии (нависает над гнездом) и багульника на бугре среди других бугров с кустарничково-лишайниковым покровом в отдалении от открытой воды, площадь межбутровых понижений около гнезда незначительна (п = 3):
-
- под прикрытием кладонии (нависает над гнездом), кустарничка или кустарничков (берёзы карликовой, багульника, брусники или подбела) на бугре или его склоне среди других бугров с кустарничковолишайниковым или кустарничково-сфагновым покровом около озера или сильно обводненной мочажины (п = 5);
-
- под прикрытием кладонии (не нависает над гнездом) и багульника на бугре, соседствуют другие бугры с кустарничково-лишайниковым покровом и сильно обводненная мочажина с путницей (n = 1);
-
- полуоткрьггое гнездо, устроенное на бугре в окружении кладонии, багульника и берёзы карликовой среди других бугров с КуСтарнИЧКОБО-ДИШаЙНИКОБЫМ покровом в отдалении аг открытой воды, площадь межбутровых понижении около гнезда незначительна (П= 1)1
-
- полуоткрытое гнездо, устроенное на склоне бугра В окружении кладонии, багульника и брусники около сильно обводненной мочажины в 50 ем от открытой воды (п = 1);
-
- открытое гнездо, устроенное на сфагнуме. на склоне в нижней части бугра с редкой берёзой карликовой и хамедафне среди других бугров с кустарничково-лишайниковым или сфагновым покровом, со-
- седствует сильно обводненная мочажина с пушицей (п=1)
Гнезда М. f. bema размещались в следующих мик-ростациях:
-
- на мхе рода полигрихум, под прикрытием (навесом) сухих и зеленых побегов злаков на иван-чайном злаковом лугу (п= 1);
-
- на глинистой почве под прикрытием (навесом) сухих и зеленых побегов злаков у обошгны дороги (и = 1);
-
- в ямке под прикрытием прошлогодних злаков на лугу' с мятликом и одуванчиком (п = 1);
-
- на залежи среди лугов покосов и осиново-березовых колок (п = 1):
-
- на гари в понижении среди осоки под прикрытием пу^гка сухой травы недалеко от кустов ивняка (п = 1);
-
- на кочкарном осоковом луту ПОД хорошим утфы-тием сухих обгоревших побегов тростника в 10 м от живых растений (n = 1);
-
- на кочкарном ОСОКОВОМ лугу ПОД Прикрытием сухих стеблей осоки в 25 м от тростника (и = 1);
-
- на кочкарном сыром осоковом лугу, истоптанном коровами в 15 м от ивняка (и = 1).
Таким образом, в соответствии с предпочитаемыми гнездовыми стациями подвиды берингийской (жёлтой) и жёлтой трясогузок устраивали гнезда средн растительности и субстрата, свойственных строго определенным биогеоценозам. Предпочтения птиц конкретных местообитании могут быть связаны с особенностями их экологии и поведения, в частности, с выбором оптимальных условий для кормодобы вающей деятельности [Фионина, 2008 и др.]. Дальнейшее изучение способов размещения гнезд и условий, их определяющих, позволит накопить необходимый объем данных для конкретных выводов и обобщений.
Оологические показатели
Число яиц (птенцов) в полных кладках Л/ г. р!еха составляло 4 (п = 1), 5 (п = 6), б (п = 7) и 7 (и = 1), в кладкахЛ// Ьета — 4 (n = 1), 5 (п = 4) и б (п = 1),
Промеры яиц из всех обнаруженных кладок позволили определить основные оологические показатели у рассматриваемых подвидов (табл. 2. 3\ Ввиду того, что выборочное распределение значений L для М. t. ркта отлично от нормального (подтверждается критерием Шапиро-Уилка. р = 0.012), но достоверность таких проверок не всегда приемлема [Орлов. 2006], дополнительно приводим персентильную характеристику отдельных морфологических признаков (табл. 4). Средние размеры яиц М. l ркха отличаются от таковых М. / Ьета. Использование непарамег-рических критериев Вилкоксона (Манна-Уитни) [Орлов, 2014] и Смирнова [Орлов, 2012] при сопоставлении L, В и V дает следующие результаты: р = 0 ятя всех признаков (критерий Вилкоксона) или р = 0.009 для Ц р = 0.037 ятя В р = 0.0003 ддя V (критерий Смирнова). Выявленные отличия ожидаемы и могут быть характерны не только для близких ВИДОВ, НО и ддя популяций ОДНОГО подвида, населяющих территории несколько отдаленные географически [Куранов 2007; Тарасов, 2011 и др.].
