Некоторые черты эмбриологии Asphodelinetenuior (bieb.) Ledeb. (сем. Asphodelaceae)
 Ledeb. (сем. Asphodelaceae)")
Автор: Шевченко С.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2018 года.
Бесплатный доступ
Приведены результаты изучения особенностей эмбриологии интродуцированного в Никитский ботанический сад узкоареального вида из семейства Asphodelaceae - асфоделины тонкой (Asphodeline tenuior (Bieb.) Ledeb.), эндемика России, в естественных условиях произрастающего на каменистых склонах низкогорий Северного Кавказа. Исследованы процессы цветения, показаны особенности строения элементов цветка и генеративной сферы, определены типы формирования микроспорангия и мужского гаметофита, описано строение семязачатка и зародышевого мешка, показаны некоторые этапы эндоспермо- и эмбриогенеза. Установлено, что в условиях интродукции на Южном берегу Крыма у Asphodeline tenuior могут успешно проходить процессы формирования генеративных структур, опыления, оплодотворения, семяобразования и прорастания семян.
Микроспорангий, мегаспорангий, мужской и женский гаметофиты, эндоспермогенез, эмбриогенез
Короткий адрес: https://sciup.org/147227014
IDR: 147227014 | УДК: 582.573.41:581.3
Some features of embryology Asphodelinetenuior (bieb.) Ledeb. (Asphodelaceae)
The paper presents the results of studying the features of embryology introduced into the Nikita Botanical Gardens of a narrow-aceal species from the family Asphodelaceae - asphodelin fine Asphodeline tenuior (Fisch ex M.Bieb.) Ledeb. - endemic to Russia, in natural conditions growing on stony slopes of the lowlands of the North Caucasus. The flowering processes have been studied, the features of the structure of flower elements and the generative sphere have been revealed, the types of formation of microsporangium and the male gametophyte have been determined, the structure of the ovule and the embryo sac has been described, and some stages of endospermo- and embryo-genesis are shown.It has been established that under the conditions of introduction on the Southern coast of Crimea, Asphodeline tenuior can successfully undergo the processes of formation of generative structures, pollination, fertilization, seed formation and germination of seeds.
Текст научной статьи Некоторые черты эмбриологии Asphodelinetenuior (bieb.) Ledeb. (сем. Asphodelaceae)
Проблема сохранения биоразнообразия в настоящее время является одной из актуальных. При этом особого внимания заслуживают редкие и исчезающие виды растений, поскольку знание особенностей биологии их развития и воспроизведения в условиях природных ареалов позволяет разработать приемы оптимизации размножения и освоения новых территорий [Голубев, 1996; Корже-невский и др., 2004; Ена, 2012].
Одним из редких и сокращающихся в численности видов растений является Asphodeline tenuior (Bieb.) Ledeb. – эндемик России, встречающийся в Краснодарском и Ставропольском краях, в Карачаево-Черкессии и Кабардино-Балкарии. В силу своей малочисленности вид включен в Красную книгу России и региональные Красные книги Краснодарского и Ставропольского краев [Михеев, 2008].
Цель настоящего исследования – выявление особенностей формирования генеративных структур и возможностей воспроизведения A. tenuior вне пределов природного ареала, в частности, в условиях интродукции в Никитском ботаническом саду.
Объекты и методы исследований
Объектом исследований явился эндемик низко-горья Северного Кавказа, травянистый многолетник асфоделина тонкая ( Asphodeline tenuior (Bieb.) Ledeb.), произрастающая в природных условиях на
каменистых склонах и осыпях на высоте до 1000 м н.ур.м. В Никитский ботанический сад вид интродуцирован в начале 80-х гг. ХХ в. Фенологические наблюдения проводили в условиях парковых композиций с учетом рекомендаций А.Н. Пономарева [1960], В.Н. Голубева и Ю.С. Волокитина [1986]. Изучение эмбриологических процессов осуществ- ляли на постоянных препаратах, приготовленных по общепринятым методикам [Ромейс, 1954; Пау-шева, 1990] и окрашенных метилгрюпиронином с подкраской алциановым синим [Шевченко, Ругу-зов, Ефремова, 1986; Шевченко, Чеботарь, 1992]. Парафиновые срезы толщиной 10–12 мкм получали на ротационном микротоме MRD-3000. Анали-
Enamed 2, фото выполняли цифровым фотоаппаратом Canon A 550.
