Некоторые морфометрические параметры Avena sativa и содержание в нем пролина при засолении NaCl на фоне кислой и щелочной реакции почвенной среды

Автор: Четина Оксана Александровна, Устинова Ксения Игоревна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 3, 2019 года.
Бесплатный доступ
Изучены некоторые физиолого-биохимические показатели в листьях овса посевного Avena sativa L. при засолении на фоне разных рН-уровней почвенной среды. Отмечено негативное влияние этих факторов на всхожесть семян и морфометрические показатели овса посевного. Кислотный и щелочной уровень рН почв подавляли рост растений овса в высоту в среднем на 20% по сравнению с контролем и вызывали сокращение листовой поверхности на 30%. При засолении на фоне разных уровней рН эти показатели сокращались практически в 2 раза. Максимальная масса растений овса зафиксирована на щелочной среде, как с засолением, так и без него. Увеличение массы при общей тенденции снижения других ростовых показателей, скорее всего, связано с накоплением воды, что, в свою очередь, может быть обусловлено повышением количества осмолитов в клетках. Количество засоляющих ионов Na+ и Cl- в листьях овса увеличивается во всех вариантах опыта с засолением почвы. Максимальное накопление зафиксировано при действии щелочного стресса, что, вероятно, связано с изменением проницаемости мембран. Содержание пролина в надземных органах овса посевного возрастает при засолении почвы во всех вариантах рН-уровней. На кислом и щелочном фоне наблюдается снижение количества пролина относительно нейтральной среды, как в условиях засоления, так и без него.
Засоление, кислотный стресс, щелочной стресс, адаптация, пролин, морфометрические показатели
Короткий адрес: https://sciup.org/147227097
IDR: 147227097 | УДК: 581.1 | DOI: 10.17072/1994-9952-2019-3-345-352
Some morphometric Avena sativa parameters and content of proline in it at salinization on the background of acid and alkaline reaction in сespitose-podsolic soil
Some physiological and biochemical parameters were studied in the leaves of Avena sativa L. at salinization and different рН-levels of the soil environment. The negative influence of these factors on seed germination and morphometric parameters of oats is noted. Acid and alkaline pH levels of soil suppressed plant growth in height by 20% compared with the control and reduced leaf surface by 30%. At salinization against the background of different pH levels these indicators were reduced almost twice. The maximum mass of oat plants is fixed on an alkaline medium, both with salinity and without salinity. The increase in mass with a decrease in other growth indicators, most likely due to the accumulation of water. This, in turn, may be due to an increase in the number of osmolytes in the cells. The amount of salting Na+ and Cl- ions in the leaves of oats increases in all variants of the experiment with salinization of the soil. The maximum accumulation was recorded under the action of alkaline stress, which is probably due to changes in membrane permeability. The content of proline in the leaves of oats increases at salinization of the soil in all variants of pH levels. On the acidic and alkaline background, a decrease in the amount of proline in a relatively neutral environment is observed, both under and without salinity.
Текст научной статьи Некоторые морфометрические параметры Avena sativa и содержание в нем пролина при засолении NaCl на фоне кислой и щелочной реакции почвенной среды
Засоление является одним из основных абиотических факторов, действие которого приводит к угнетению роста и развития растений, снижению их продуктивности. Глобальное потепление и опустынивание земель в значительной степени способствуют расширению масштабов почвенного засоления, которое в ближайшей перспективе может только прогрессировать. Повышение устойчи-
вости растений к засолению пахотных земель является одной из ключевых задач современного сельскохозяйственного производства [Аверина и др., 2014].
В почвах накопление солей может сопровождаться увеличением щелочности, техногенное засоление развивается и на фоне кислой реакции среды [Еремченко и др., 2017]. В свою очередь, для большинства сельскохозяйственных культур, в особенности для молодых растений, требуется благоприятная среда, близкая к pH 6.5. Снижение рН внешней среды на 1 единицу уменьшает рН цитоплазмы примерно на 0.1 единицы [Wilkinson et all., 1989]. С другой стороны, при увеличении рН внешней среды происходит осаждение ионов фосфора и металлов, увеличивается поглощение неорганических анионов, нарушается ионный баланс [Yang et all., 2008]. Поэтому и кислотный, и щелочной уровень рН корневой среды изменяет рН клеток и тканей. Уровень рН среды существенно влияет на фотосинтез, генерацию активных форм кислорода и усиление перекисного окисления липидов, проницаемость мембран и т.д. [Bhuyan et all., 2019; Shi, Sheng, 2005; Guo et all., 2015].