Таблица 2
Длина (L) и максимальный (В) диаметр яиц
|
Показатели |
Ц мм |
В, мм |
||||
|
Lini |
М ± а |
CV |
Liin |
М ± а |
CV |
|
|
XI 1, ркха (п = 74) |
15,60-20.10 |
18.21 ± 1.00 |
0.0547 |
12,78-14,76 |
13.99 ±0.43 |
0.0310 |
|
M._f. beenta (n - 27) |
16,16-18.92 |
17,68 ±0,72 |
0.0408 |
13.04-14.86 |
13.88 ±0.39 |
0.0281 |
Таблица 3
Объем (V) и форма (Sph) яиц
|
Показатели |
V, мм |
Sph, % |
||||
|
Lim |
M ± a |
CV |
Lim |
M ± a |
CV |
|
|
Л/. г plcxa (n = 74) |
1449.98-2171.04 |
1822.48 ± 183.98 |
0.1010 |
71,54-87,18 |
77,02 ±3,68 |
0,0477 |
|
XI, f beema (n = 27) |
1573,12-2069,92 |
1737,41 ± 105.38 |
0.0607 |
69,03-87,25 |
78,66 ±4,45 |
0,0566 |
Таблица 4
Персентильная характеристика линейных размеров и объема яиц
|
Показатели |
XL L pkxu (n = 74) |
XL / beemu (n = 27) |
|
|
L, мм |
Me |
1834 |
1730 |
|
5% |
16,13 |
16,30 |
|
|
25 % |
1738 |
17.46 |
|
|
75 % |
18,96 |
18,30 |
|
|
95 % |
1939 |
1831 |
|
Окончание табл. 4
|
Показатели |
XL L piexa (n = 74) |
XL C beema (n = 27) |
|
|
В, мм |
Me |
13.99 |
13.94 |
|
5% |
13.27 |
13.13 |
|
|
25 % |
13.72 |
13.70 |
|
|
75 % |
14.34 |
14.12 |
|
|
95 % |
14.55 |
14.29 |
|
|
Ч мм3 |
Me |
1834.07 |
1713.81 |
|
5% |
1469.33 |
1619.76 |
|
|
25 % |
1738.85 |
1659.94 |
|
|
75 % |
1942.79 |
1797.69 |
|
|
95 % |
2110.99 |
1890.35 |
|
Накопление данных по размерам кладок и яиц трясогузок анализируемого комплекса в будущем позволит говорить об их морфологических отличиях на популяционном уровне, проводить соответствующие аналогии.
Сроки гнездования
К строительству гнезд М* /. piexa. скорее всего, приступают вскоре после прилета. Птиц со строительным материалом мы наблюдали 28 мая (9-й ключевой участок) и 4 июня (10-й ключевой участок) 2006 г., тогда как стаи из пролетных птиц фиксировали 24 и 26 мая 2006 г. (9-й ключевой участок).