Результаты и их обсуждение
Асфоделина тонкая ( Asphodeline tenuior ) – многолетнее травянистое растение до 40–50 см высотой с простым, слабо ветвистым стеблем (рис. 1, А). Листья тонкие, шиловидные, в условиях интродукции на ЮБК цветет в июне, плодоносит в июле. Цветки расположены в верхней части стебля, собраны в рыхлую кисть, образованную пазушными пучками из 2–3 цветков (рис. 1). Изредка встречаются пучки, состоящие из 5 цветков.
зировали препараты с помощью микроскопа


Рис. 1 . . Фрагменты растений Asphodeline tenuior (А – фрагмент разветвленного стебля; Б – фрагменты соцветий с раскрытыми и увядающими цветками)
Цветение А. tenuior волнообразное, сначала раскрываются цветки нижних пучков соцветия, по одному в пучке, постепенно поднимаясь вверх до середины соцветия; на верхушке его в это время цветки находятся на стадии бутонизации (рис. 1, Б). Затем снова раскрываются следующие цветки (по одному в каждом пучке) пучков в нижней части соцветия. При этом мы наблюдаем пучок, состоящий из маленькой завязи, раскрытого цветка и бутона. Раскрываются цветки к вечеру (в 17–18 часов), и к утру они закрываются и увядают в отличие от А. lutea и A. taurica, у которых цветки раскрываются утром [Шевченко, 2017а, г].
Цветок А. tenuior имеет простой венчиковидный околоцветник, состоящий из 6 сегментов желто-зеленоватого цвета с зеленой полоской посредине сегмента. Полностью раскрытый цветок зигоморфный, 5 его сегментов направлены вверх, а 1 – вниз (рис. 2, А). В завязи формируются септаль- ные нектарники, которые открываются в боковых ее частях.
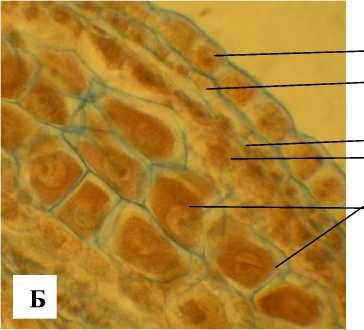
Андроцей состоит из 6 неравных тычинок, 3 внутренние имеют тычиночные нити, которые значительно длиннее внешних. Основания тычиночных нитей расширены и плотно окружают завязь, вследствие чего нектарники недоступны мелким насекомым, а крупные насекомые их достигают хоботком. Вверху тычиночная нить срастается со связником в середине пыльника, входя в своеобразную канавку (рис. 2, Б). Это дает дополнительную возможность для движения раскрывающимся интрорзно пыльникам и привлечения насекомых.
Пыльники 2-тековые, 4-гнездные (рис. 3, А). Стенка микроспорангия развивается центростремительно по однодольному типу и, сформированная, состоит из эпидермы, эндотеция, одного среднего слоя и секреторного тапетума (рис. 3, Б). Клетки эпидермы довольно крупные, с четко выраженными ядром и ядрышком. Средний слой эфемерный и к началу мейоза дегенерирует. Тапетум является производным вторичной париетальной ткани, клетки его одно- и двуядерные. Споро-

генная ткань представлена двумя-тремя слоями (рис. 3, Б).

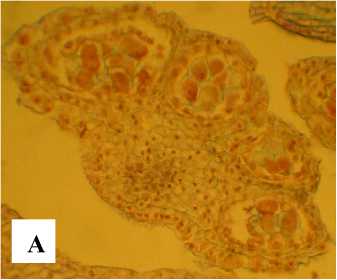
Рис. 2 . . Раскрытый цветок А. tenuior (А) и фрагмент его тычинки (Б)
ЭЭН
СС Т
СТ
Рис. 3 . . Поперечный срез пыльника А. tenuior (А) и фрагмент стенки микроспорангия на стадии сформированной спорогенной ткани (Б):
Э – эпидерма; ЭН – эндотеций; СС – средний слой; Т – тапетум; СТ – спорогенная ткань
К началу мейоза стенка микроспорангия дезинтегрирует, ткань тапетума частично распадается на отдельные клетки, вокруг клеток спорогенной ткани откладывается каллоза (рис. 4, А). Тетрады микроспор образуются сукцессивно (рис. 4, Б). На стадии тетрады микроспор стенка микроспорангия представлена эпидермой, эндотецием и тапетумом (рис. 5, А). Клетки тапетума одно- и двуядерные. В период дифференцирующего митоза и на стадии 2клеточного пыльцевого зерна стенка микроспорангия состоит из крупных клеток эпидермы и фиброзного эндотеция (рис. 5, Б).