Одним из признанных маркеров устойчивости растения считается уровень пролина. Многочисленные публикации свидетельствуют в пользу его протекторного действия при стрессах. Свободный пролин при стрессе обладает полифункциональ-ным биологическим эффектом, который проявляется не только в осморегуляторной, но также и в антиоксидантной, энергетической и других функциях, обеспечивающих поддержание клеточного гомеостаза и его переход в новое адаптивное состояние [Кузнецов, Шевякова, 1999; Zhu, 2001; Shi, Sheng, 2005; Lv et al., 2013; Guo et al., 2015; Tomar, 2018; Bhuyan et al., 2019].
Таким образом, актуальность изучения механизмов адаптации растений к совместному воздействию засоления и кислотности / щелочности корневой среды, в том числе, с участием универсального осмопротектора – пролина, обусловлена тем, что эти факторы сочетаются в природных и техногенных почвах. В современной физиологии растений недостаточно изучена ответная реакция растений на комбинированное воздействие этих факторов корневой среды, особенно, сочетание засоления и кислотности [ Shi, Sheng, 2005; Liu, Shi, 2010; Lv et al., 2013 ] .
Целью данной работы являлось изучение морфометрических параметров Аvena sativa и оценка уровня накопления в нем пролина при засолении почвы NaCl на фоне кислой и щелочной почвенной среды.
Объект и методы исследования
Ответную реакцию овса посевного Avena sativa L. (несортовой) изучали в модельном эксперименте. Растения выращивали в контейнерах с дерново-подзолистой почвой (элювиальный горизонт) в различных сочетаниях pH-уровня и засоления NaCl. Исходная реакция почвы – 4.9 рН. Нейтральная среда почвенной среды до pH=7.1 создавалась путем добавления CaCO3, для подщелачивания до 8.2 использовали Na2CO3. Засоление NaCl создавалось в процентном отношении к сухому весу почвы. Контролем в опыте считали вариант без засоления с 7.1 рН.
Варианты опыта:
|
Без засоления |
С засолением, % |
|
|
pH=4.9 |
pH=4.9+0.5 NaCl |
pH=4.9+1 NaCl |
|
pH=7.1 |
pH=7.1+0.5 NaCl |
pH=7.1+1 NaCl |
|
pH=8.2 |
pH=8.2+0.5 NaCl |
pH=8.2+1 NaCl |
У растений в течение 8 дней с момента появле- ния всходов определяли всхожесть, длину и массу надземной части, площадь листовой пластины [по: Аникиев, Кутузов, 1961] в 30-кратной повторности. На 8-й день после появления всходов листья растений фиксировали при 105оС и досушивали при 60оС. В сухой массе изучили количество ионов Na+ – методом пламенной фотометрии, Cl- – мер-курометрическим методом, пролин – методом Бейтса – в 3-кратной биологической повторности. Данные обработаны с применением пакета статистических программ Stadia 8.0 и использованием методов описательной статистики, сдви-га/положения, непараметрической корреляции, регрессионного и дисперсионного анализов.
Результаты и их обсуждение
Максимальная всхожесть семян овса посевного наблюдалась в контрольном варианте нашего эксперимента. В варианте без засоления почвы при щелочной и кислой реакции среды всхожесть была на 8 и 17% ниже контроля соответственно. Значительное снижение всхожести наблюдалось в вариантах с засолением NaCl 0.5%; при этом на фоне нейтральной среды всхожесть была ниже в 6.6 раза, а при кислой и щелочной средах – в 13 раз. В вариантах с засолением почвы NaCl 1% семена не взошли.