Начало откладки яиц в центральной и северной части ХМАО - Югры (север средней тайги и южная подзональная полоса северной тайги Западной Сибири), по-видимому, приходится на конец мая -первую декаду’ июня. В зависимости от фенологических проявлений конкретного года, эти сроки могут несколько варьироваться. Так, гнездо со средне насиженными яйцами осмотрено 13 июня 2004 г. (3-й ключевой участок). Птица с кормом отмечена 13 июня 2005 г. (7-й ключевой участок). Гнездо с готовыми к вылету птенцами обнаружено 29 июня 2006 г. (1-й ключевой участок). Птенцы с формирующимися маховыми (освобождение опахал на кончиках пеньков) пребывали в гнезде 6 июля 2015 г. (23-й ключевой участок). Хорошо летающих слетков регистрировали в первых числах июля 2006 г. (12-й ключевой участок).
В южной части ЯНАО (северная подзональная полоса северной тайги Западной Сибири) сроки гнездования М. t. piexa несколько смещены и подчинены времени схода снега на этой территории. Гнезда с кладками на начальных стадиях насиживания (п = 6) описывали 9-11 и 13 июня 2008 г. (16-й ключевой участок), неполная кладка зарегистрирована 11 июня 2013 г. (22-й ключевой участок). Яйца средней насиженности обнаружены в гнезде 17 июня 2008 г. (16-й ключевой участок). На 14-м ключевом участке 21 июня 2007 г. зафиксирована кладка из яиц с высокой степенью насиженности, на 16-м ключевом участке такие кладки
(п = 4) попадались с 13 июня 2008 г. Первых птиц с кормом на 14-м ключевом участке начали регистрировать с 23 июня 2007 г., появление птенцов в одном из найденных гнезд 16-го ключевого участка отметили 21 июня 2008 г.
О точных сроках гнездования белоухих желтых трясогузок в отдельные годы на территории Среднего Приобья (средняя тайга Западной Сибири) мы можем судить лишь по тем немногим гнездовым находкам и наблюдениям, которые имеются в нашем распоряжении. Гнездо с насиженными яйцами описано 9 июля 2004 г. (5-й ключевой участок), со средне насиженными яйцами -27 июня 2015 г. (23-й ключевой участок).
Вероятно, в лесостепной зоне Западной Сибири гнездование М. / bee та несколько растянуто. Гнездо с неполной кладкой зарегистрировано здесь 4 июня 2009 г., со средне насиженными яйцами -30 мая 2010 г. Вылупление птенцов отмечено 28 мая 2011 г., а 31 мая 2011 г. найдено гнездо с кладкой на ранней стадии насиживания.
Плотность гнездования М. L piexa
В северной части ХМАО — Югры на плоскобугристых верховых болотах с грядово-мочажинными или грядово-мочажинно-озерковыми комплексами берингийские (жёлтые) трясогузки рассматриваемого подвида были распространены неравномерно. На одних болотах они были обычными или многочисленными, на других - редкими или отсутствовали вовсе.
Прослежено изменение гнездовой плотности М. L piexa на трансформированных участках комплексных верховых болот южной подзональной полосы северной тайги Западной Сибири. Птицы часто селились вдоль дорог и трубопроводов, у пустошей, песчаных отсыпок и вахтовых поселков. Их число здесь, по сравнению с менее нарушенными местообитаниями, возрастало в несколько раз (рис. 2). Так, в районе оз. Кутлопъяунлор (11-й ключевой участок) в третьей декаде июня — начале июля 2006 г. плотность гнездования М. t. piexa на придорожном участке комплексного верхового болота составляла 9.2 пары/км', а на отдаленной и менее нарушенной площадке — 4 пары/км3.