Э ЭН КТ МС
МС
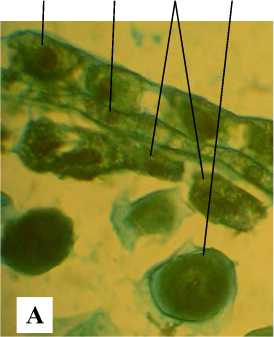
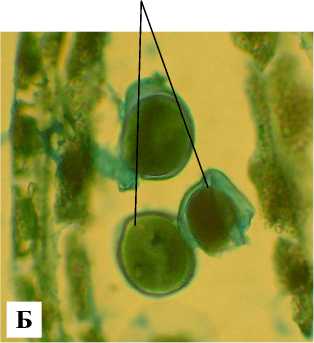
Рис. 4 . Фрагменты микроспорангиев на стадии I деления мейоза (А) и II деления мейоза (Б): Э – эпидерма; ЭН – эндотеций; КТ – клетки тапетума; МС – микроспороцит
Следует обратить внимание на то, что цитоплазма и ядро в клетках фиброзного эндотеция вакуолью смещены к внутренней стенке, направленной к центру микроспорангия. Зрелые пыльцевые зерна 2-клеточные. Типы формирования мужских
генеративных структур подобны таковым у A. lutea и A. taurica [Шевченко, 2017а; Шевченко, Багри-кова, Крайнюк, 2017; Шевченко, Крайнюк, Багри-кова, 2017].
Э ЭН КТ ТМ
ПЗ ФЭ Э
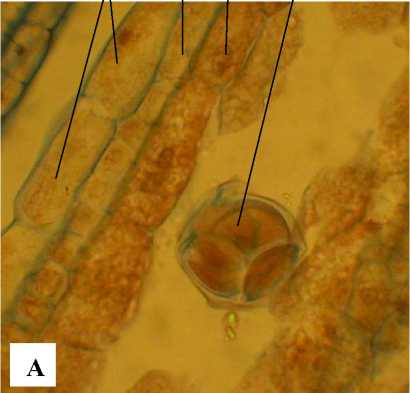
Рис. 5 . Фрагменты микроспорангия на стадиях тетрады микроспор (А) и 2-клеточных пыльцевых зерен (Б):
Э – эпидерма; ЭН – эндотеций; КТ – клетки тапетума; ТМ – тетрада микроспор; ПЗ – пыльцевые зерна; ФЭ – фиброзный эндотеций
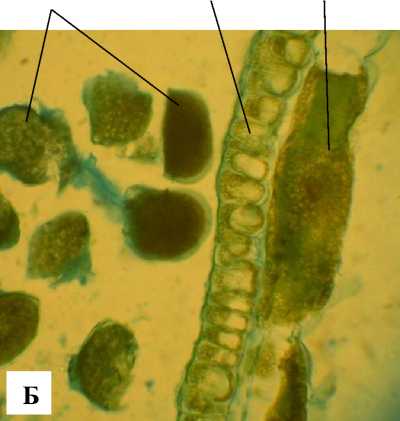
Гинецей А. tenuior синкарпный, завязь состоит из трех плодолистиков. Семязачаток анатропный, битегмальный, крассинуцеллятный (рис. 6). Микропиле образовано внутренним интегументом, который в основном 2-слойный, в микропилярной зоне его клетки активно делятся и формируют оперкулум. В халазальной зоне у основания семязачатка развивается истинный ариллус (рис. 6). Внутренний интегумент в своем развитии опережает наружный (рис. 7, 8, А), образованный тремя слоями клеток. В основании фуникулуса формируется фуникулярный обтуратор. Халазальная зона нуцеллуса сильно развита, ниже зародышевого мешка, как и у A. taurica [Шевченко, 2017б], развивается постаментоподиум. Эпидермальные клетки нуцеллуса, делясь, в микропилярной зоне образуют нуцеллярный колпачок (рис. 6).
Археспорий закладывается в субэпидермальной зоне примордия, делясь, образует париетальную и спорогенную клетки. Затем в процессе деления париетальных клеток формируется микропилярная зона нуцеллуса, а спорогенная клетка дифференцируется в мегаспороцит. Мейотическое деление мегаспороцита приводит к образованию тетрады мегаспор. Функционирующей является халазаль-ная мегаспора, из которой и развивается зародышевый мешок Polygonum-типа (рис. 8, Б). Подоб-
ное развитие женской генеративной сферы наблю-
дали мы и у A. lutea [Шевченко, 2017в].