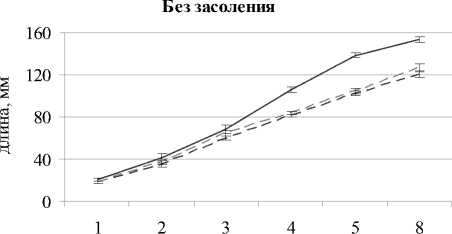
Известно, что интенсивность роста является интегральным показателем состояния растений, отражающим функциональные и метаболические изменения в растении. При наблюдении за изменением длины надземной части растений установлено, что при кислой и щелочной среде без засоления достоверно снижалась скорость роста овса. На 8-й день после появления всходов высота растений была ниже 18% при щелочной среде и на 22% – при кислой среде относительно контроля (рис. 1).
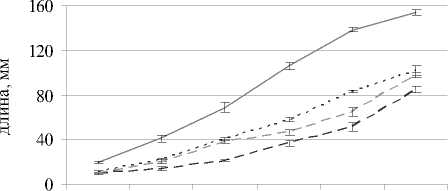
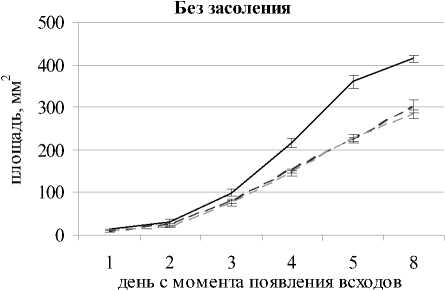
Во всех вариантах засоление 0.5% NaCl способствовало значимому снижению высоты растений в каждый срок наблюдений по сравнению с контролем. На 8-й день после появления всходов в варианте с засолением при рН 7.1 длина надзем- ной части была на 34% меньше, чем без засоления; а при щелочной среде с засолением – ниже на 36% и при кислой с засолением – ниже на 45% по сравнению с незасоленными вариантами. Таким образом, сильнее всего подавлялся рост овса при сочетании засоления и кислой реакции корневой среды, что может быть связано не только с действием NaCl и кислотностью среды, но и с дополнительным негативным влиянием токсичных катионов, таких как A13+ и Mn2+ [Shavrukov, Hirai Yo, 2016]. В вариантах без засоления почвы при кислой и щелочной среде наблюдалось значимое уменьшение площади листовой пластинки во все сроки наблюдений относительно варианта с нейтральной средой (рис. 2); на 8-й день после появления всходов площадь листа овса посевного была ниже на 27 и 32% соответственно.
день с момента появления всходов
Засоление NaCl 0,5%
день с момента появления всходов
pH=7,1 (контроль) pH=4,9 + 0,5% NaCl pH=4,9
pH=7,1 (контроль) pH=8,2
pH=7,1 + 0,5% NaCl pH=8,2 + 0,5% NaCl
pH=4,9
pH=7,1 pH=8,2
Рис. 2. Площадь листовой пластинки Avena sativa L. при разном уровне pH почвы и засолении
Рис. 1 . Длина надземной части Avena sativa L. при разном pH-уровне почвы и засолении
день с момента появления всходов pH=7,1 pH=4,9 + 0,5%NaCl
pH=7,1 + 0,5%NaCl pH=8,2 + 0,5%NaCl
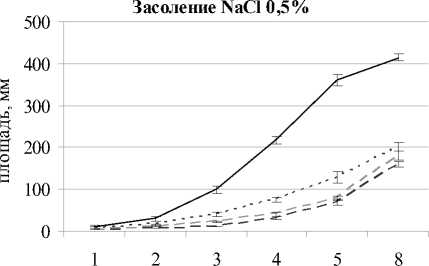
Во всех вариантах с засолением 0.5% NaCl в каждый срок наблюдений отмечали значимое снижение площади листовой пластинки растений. На 8-й день после появления всходов в варианте с засолением при рН 7.1 площадь листа была меньше на 51%, при кислой среде – на 62%, при щелочной – на 57% относительно незасоленных вариантов.
Медленный рост позволяет растениям выживать при стрессе, так как освобождает множество ресурсов (строительных блоков и энергии) для реализации защитной программы, что, по-видимому, необходимо в условиях неблагоприятной реакции корневой среды. В исследованиях С. Yang [2009] отмечено уменьшение роста как кор- ней, так и побегов ячменя в щелочных условиях, снижение содержания воды и фотосинтетических пигментов. Аналогичное ингибирование роста при щелочном стрессе было также зарегистрировано у растений томата Solanum lycopersicum [Wang et al., 2011]. Кислотный стресс тормозит рост корней, что, в свою очередь, приводит к снижению поглощения воды и питательных веществ. Основными ограничениями для роста растений на кислых минеральных почвах являются токсичные концентрации минеральных элементов, таких как Al3+, H+, и/или низкая доступность минеральных питательных веществ [Marschner, 1991]. В исследованиях M.H.M.B. Bhuyan и др. [2019], у проростков пше- ницы, выращенных при рН 4.0 и 8.5, наблюдалось снижение содержания биомассы, воды и хлорофилла, в отличие от растений нейтральной среды.