Изменения в составе населения и обилии птиц на трансформированных участках западно-сибирской северной тайги отмечались также другими исследователями [Юдкин и др., 1996; Вартапетов, 1991, 1998 и др ]. Несмотря на то, что болотные биоценозы после нарушения почти полностью восстанавливаются в течение 32 лет по количественным показателям запаса надземной фитомассы и проективного покрытия [Орехов, 2007], преобразование экологических систем местообитания приводит к модификации параметров фитоценозов, отличающихся от исходных, главным образом. составом слагающих его видов [Хорошева. 1985 и др*]* Увеличение числа гнездящихся особей М. t. plexa происходит, по-видимому. за счет появления более привлекательных для их существования стаций (с определенными микрорельефом и растительным покровом). Скорее всего, увеличивается и кормность таких стаций* По нашим наблюдениям, кормление и сбор пищевых объектов М. Г plexa часто осуществлялись на мохово-пушицевых мочажинах*
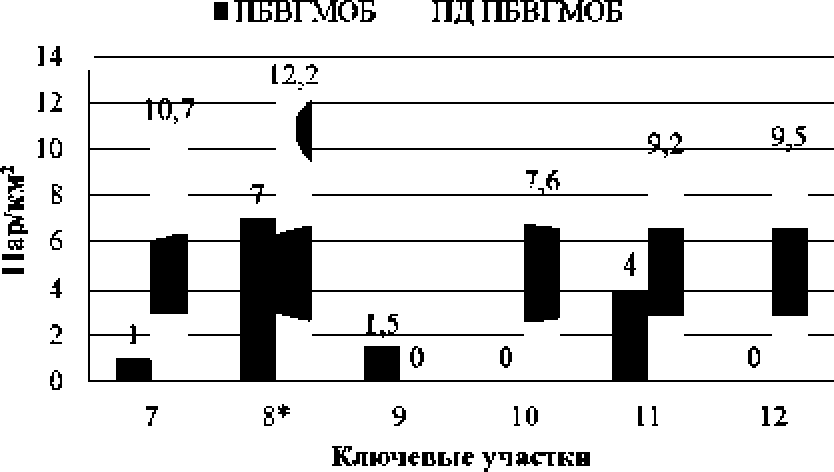
Рис. 2* Изменение гнездовой плотности М. I. plexa на участках местообитаний с разной степенью техногенной нагрузки:
ПБВГМОБ — плоскобугристыс верховые грядо-во-мочажинно-озерковые болота; ЦД ПБВГМОБ — придорожные участки плоскобугристых верховых грядово-мочажинно-озерковых болот; * — на ПБВГМОБ исследования велись в 2005 г., на ПД ПБВГМОБ — в 2006 г.
В ЯНАО на открытых комплексных верховых болотах, багульниково-ерниково-лишайниковых на буграх и осоково-сфагновых в мочажинах и их окраинах это были многочисленные птицы. Их плотность гнездования на различных участках колебалась от 5.5 (14-й ключевой участок) до 12.7-15.9 (13-й ключевой участок) или 10.9-14.6 (16-й ключевой участок) пар/км2. На сходных верховых болотах крайнего севера ХМАО — Югры, в верховьях р. Айкаеган, трясогузки были обычными [Рябицев, Тарасов, 1998]. Таким образам, прослеживается некоторая приуроченность подвида в пределах северной тайги Западной Сибири к местообитаниям северного облика.
Распространение М. f Ьеета
Согласно современным данным, северная граница распространения А/. / Ьеепш проходит в центральной части Западной Сибири, на широте 61°20-40' [Рябицев, 2014]. Исследования, проведенные нами в Среднем Приобье* подтверждают данную схему’. Координаты крайних точек обнаружения птиц на гнездовании — 61O19.9'N, 72°3L2'E (19-й ключевой участок) и 61°20.7,N* 73°25Ч8'Е (21-й ключевой участок)*
Окраска некоторых взрослых особей М» f.