НИ
НК
ВИ
ЗМ
АР
Рис. 6 . Общий вид семязачатка А. tenuior :
НИ – наружный интегумент, НК – нуцеллярный колпачок, ВИ – внутренний интегумент, ЗМ – зародышевый мешок, АР – ариллус
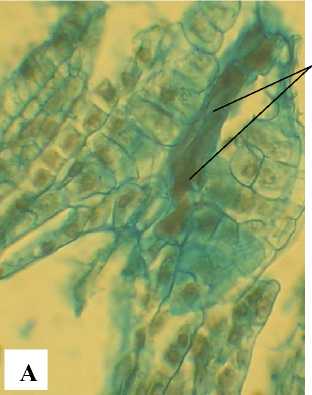
Зрелый зародышевый мешок 8-ядерный, 7клеточный. Яйцевой аппарат состоит из 2 синергид с нитчатым аппаратом и яйцеклетки с крупным ядром и ядрышком (рис. 9). Антиподы состоят из 3 клеток, расположенных Т-образно в халазальной части зародышевого мешка. Полярные ядра сливаются до оплодотворения. Оплодотворение порогамное. Пыльцевая трубка после прорастания по тканям пестика через микропиле достигает зародышевого мешка, проходит через одну из синергид и изливает свое содержимое (рис. 10, А).
О
НИ
ЭН
ЯА
ЗМ
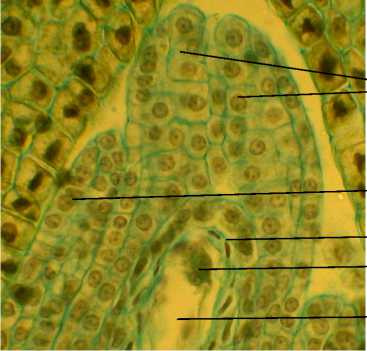
Рис. 7 . Фрагмент семязачатка А. tenuior :
НИ – наружный интегумент, О – оперкулум;
ЭН – эпидерма нуцеллуса; ЯА – яйцевой аппарат , ЗМ – зародышевый мешок
После слияния спермия с ядром центральной клетки зародышевого мешка, образующееся первичное ядро эндосперма сначала находится в центре, затем мигрирует в халазальную зону и располагается над антиподами (рис. 10, Б).
Как и у многих других видов семейства Aspho-delaceae [Поддубная-Арнольди, 1982], эндосперм у А. tenuior гелобиального типа, однако по классификации О.П. Камелиной [1994] он тубифлораль-ный. Первичное ядро делится с образованием двух клеток разного размера: халазальной маленькой и крупной микропилярной. Множественные деления ядра в микропилярной клетке образуют многоядерную структуру (рис. 11, А), ядра которой располагаются в узких тяжах цитоплазмы, в то время как в халазальной клетке проходит обычно одно деление с образованием двух ядер, выполняющих функцию гаустория (рис. 11, Б).
Следует отметить, что полностью раскрывается цветок у А. tenuior в вечернее время и к утру он увядает. Пыльники вскрываются еще в полураскрытом бутоне, а лопасти рыльца и в раскрытом цветке сомкнуты. То есть и на этом этапе репродуктивного цикла проявляется протерандрия, когда мужские генеративные структуры в своем развитии опережают женские. Рыльце готово к приему пыльцы в темное время суток, и пыльца может быть перенесена на рыльце пестика ночными насекомыми.
При увядании сегменты цветка снова смыкаются, пестик выдается за пределы околоцветника, что создает дополнительную возможность для опыления, если оно еще не произошло. Образующиеся в результате эффективных процессов опыления и оплодотворения плоды – трехгнездные кожистые коробочки на довольно длинных плодоножках с сочленением, ритмы их завязывания соответствуют ритмам цветения (рис. 12, А). Семена трехгранные, до 5 мм длиной (рис. 12, Б).
