Некоторые использующиеся в исследованиях лиганды ASIC каналов

Автор: Храмов А.П., Уграицкая В.М.
Журнал: Форум молодых ученых @forum-nauka
Статья в выпуске: 5 (69), 2022 года.
Бесплатный доступ
В этой статье приведены обзорные сведения о веществах, воздействующих и модулирующих активность ASIC каналов.
Asic-каналы, лиганды, токсины, модуляция активности
Короткий адрес: https://sciup.org/140293199
IDR: 140293199 | УДК: 577.25
Some ASIC channel ligands used in research
This article provides an overview of substances that affect and modulate the activity of ASIC channels.
Текст научной статьи Некоторые использующиеся в исследованиях лиганды ASIC каналов
Вступление
Наиболее сильно и избирательно на различные ASIC-каналы действуют соединения, полученные из различных ядов. На сегодняшний день открыто около десятка модуляторов активности ASIC-каналов, добытых из различных ядов.
Причём, на ASIC1 преимущественно действуют вещества из ядов змей и пауков, а на ASIC3 преимущественно действуют вещества из ядов морских анемонов. [5]
PcTx1.
Пожалуй, наиболее изученным модулятором активности ASIC каналов можно считать PcTx1.
Псалмотоксин - PcTx1 (также известный как π-TRTX-Pc1a) это пептид, выделенный из яда Тринидадского шевронного тарантула Psalmopoeus cambridgei (Escoubas et al., 2000). Этот белковый яд способен избирательно блокировать работу ASIC1а канала
Псалмотоксин состоит из 40 аминокислот. Трехмерная структура состоит из компактного дисульфидно-связанного ядра, от которого отходят три петли и N- и C-концы. Основным элементом структуры является трехцепочечный антипараллельный β-лист. [10]
Этот яд может быть отнесён к ингибиторам цистинового узла (ICK) белка.
Немало подобных ядов было выделено из улиток и скорпионов, но среди ICK-токсинов псалмотоксин - единственный известный пептид, действующий на гомомерные каналы ASIC1. [9]
При связывании PcTx1 с ASIС1а увеличивает сродство этого протон-чувствительного канала к H+. При этом канал переходит в десенситизированное состояние при привычной концентрации протонов (pH = 7,4). При десенситизации канал связан со своим лигандом, с Н+, но не способен пропускать через себя ионы.
PcTx1 был выделен как первый высокомощный и селективный ингибитор гомомерного ASIC1a (крысиный ЛД50 ~1 нМ при pH 7,45 и ЛД50 ~3 нМ при pH 7,2). Впоследствии было показано, что PcTx1 также ингибирует токи в коэкспрессирующих ASIC1a/2b клетках (ЛД50 мыши ~3 нМ) и гетеротоки в гетеротоках. ~3 нМ) и гетеромерных крысиных каналов ASIC1a/2a (на ~35-85% при 50 нМ в состоянии при pH 6,95 в зависимости от состава тримера) (Joeres et al., 2016; Sherwood et al., 2011).
ЯМР-структура PcTx1 ( Рис.1 ) , которая была впервые раскрыта в 2003 году и уточнена в 2011 году, показала наличие ингибиторного цистеинового узла (ICK) и то, что доминирующая β-волосковая петля (петля 4), в которой представлены некоторые из ключевых фармакофорных остатков, является очень гибкой (Escoubas et al., 2003; Saez et al., 2011).
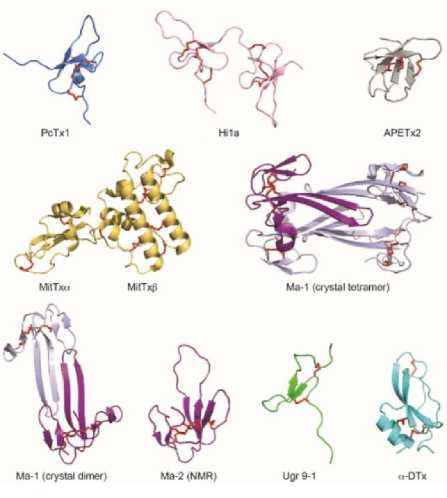
Рисунок 1. Модулирующие активность ASIC каналов пептидные вещества..