Ьеета
Большинство встреченных нами птиц М» / Ьеета имели характерную для северных трясогузок темно-серую окраску шапочки, сливающуюся с более темной маской (кроющими перьями уха, лба и у глаз), без брови или с просветлением позади и ч>ть выше глаза в виде светлого пятна. Также у нежилого пос. Военто (16-й ключевой участок) в 2008 г. регистрировались птицы (возможно, самки) с очень светлой отчетливой бровью, выступающей далеко позади глаза, серой шапочкой и маской. Были птицы с серыми верхом головы и боками* но без брови. Птица со светлой широкой бровью и светлым пятном позади и ниже глаза отмечена у д. Юган (23-й ключевой участок) в 2015 г. Изредка трясогузки с просветлением над глазом и светлой (серой) раскраской головы отмечались в 2005 г. (8-й ключевой участок).
Выводы
-
1. В Западной Сибири М. t. plexa и М. / Ьеета устраивают гнезда преимущественно в разных микростациях, отличающихся составом слагающих их растительности, субстрата, а также некоторыми другими компонентами биотической и абиотической природы.
-
2. М. t, plexa и М. / Ьеепш отличаются по основным оологическим показателям, таким как длина яиц* их максимальный диаметр и объем
-
3 Начало гнездования М. t. plexa в центральной части Западной Сибири приходится на конец мая — первую половину июня. В юго-западной части Западной Сибири М. / Ьеета гнездится с середины мая.
-
4. В южной подзональной полосе северной тайги Западной Сибири плотность гнездования М. г. plexa на трансформированных участках комплексных верховых болот увеличивается по сравнению с этим же показателем на менее нарушенных участках.
-
5. Среди взрослых представителей М. t. plexa встречаются особи с относительно более светлой (серой) окраской шапочки и маски, а также с выраженной бровью*
Список литературы Некоторые аспекты гнездовой биологии и экологии Motacilla tschutschensis plexa и Motacilla flava beema в Западной Сибири
- Бойко Г.В. Интересные фаунистические находки на Урале и в Западной Сибири//Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург, 1998а. С. 21-24
- Бойко Г.В. Фауна гнездящихся птиц Кондинской низменности//Беркут. 1998б. Т. 7, вып. 1-2. С. 12-18
- Вартапетов Л.Г. Сообщества птиц нефтепромыслов северной тайги Западной Сибири//Орнитологические проблемы Сибири: тез. докл. к конф. Барнаул, 1991. С. 165-166
- Вартапетов Л.Г. Птицы северной тайги Западно-Сибирской равнины. Новосибирск: Наука, 1998. 327 с
- Жуков В. С. Жёлтая Motacilla flava и берингийская M. tschutschensis трясогузки на территории Верх-Тарского нефтяного месторождения (Новосибирская область)//Русский орнитологический журнал: экспресс-вып. 2011. Т. 20, № 691. С. 1899-1906
- Коблик Е.А., Редькин Я.А., Архипов В.Ю. Список птиц Российской Федерации. М.: Товарищество научных изданий КМК, 2006. 288 с
- Коблик Е.А., Архипов В.Ю. Фауна птиц Северной Евразии в границах бывшего СССР: списки видов . 2014. URL: http://zmmu.msu.ru/spec/publikacii/neserijnye-izdaniya/fauna-ptic-stran-severnoj-evrazii
- Куранов Б.Д. Гнездовая биология урбанизированной популяции мухоловки-пеструшки (Ficedula hypoleuca)//Вестник Томского государственного университета. 2007. № 297. С. 192-200
- Меднис А.А., Блум П.Н. Отлов насиживающих уток и их птенцов//Кольцевание в изучении миграций птиц фауны СССР. М., 1976. С. 157167
- Меднис А.А. Определение сроков вылупления утят по плавучести яиц в воде//Русский орнитологический журнал: экспресс-вып. 2002. Т. 11, № 202. С. 1011-1013