НИ ВИ ПК МС
ПК ДМ ЧЯ ЗМ
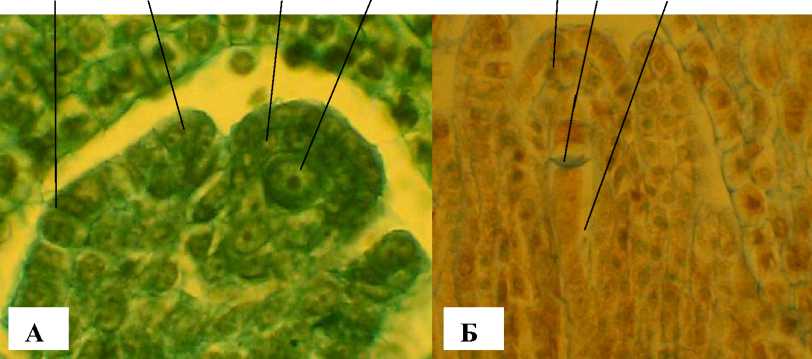
А
Б
Рис. 8 . Начальные стадии формирования семязачатка и зародышевого мешка А. tenuior :
НИ – наружный интегумент, ВИ – внутренний интегумент, ПК – париетальные клетки; МС – мегаспороцит; ДМ – дегенерирующие мегаспоры; ЧЯ ЗМ – четырехядерный зародышевый мешок
НК СНА
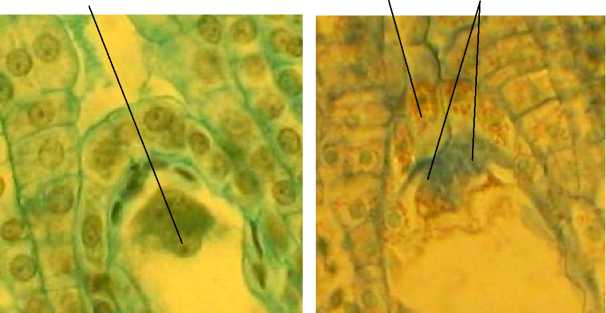
Рис. 9 . Фрагменты семязачатков А. tenuior :
Я – яйцеклетка; НК – нуцеллярный колпачок; СНА – синергиды с нитчатым аппаратом
ПТ
ПЯЭ
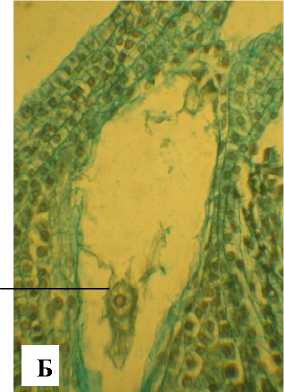
Рис. 10 . Фрагменты семязачатков А. tenuior в период оплодотворения:
ПТ – пыльцевая трубка; ПЯЭ – первичное ядро эндосперма
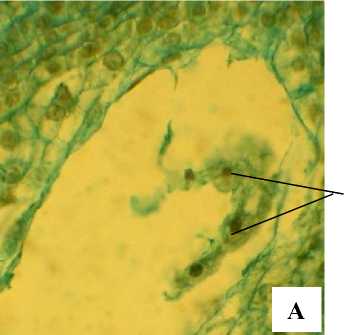
ХГ
ЯЭ
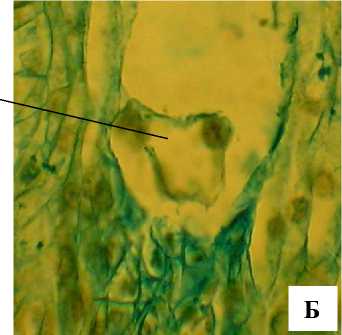
Рис. 11 . . Отдельные этапы формирования эндосперма А. tenuior :
ЯЭ – ядра эндосперма, ХГ – халазальный гаусторий
Плодов обычно завязывается довольно много, но это зависит не только от нормально развитых и функционирующих генеративных структур, но и от погодных условий и наличия насекомых-опылителей, обеспечивающих эф- фективный процесс опыления. Размножается А. tenuior семенами и вегетативно путем деления корневищ взрослых растений. Данные характеристики А. tenuior подобны таковым у A. lutea и A. taurica, произрастающих в горном Крыму
[Шевченко, 2017а,б,в; Шевченко, Крайнюк, Багрикова, 2017].

Рис. 12. . Фрагменты генеративных побегов А. tenuior с плодами разной степени зрелости
Заключение
Таким образом, по основным эмбриологическим признакам А. tenuior сходна с другими видами семейства Asphodelaceae: центростремительное формирование стенки микроспорангия (однодольный тип), сукцессивный тип образования тетрады микроспор, 2-клеточные пыльцевые зерна, анатропный, битегмальный, крассинуцеллятный с оперкулумом и ариллусом семязачаток, Polygonum-тип зародышевого мешка, наличие поста-ментоподиума, порогамное оплодотворение, эндосперм гелобиального (тубифлорального) типа и развитие халазального гаустория. Формирующиеся элементы генеративной сферы, ритмы и продолжительность цветения, образующиеся плоды и семена могут обеспечить воспроизведение А. tenuior при его интродукции в Крым, а декоративность в период цветения позволяет рекомендовать данный вид для использования в садово-парковом строительстве.