(А) Аминокислотная последовательность пептидов сгруппированных с гомологами, где они присутствуют. Различия в последовательностях обозначены серыми рамками, а активные сайты -треугольниками над фармакофорными остатками. а активные сайты - треугольниками над фармакофорным остатком. (B) Трехмерная структура PcTx1 (PDB код 2KNI; (Saez et al., 2011)), APETx2 (PDB код 2MUB; [11], Hi1a (PDB-код 2N8F; (Chassagnon et al., 2017)), Ma-1 (PDB-коды 5DU1, 5DZ5; (Mourier et al., 2016)), Ma-2 (PDB код 2MFA; (Schroeder et al., 2014)), MitTx (PDB код 4NTW; (Baconguis et al., 2014)), Ugr 9-1 (PDB код 2LZO; (Osmakov et al., 2013)), и α-DTx (PDB-код 1DTX; (Skarżyński, 1992)).
Hm3a
Помимо псалмотоксина, учёные смогли выделить также другой интересный модулятор активности ASIC каналов - π-TRTX-Hm3a (Hm3a). Этот модулятор был выделен из яда Тоголезского звездно-полосатого тарантула (Heteroscodra maculata)
Структура этого яда отличается от структуры псалмотоксина на пять аминокислот и короче на три остатка на С-конце.
Hm3a чрезвычайно стабилен в человеческой сыворотке в течение 48 часов (оставалось ~87%), в то время как PcTx1 оставалось в течение того же времени ~35%. Hm3a практически не разрушается в течение 48 часов при 55 °C, а PcTx1 разрушается (~24%).
Превосходная стабильность Hm3a по сравнению с PcTx1, в сочетании с аналогичной фармакологией пептидов делает Hm3a потенциально более привлекательным инструментом для исследований ASIC in vivo. [5]
Hi1a
Этот модулятор активности ASIC каналов был найден относительно недавно при анализе транскриптома ядовитых желез австралийского воронковидного паука (Hadronyche infensa).
N-концевой участок этого соединения имеет высокое сродство в строение с PcTx1 (62% идентичности). При этом Hi1a почти вдвое длиннее псалмотоксина и состоит из 75 аминокислотных остатков.
С-концевой домен Hi1a тоже очень схож с строением псалмотоксина (50% идентичности). [4]
В исследованиях 2010 года было показано, что Hi1a является очень сильным ингибитором ASIC1а (ЛД50 400-500 мкМ) и имеет два последовательно расположенных ICK мотивов, аналогично агонисту TRPV1 DkTх [2]
В отличие от PcTx1 и Hm3a, которые влияют на десенситизацию ASIC1 в рН-зависимой и быстро обратимой манере, Hi1a селективно ингибирует активацию ASIC1a в медленно обратимой манере и оказывает незначительное влияние на ASIC1b до 1 мкМ.
APETx2
APETx2 - это пептид, состоящий из 42 аминокислотных остатков. Его выделили из экстрактов морского анемона Anthopleura elegantissima.
Этот пептид стал первым мощным селективным ингибитором ASIC3 субъединиц - ЛД50 63 нМ для гомомерных rASIC3 и 0,1-2 мкМ для гетеромерных rASIC3-содержащих каналов [8].
APETx2 ингибирует типичный pH-индуцированный ток; устойчивый ток, вызванный pH 7,0 ASIC3 [7], а также щелочно-индуцируемый ток hASIC3.[6] При этом, APETx2 не способен ингибировать сильный ток, который наступает при снижении pH ниже 5,0. [8]
В состав APETx2 входят: компактное гидрофобное ядро, которое состоит из четырёхцепочечного β-листа, в котором находятся три дисульфидные связи [3] [11]
В 2012 году было показано, что фармакофор APETx2 для ASIC3 представляет собой непрерывную поверхность, состоящую из петель 2 и 4 и N-конца (рис.2)
Рисунок 2.
