Некоторые особенности транспортной функции клеточной стенки растений Арктики

Автор: Теребова Елена николаевнА., Марковская Евгения федоровнА., Шмакова Наталья юрьевнА.
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 8 (137), 2013 года.
Бесплатный доступ
Исследуются ионообменная способность (количество функциональных групп) и набухание клеточной стенки листа растений Betula nana L., Salixpolaris Walenb., Dryas octopetala L. и Cassiope tetragona (L.) D. Don., произрастающих на острове Западный Шпицберген. Установлено, что клеточная стенка листа растений Арктики является катионообменником, который представлен сходным составом ионообменных групп (аминогруппы, карбоксильные и фенольные ОН-группы) с клеточными стенками растений тундровой зоны. У всех видов наибольший процент в структуре клеточной стенки листа приходится на карбоксильные группы гидроксикоричных кислот и фенольные ОН-группы, которые входят в состав соединений фенольной природы. Особенностью растений Арктики является увеличение в 2-3 раза количества ионообменных групп каждого типа и высокие коэффициенты набухания клеточной стенки листа по сравнению с видами других климатических зон. Делается предположение, что высокие значения ионообменной способности и коэффициента набухания клеточной стенки у всех исследованных видов способствуют более активному водному потоку по системе апопласта и усилению метаболических процессов в клеточной стенке у растений в условиях высоких широт.
Клеточная стенка, ионообменные свойства, растения арктики
Короткий адрес: https://sciup.org/14750550
IDR: 14750550 | УДК: 577+581.17
Some features of cell wall transport function in Arctic plants
Ion exchange of the leaves' cell walls in species Betula nana L., Salix polaris Walenb., Dryas octopetala L. and Cassiope tetragona (L.) D. Don. growing on the island of Western Spitsbergen were investigated. It was found out that the cell wall of arctic plants is a cation exchanger, which is comparable to the functional groups (amine groups, carboxyl groups and phenolic OH-groups) with the cell wall of plants from tundra zone. In all investigated species, the highest percentage in the structure of the cell wall was reported for the carboxyl groups of hydroxycinnamic acids and phenolic OH-groups, which are a part of the phenolic compound. In comparison with species from other climatic zones leaves' cell walls of arctic plants have 2-3 times higher amount of ion exchange groups of all types as well as a higher value of swelling coefficient. It was proposed that the high values of the ion-exchange capacity and swelling coefficient of the cell wall of all studied species contribute to greater water flow system by the apoplast and enhance metabolic processes in the cell wall of the plants growing in conditions of high latitudes.
Текст научной статьи Некоторые особенности транспортной функции клеточной стенки растений Арктики
Клеточная стенка является одной из наиболее важных структур растительной клетки, которая первой включается в адаптационные процессы, реагируя на условия произрастания. Ее сложный состав и активная функциональная деятельность [3] свидетельствуют о перестройках в зависимости от условий среды. Экстремальные условия Арктики диктуют особые требования к структуре, в том числе и к клеточной стенке тех видов, которые произрастают в этих условиях. Однако работ, выполненных на клеточной стенке растений, произрастающих и адаптированных к высоким широтам, практически нет. В задачу настоящего исследования входило изучение ионообменных способностей клеточной стенки листа растений, произрастающих в условиях Арктики.
МЕСТО, ОБЪЕКТЫ И МЕТОДИКА ИССЛЕДОВАНИЯ
Работа выполнена в зоне тундровой растительности внутренних фьордов Западного Шпицбергена в 2009–2010 годах. Климатические
особенности включают: короткий вегетационный период (40–70 дней), продолжительность которого определяется временем схода снега в локальных местообитаниях; низкая среднесуточная температура (8 °C) и круглосуточный полярный день с 19 апреля по 24 августа [4].
Объектами исследования были виды естественной флоры архипелага Шпицберген – Betula nana L. (сем. Betulaceae ), Salix polaris Walenb. (сем. Salicaceae ), Dryas octopetala L. (сем. Rosaceae ) и Cassiope tetragona (L.) D. Don. (сем. Ericaceae ). Все виды собраны в районе залива Колсбей Западного Шпицбергена.