Список литературы Некоторые черты эмбриологии Asphodelinetenuior (bieb.) Ledeb. (сем. Asphodelaceae)
- Голубев В.Н. Биологическая флора Крыма. Ялта: НБС-ННЦ, 1996. 126 с
- Голубев В.Н., Волокитин Ю.С. Методические рекомендации по изучению антэкологических особенностей цветковых растений. Функционально-экологические принципы организации репродуктивной структуры. Ялта, 1986. 36 с
- Ена Ан.В. Природная флора Крымского полуострова. Симферополь: Ор1анда, 2012. 232 с
- Камелина О.П. Новый подход к классификации типов тапетума//Эмбриология цветковых растений. Терминология и концепции. СПб.: Мир и семья, 1994. Т. 1. С. 56-60
- Корженевский В.В., Багрикова Н.А., Рыфф Л.Э., Бондарева Л.В.. Редкие и находящиеся под угрозой исчезновения виды и проблемы их охраны в Севастополе (Крым)//Экология, фитоценология и оптимизация экосистем: сб. науч. тр. Никит. бот. сада. 2004. Т. 123. С. 196-210
- Михеев А.Д. Асфоделина крымская Asphodeline taurica (Pall. ех Bieb.) Endl.//Красная книга Российской Федерации (растения и грибы). М., 2008. 856 с
- Паушева З.П. Практикум по цитологии растений. М.: Колос, 1990. 283 с
- Поддубная-Арнольди В.А. Характеристика семейств покрытосеменных растений по цитоэм-бриологическим признакам. М.: Наука, 1982. 352 с
- Пономарев А.Н. Изучение цветения и опыления растений//Полевая геоботаника. М.; Л., 1960. Т. 2 С. 9-19
- Ромейс Б. Микроскопическая техника. М.: Изд-во иностр. лит-ры, 1954. 718 с
- Шевченко С.В. Формирование мужских генеративных структур Asphodeline lutea (L.) Rchb. (семейство Asphodelaceae)//Бюллетень Государственного Никитского ботанического сада. 2017а. Вып. 124. С. 97-103
- Шевченко С.В. Особенности репродуктивных процессов некоторых редких видов флоры Крыма//Актуальные проблемы ботаники и охраны природы: сб. науч. статей. Симферополь, 2017б. С. 320-330
- Шевченко С.В. К вопросу о сохранении Asphodeline lutea (L.) Rchb. и Asphodeline taurica (Pall.) Endl. в Крыму//Современные технологии в изучении биоразнообразия и интродукции растений: сб. материалов Междунар. науч. конф. Ростов-н/Д.; Таганрог, 2017в. С. 95-98
- Шевченко С.В. О цветении и опылении асфодели-ны крымской (Asphodeline taurica (Pall.) Endl. (сем. Asphodelaceae)//Актуальные проблемы биологии и экологии: материалы междунар. на-уч.-практ. конф. Грозный; Махачкала, 2017г. С. 143-147
- Шевченко С.В., Багрикова Н.А., Крайнюк Е.С. Некоторые морфобиологические особенности и распространение Asphodeline taurica (Pall. ex Bieb.) Endl. (сем. Asphodelaceae) в Крыму//Роль ботанических садов и дендрариев в сохранении, изучении и устойчивом использовании разнообразия растительного мира. Минск, 2017. С. 330-333
- Шевченко С.В., Крайнюк Е.С., Багрикова Н.С. Структура популяций Asphodeline lutea (L.) Rchb. и Asphodeline taurica (Pall.) Endl. в Горном Крыму//Научные записки заповедника «Мыс Мартьян». 2017. Вып. 8. С. 52-59
- Шевченко С.В., Ругузов И.А., Ефремова Л.М. Методика окраски постоянных препаратов метиловым зеленым и пиронином//Бюллетень Государственного Никитского ботанического сада. 1986. Вып. 60. С. 99-101
- Шевченко С.В., Чеботарь А.А. Особенности эмбриологии маслины европейской (Olea europaea L.)//Сборник научных трудов Никитского ботанического сада. 1992. Т. 113. С. 5261