Пока ещё не были в полной мере раскрыты молекулярные детали взаимодействия между APETx2 и ASIC3. Предполагается, что APTEx2 смещает видимое сродство канала к протонам [1]
Так как APETx2 является самым мощным лигандом из доступных учёным для изучения ASIC3 субъединиц, однозначное определение места его связывания и механизма действия очень важно для последующей разработки более селективных аналогов и для дальнейшего понимания структуры и функции ASIС3 каналов in vivo.
Список литературы Некоторые использующиеся в исследованиях лиганды ASIC каналов
- Baron, A., Diochot, S., Salinas, M., Deval, E., Noël, J., Lingueglia, E., 2013. Venom toxins in the exploration of molecular, physiological and pathophysiological functions of acid-sensing ion channels. Toxicon 75, 187-204.
- Bohlen, C. J., Priel, A., Zhou, S., King, D., Siemens, J., Julius, D., 2010. A bivalent tarantula toxin activates the capsaicin receptor, TRPV1, by targeting the outer pore domain. Cell 141, 834-845
- Chagot, B., Escoubas, P., Diochot, S., Bernard, C., Lazdunski, M., Darbon, H., 2005. Solution structure of APETx2, a specific peptide inhibitor of ASIC3 proton-gated channels. Protein Sci. 14, 2003-2010 ;
- Chassagnon, I. R., McCarthy, C. A., Chin, Y. K., Pineda, S. S., Keramidas, A., Mobli, M., Pham, V., De Silva, T. M., Lynch, J. W., Widdop, R. E., Rash, L. D., King, G. F., 2017.
- Cristofori-Armstrong, B., & Rash, L. D. (2017). Acid-sensing ion channel (ASIC) structure and function: Insights from spider, snake and sea anemone venoms. Neuropharmacology, 127, 173–184. doi:10.1016/j.neuropharm.2017.
- Delaunay, A., Gasull, X., Salinas, M., Noel, J., Friend, V., Lingueglia, E., Deval, E., 2012. Human ASIC3 channel dynamically adapts its activity to sense the extracellular pH in both acidic and alkaline directions. Proc. Natl. Acad. Sci. USA 109, 13124-13129.
- Deval, E., Noël, J., Gasull, X., Delaunay, A., Alloui, A., Friend, V., Eschalier, A., Lazdunski, M., Lingueglia, E., 2011. Acid-sensing ion channels in postoperative pain. J. Neurosci. 31, 6059-6066.
- Diochot, S., Baron, A., Rash, L. D., Deval, E., Escoubas, P., Scarzello, S., Salinas, M., Lazdunski, M., 2004. A new sea anemone peptide, APETx2, inhibits ASIC3, a major acid- sensitive channel in sensory neurons. EMBO J. 23, 1516-1525.
- Escoubas P, Bernard C, Lambeau G, Lazdunski M, Darbon H (July 2003). "Recombinant production and solution structure of PcTx1, the specific peptide inhibitor of ASIC1a proton-gated cation channels". Protein Science. 12 (7): 1332–43. doi:10.1110/ps.0307003. PMC 2323924. PMID 12824480.
- Escoubas P, De Weille JR, Lecoq A, Diochot S, Waldmann R, Champigny G, Moinier D, Ménez A, Lazdunski M (August 2000). "Isolation of a tarantula toxin specific for a class of proton-gated Na+ channels". The Journal of Biological Chemistry. 275 (33): 25116–21.
- Jensen, J. E., Cristofori-Armstrong, B., Anangi, R., Rosengren, K. J., Lau, C. H., Mobli, M., Brust, A., Alewood, P. F., King, G. F., Rash, L. D., 2014. Understanding the molecular basis of toxin promiscuity: the analgesic sea anemone peptide APETx2 interacts with acid-sensing ion channel 3 and hERG channels via overlapping pharmacophores. J. Med. Chem. 57, 9195-9203