Полученные данные по росту овса посевного в условиях засоления соответствуют мнению многих авторов о том, что солевой стресс оказывает отрицательное воздействие на ранних стадиях онтогенеза проростков, ингибируя процессы митотического цикла и тем самым замедляя развитие растений [Зайцева, 2009; Zhang, 2010; Mahmood, 2011; Shahri et all., 2012; Yousufinia et all., 2013; Pascaru et all., 2014; Гордеева, Татауров, 2017]. Подавление клеточного деления и особенно растяжения при действии неблагоприятных условий приводит к задержке роста растений, которую рассматривают как защитную реакцию. Устойчивые виды растений быстрее адаптируются к стрессу и восстанавливают рост [Белозерова, Боме, 2014].
Данные по массе надземной части овса посевного в вариантах опыта представлены на рис. 3. Максимальная масса растений зафиксирована в условиях щелочной реакции почвенной среды, как без засоления, так и при засолении 0.5% NaCl. Масса растений в вариантах с кислой и нейтральной средой не имела значимых различий (рис. 3).
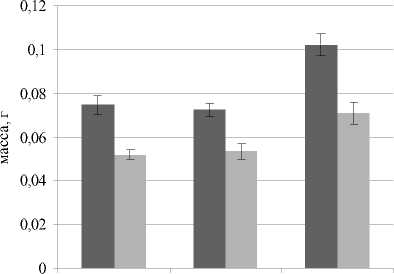
pH=4,9 pH=7,1 pH=8,2
■ без засоления ■ засоление 0,5% NaCl
Рис. 3 . Масса надземной части Avena sativa L. (для одного растения) при разном уровне pH почвы и засолении
Увеличение массы растений на щелочной почве при общей тенденции снижения других ростовых показателей, скорее всего, связано с накоплением воды, что, в свою очередь, может быть обусловлено накоплением осмолитов в клетках. Так, в исследованиях китайских ученых [Shi, Sheng, 2005] отмечено, что содержание пролина, лимонной кислоты и Na+ увеличивалось в побегах подсолнечника с повышением уровня засоления и щелочности корневой среды. Лимонная кислота может выполнять функцию осмолита, а также регулировать рН в клетке. Rui Guo и др. [2010] также зафиксировали накопление органических кислот в проростках пшеницы при действии щелочного стресса.
При проведении эксперимента установлено значимое снижение массы растений на засоленных вариантах относительно незасоленных, при ней- тральной среде с засолением масса растений овса посевного была ниже на 40, при рН 4.9 и 8.2 – на 30%.
В условиях засоления торможение роста растений связывают также и с накоплением токсичных ионов Na+ и Cl- [Munns et al., 2006]. В ходе исследования обнаружено, что на почве без засоления с разным рН-уровнем больше всего ионов Cl- накапливается на щелочном фоне (на 70% больше контрольного варианта) (рис. 4). Между контролем и вариантом с кислой средой статистически достоверных различий не обнаружено.
При кислой и щелочной реакции среды и одновременном засолении 0.5% NaCl наблюдалась аккумуляция ионов хлора в листьях; при рН 4.9 их было больше в 1.5 раза, а при рН 8.2 – в 2.3 раза, что, по-видимому, связано с нарушением проницаемости мембраны. Кислотный и щелочной стрессы усиливают генерацию активных форм кислорода и перекисное окисление липидов, что отражается на проницаемости мембран [Yang, 2009; Bhuyan et al., 2019].