- Мянд Р. Внутрипопуляционная изменчивость птичьих яиц = Intrapopulation variation of avian eggs/под ред. Т. Сутта. Талин: Валгус, 1988. 196 с
- Орехов П.Т. Изучение динамики биоценозов северной тайги, нарушенных техногенным воздействием на севере Западной Сибири//Экология: от Арктики до Антарктики: материалы Всерос. конф. молодых ученых. Екатеринбург, 2007. С. 189-194
- Орлов А.И. Прикладная статистика: учебник. М.: Экзамен, 2006. 671 с
- Орлов А.И. Состоятельные критерии проверки абсолютной однородности независимых выборок//Заводская лаборатория. Диагностика материалов. 2012. Т. 78, № 11. С. 66-70
- Орлов А.И. Двухвыборочный критерий Вилкоксона -анализ двух мифов//Политематический сетевой электронный научный журнал Кубанского государственного аграрного университета. 2014. № 104. С. 91-111
- Растительный покров Западно-Сибирской равнины/И.С. Ильина и др.; отв. ред. В.В. Воробьёв, А.В. Белов. Новосибирск: Наука, 1985. 251 с
- Редькин Я.А. Новые представления о таксономической структуре «желтых трясогузок»//Достижения и проблемы орнитологии Северной Евразии на рубеже веков: тр. междунар. конф. Актуальные проблемы изучения и охраны птиц Восточной Европы и Северной Азии. Казань, 2001. С. 150-165
- Редькин Я.А. и др. Вид или не вид? Спорные таксономические трактовки птиц Северной Евразии//Русский орнитологический журнал: экспресс-вып. 2015. Т. 24, № 1237. С. 141-171
- Романов А.Л., Романова А.И. Птичье яйцо: пер. с англ. М.: Пищепромиздат, 1959. 620 с
- Рябицев В.К. Птицы Урала, Приуралья и Западной Сибири: справ.-определитель. Екатеринбург: Изд-во Урал. ун-та, 2008. 640 с
- Рябицев В.К., Тарасов В.В. Птицы верховьев реки Айкаеган//Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург, 1998. С. 165-172
- Рябицев В.К. Птицы Сибири: справ.-определитель: в 2-х т. Москва; Екатеринбург: Кабинетный ученый, 2014. Т. 2. 452 с
- Степанян Л. С. Конспект орнитологической фауны России и сопредельных территорий: (в границах СССР как ист. обл.)/отв. ред. Д.С. Павлов. М.: Академкнига, 2003. 808 с
- Тарасов В.В. Изменчивость размеров яиц белой куропатки//Экология. 2011. № 4. C. 317-320
- Фионина Е.А. Механизмы экологической сегрегации двух совместно обитающих видов трясогузок -белой Motacilla alba и жёлтой M. flava//Русский орнитологический журнал: экспресс-вып. 2008. Т. 17, № 411. С. 527-544
- Хорошева О.В. Изменения растительности верховых болот в результате антропогенного воздействия//Научные доклады высшей школы. Биол. науки. 1985. № 11. С. 84-87
- Юдкин В.А., Вартапетов Л.Г., Козин В.Г. Изменение населения наземных позвоночных при освоении нефтяных и газовых месторождений на севере Западной Сибири//Сибирский экологический журнал. 1996. Т. 3, № 6. С. 573583
- Dunn E.H., Hussell D.J.T., Ricklefs R.E. The Determination of Incubation Stage in Starling Eggs//Bird-Banding. 1979. Vol. 50, № 2. P. 114-120
- Hoyt D.F. Practical Methods of Estimating Volume and Fresh Weight of Birds Eggs//The Auk. 1979. Vol. 96, № 1. P. 73-77
- Mayr E. The interpretation of variation among the yellow wagtails//British Birds. 1956. Vol. 49, № 3. 115-119
- Rahn H., Ar A. The Avian Egg: Incubation Time and Water Loss//The Condor. 1974. Vol. 76, № 2. P. 147-152
- van Oosten H.H., Emtsev A.A. Putative segregation of two Yellow Wagtail taxa by breeding habitat in Western Siberia: possible implications for Motacilla flava taxonomy//Ardea. 2013. Vol. 101, № 1. P. 65-70.