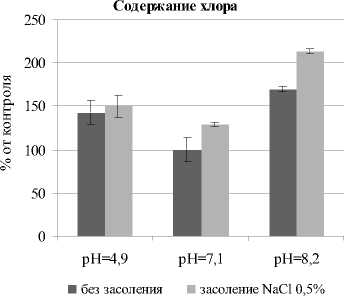
Содержание натрия
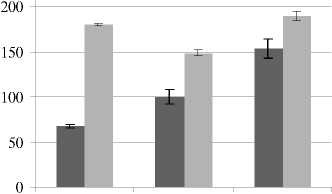
pH=4,9 pH=7,1
pH=8,2
■ без засоления ■ засоление 0,5% NaCl
Рис. 4 . Содержание свободных ионов Na+ и Cl- в надземных органах Avena sativa L. при разном уровне pH почвы и засолении
Различный pH-уровень почвенной среды влияет и на количество натрия в надземной части овса. Статистически значимые различия обнаружены между контролем и вариантами pH 4.9 и 8.2; в варианте с кислой почвой количество натрия снижено на 32, с щелочной – выше на 54%.
При засолении почвы увеличение содержания ионов натрия в листьях овса наблюдалось при всех уровнях рН: при нейтральной среде – в 1.5 раза, при кислой и щелочной – в 1.8–1.9 раз соответственно. Известно, что состояние гиперосмотического шока заставляет растение поддерживать снижающееся осмотическое давление клеток путем поглощения катионов натрия и сопровождающего аниона хлора. Повышение концентрации этих ионов приводит к нарушению метаболических процессов из-за возникающего ионного дисбаланса и токсичности натрия [Палладина, 1999]. Кроме того, по данным Rui Guo и др. [2010] повреждение клеток, вызванное щелочным стрессом, может вызывать массовый приток Na+, что приводит к серьезному дефициту отрицательного заряда. Растения, пытаясь восстановить осмотический баланс, синтезируют органические кислоты в качестве контрионов. Для устранения ионного дисбаланса, возможно, накапливаются и анионы Cl- в листьях овса при щелочной почвенной среде.
В большинстве исследований феномен накопления пролина связывается с устойчивостью к стрессу [Франко, Мело, 2000]. Результаты нашего эксперимента показали, что содержание пролина в надземных органах овса посевного в вариантах без засоления при рН 4.9 и 8.2 достоверно ниже контрольного варианта (в 2.5–3 раза) (рис. 5).
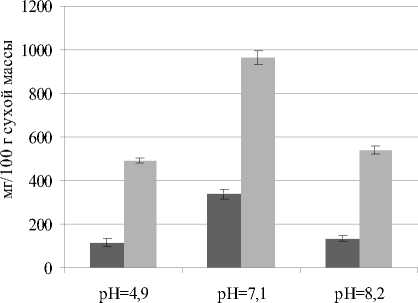
без засоления засоление 0,5% NaCl
Рис. 5 . Содержание пролина в надземной части Avena sativa L. при разном уровне pH почвы и засолении
Максимальным содержанием пролина в надземных органах овса посевного отличались варианты с засолением. В варианте с нейтральной почвой его количество выше в 3 раза относительно контроля, что свидетельствует об осморегулятор-ной роли этой аминокислоты, и наиболее успешной адаптацией растений к засолению в условиях нейтральной реакции корневой среды.
В вариантах с засоленной почвой и при сочетании с кислой и щелочной реакцией среды количество пролина было больше, чем в контроле с соот- ветствующей рН (на 45 и 60%), но оставалось все-таки ниже, чем на нейтральной засоленной почве (практически в 2 раза).
Во многих исследованиях отмечается накопление пролина как при солевом, так и при кислотнощелочном стрессах. Так, в работах Shi, Sheng [2005] наблюдалась концентрация пролина в листьях подсолнечника при солевом и щелочном стрессах. Повышение содержания пролина в растениях риса прослежено при щелочном стрессе [Lv et al., 2013]. Кислые условия корневой среды способствовали накоплению пролина в листьях томатов [Tomar, 2018]. По Bhuyan и др. [2019] аккумуляция пролина в проростках пшеницы при кислотном и щелочном стрессах могла обеспечить защиту от окислительного повреждения и поддержать водный баланс растений.
В нашем эксперименте отличительной особенностью воздействия щелочной и кислой почвенной среды было снижение уровня пролина в листьях овса относительно нейтрального варианта. Роль пролина в рН-регуляции отмечалась в отдельных работах [ Zhu, 2001; Shi, Sheng, 2005; Lv et all., 2013; Tomar, 2018; Bhuyan et all., 2019 ] , но в нашем случае накопления пролина не происходит, а, следовательно, его роль в отсутствие засоления оказалась малозначительной. Снижение уровня пролина может также указывать и на повышенное его потребление в условиях стресса [Сергеева и др., 2011].
Заключение
Таким образом, результаты наших опытов показали негативное влияние засоления NaCl, кислой и щелочной почвенной среды на всхожесть семян и морфометрические показатели овса посевного. В вариантах со щелочной и кислой почвой рост растений овса был в среднем ниже на 20%, а листовая поверхность – на 30% относительно варианта с нейтральной почвой. При сочетании засоления с кислой и щелочной средой эти показатели состояния растений уменьшались практически еще в 2 раза.
Максимальная масса растений овса отмечена в варианте со щелочной почвой, как при засолении NaCl, так и без него. Увеличение массы при общей тенденции снижения других ростовых показателей, скорее всего, связано с накоплением воды, что, в свою очередь, может быть обусловлено аккумуляцией осмолитов в клетках.
Количество засоляющих ионов Na+ и Cl- в листьях овса увеличивалось во всех вариантах опыта с засолением почвы. Максимальное накопление этих ионов зафиксировано в условиях засоления и щелочной среды, что может быть связано с дополнительным внесением щелочной соли натрия, а также с нарушением проницаемости мембран.
Содержание пролина в надземных органах овса посевного возрастало при засолении почвы во всех вариантах рН-уровня, что связано с осмопротек-торными функциями этой аминокислоты. При выращивании овса на кислой и щелочной почве наблюдалось снижение количества пролина относительно варианта с нейтральной средой; следовательно, значение этого многофункционального протектора в адаптации овса посевного к отдельному действию неблагоприятной почвенной рН не прослежено.
Список литературы Некоторые морфометрические параметры Avena sativa и содержание в нем пролина при засолении NaCl на фоне кислой и щелочной реакции почвенной среды
- Аверина Н.Г. и др. Роль метаболизма азота в формировании солеустойчивости растений ячменя // Физиология растений. 2014. Т. 61, № 1. С. 106-113.
- Аникиев В.В., Кутузов Ф.Ф. Новый способ определения площади листовой поверхности у злаков // Физиология растений. 1961. Т. 8, № 3. С. 375-377.
- Белозерова А.А., Боме Н.А. Изучение реакции яровой пшеницы на засоление по изменчивости морфометрических параметров проростков // Фундаментальные исследования. 2014. № 12-2. С. 300-306.
- Гордеева И.В., Татауров В.П. Сравнительный анализ воздействия кратковременного и длительного солевого стресса на всхожесть и морфометрические параметры Secale cereale и Triticum durum // Международный научно-исследовательский журнал. 2017. № 9-2 (63). С. 6-10.
- Еремченко О.З., Митракова Н.В., Шестаков И.Е. Природно-техногенная организация почвенного покрова территории воздействия солеотвалов и шламохранилищ в Соликамско-Березниковском экономическом районе // Вестник Пермского университета. Сер. Биология. 2017. Вып. 3. С. 311-320.
- Зайцева Р.И. Желнакова Л.И., Никитина Н.С. Характеристика солеустойчивости кормовых культур в начальной фазе вегетации при засолении чернозема хлоридом натрия // Бюллетень почвенного института им. В.В. Докучаева. 2009. № 63. С. 25-40.
- Кузнецов В.В., Шевякова Н.И. Пролин при стрессе: биологическая роль, метаболизм, регуляция // Физиология растений. 1999. Т. 46, № 2 С. 321-336.
- Палладина Т.А. Роль протонных насосов плазмалеммы и тонопласта в устойчивости растений к солевому стрессу // Успехи современной биологии. 1999. Т. 119, № 5. С. 451-461.
- Сергеева Л.Е., Бронникова Л.И., Тищенко Е.Н. Содержание свободного пролина как показатель жизнедеятельности клеточной культуры Nikotiana tabacum L. при стрессе // Биотехнология. 2011. Т. 4, № 4. С. 87-94.
- Франко О.Л., Мело Ф.Р. Осмопротекторы: ответ растений на осмотический стресс // Физиология растений. 2000. Т. 47, № 1. С. 152-159.
- Bhuyan M.H.M.B. et al. Unraveling Morphophysiological and Biochemical Responses of Triticum aestivum L. to Extreme pH: Coordinated Actions of Antioxidant Defense and Glyoxalase Systems // Plants. 2019. Vol. 8, № 1. P. 24.
- Guo R. et al. Comparative metabolic reponses and adaptive strategies of wheat (Triticum aestivum) to salt and alkali stress // ВМС Plant Biology. 2015. Vol. 15. P. 170.
- Guo R. et al. Effect of saline and alkaline stress on germination, seedling growth, and ion balance in wheat // Agronomy Journal. 2010. Vol. 102, № 4, P. 1252-1260.
- Liu J., Shi D.C. Photosynthesis, chlorophyll fluorescence, inorganic ion and organic acid accumulations of sunflower in responses to salt and salt-alkaline mixed stress // Photosynthetica. 2010. Vol. 48, № 1. Р. 127-134.
- Lv B.S. et al. Differences in Growth and Physiology of Rice in Response to Different Saline-Alkaline Stress Factors // Agronomy Journal. 2013. Vol. 105, № 4. Р. 1119-1128.
- Mahmood K. Salinity tolerance in barley (Hordeum vulgare, L.): effects of varying NaCl, K+/Na+ and NaHCO3 levels on cultivars differing in tolerance // Pakistan Journal of Botany. 2011. Vol. 43, № 3. P. 1651-1654.
- Marschner H. Mechanisms of adaptation of plants to acid soils // Plant Soil. 1991. Vol. 134. P. 1-20.
- Munns R., James R., Lauchli A. Approaches to increasing the salt tolerance of wheat and other cereals // Journal of Experimental Botany. 2006. Vol. 57, № 5. P. 1025-1043.
- Pascaru A., Giorgievici A., Gaman C. Sodium chloride effect on rye (Secale cereale) // Journal of Horticulture, Forestry and Biotechnology. 2014. Vol. 18, № 4. P. 147-150.
- Shahri S., Tilaki G., Alizadeh M. Influence of salinity stress on seed germination and seedling early growth stages of three Secale species // The Asian and Australian Journal of Plant Science and Biotechnology. 2012. Vol. 12. P. 28-31.
- Shavrukov Y., Hirai Y. Good and bad protons: genetic aspects of acidity stress responses in plants // Journal of Experimental Botany. 2016. Vol. 67, № 1. Р. 15-30.
- Shi D., Sheng Y. Effect of various salt-alkaline mixed stress conditions on sunflower seedlings and analysis of their stress factors // Environmental and Experimental Botany. 2005. Vol. 54, № 1. P. 8-21.
- Tomar P. Estimation of some Biochemical Parameters in Lycopersicon lycopersicum (L.) cv. Damyanti in Response to Acid Rain // Voyager. 2018. Vol. 9, № 1. Р. 34-41.
- Wang X. et al. Physiological responses and adaptive strategies of tomato plants to salt and alkali stresses // Scientia Horticulturae. 2011. Vol. 130. P. 248-255.
- Wilkinson R.E., Duncan R.R. Sorghum seedling growth as influenced by H+, Ca2+, and Mn2+ concentrations // Journal of plant nutrition. 1989. Vol. 12. P. 1379-1394.
- Yang C.W. et al. Comparison of effects of salt and alkali stresses on the growth and photosynthesis of wheat // Photosynthetica. 2008. Vol. 46. P. 107-114.
- Yang, C. et al. Comparative effects of salt stress and alkali-stress on the growth, photosynthesis, solute accumulation, and ion balance of barley plants // Photosynthetica. 2009. Vol. 47. P. 79-86.
- Yousufinia M. et al. The effect of NaCl on the growth and K+ content of barley (Hordeum vulgare, L.) cultivares // Annals of Biological Research. 2013. Vol. 4, № 1. P. 80-85.
- Zhang H. et al. The effects of salinity and osmotic stress on barley germination rate: sodium as an osmotic regulator // Annals of Botany. 2010. Vol. 106. P. 1027-1035.
- Zhu J.K. Plant salt tolerance // Trends in Plant Science. 2001. Vol. 6, № 2. P. 66-71.