Для определения состава и количества ионогенных групп в клеточных стенках листа использовали метод потенциометрического титрования, количество групп оценивали по значению сорбционной емкости (S, мкмоль/г сух. массы клет. ст.), весовой коэффициент набухания клеточных стенок определяли гравиметрическим методом [10], расчет константы ионизации – по [6; 7].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Транспортная функция растительной клеточной стенки состоит в передвижении веществ по апопласту и связана с образованием специальных структур клеточной стенки, которые отлагаются в период ее формирования и направляют и регулируют транспортные потоки [3], формирующиеся во время роста и развития растений. Клеточная стенка – слабый катионообменник, который способствует передвижению небольших водорастворимых молекул, куда входят вода, газы и различные низкомолекулярные вещества. Все они испытывают слабое сопротивление со стороны структур клеточной стенки (до 10–15 %).
Исследования показали, что в структуре клеточных стенок листа исследованных растений Арктики выявлено четыре типа ионообменных или функциональных групп (табл. 1), которые включают одну анионообменную аминогруппу с константой ионизации (рКа ~ 3–4) и три катионообменные группы: карбоксильные группы ά-D-полигалактуроновой кислоты (СООН ПГУК) (рКа ~ 5–6), карбоксильные группы ги-дроксикоричных кислот (СООН ГКК) (рК а ~ 7–8) и фенольные ОН-группы (рКа ~ 9–10). Количество и состав ионообменных групп клеточных стенок листа изученных растений сходен с составом клеточной стенки листа и корня других видов древесных [8] и травянистых растений [14; 6]. Общее количество ионообменных групп в структуре клеточной стенки листа растений определяет максимальные значения сорбционной емкости клеточной стенки. Эти группы выполняют транспортную функцию и принимают участие в реакциях обмена.
Таблица 1 Содержание ионообменных групп в клеточной стенке листа растений Арктики
|
Тип группы |
Кол-во групп, AS мкмоль на г сухой массы клет. стенки |
|||
|
Betula nana |
Dryas octopetala |
Salix polaris |
Cassiope tetragona |
|
|
Аминогруппы |
1200 ± 50 |
500 ± 50 |
450 ± 30 |
400 ± 20 |
|
СООН ПГУК* |
1900 ± 30 |
1100 ± 50 |
450 ± 20 |
600 ± 40 |
|
СООН ГКК** |
2500 ± 90 |
3000 ± 60 |
1700 ± 90 |
750 ± 30 |
|
Фенольные ОН-группы |
2700 ± 80 |
2600 ± 60 |
550 ± 25 |
950 ± 70 |
|
Общее кол-во катионообменных групп |
7100 ± 100 |
6700 ± 80 |
2700 ± 50 |
2300 ± 50 |
|
Общее кол-во функциональных групп |
8300 ± 100 |
7200 ± 90 |
3150 ± 50 |
2700 ± 50 |
Примечание. * – карбоксильные группы ά-D-поли-галактуроновой кислоты; ** – карбоксильные группы ги-дроксикоричных кислот.
Анализ количества аминогрупп клеточной стенки в листьях исследуемых растений показал, что максимально высокие значения (до 1200 мкмоль/г сух. массы клет. ст.) получены для Betula nana – редкого вида для Западного Шпицбергена, для остальных видов этот показатель значительно ниже и для у Cassiope tetragona – широко распространенного, особенно на каль-ценосных почвах вида, было показано количество групп – 400 мкмоль/г сух. массы клет. ст. Ионообменные аминогруппы клеточной стенки входят в состав структурных белков и ферментов, содержание которых может достигать 15 % от массы клетки. Как отмечает Т. А. Горшкова, они представляют непрерывный ряд от щелочных, минимально гликозилированных белков (пролин-богатые) до кислых, высокогликози-лированных протеогликанов (арабиногалактоновые белки) [3]. Основной аминокислотой в гидролизате клеточной стенки является оксипролин, содержание которого может достигать 40 % [13]. Ферментная система клеточной стенки представлена гидролазами, которые гидролизуют полисахариды, приводя к изменениям физических свойств клеточной стенки и ионного обмена. В формировании поверхностных слоев эпидермиса (кутина и суберина), которые присутствуют почти на всех листьях растений Арктики, участвуют трансацилазы.
В группу катионообменников входит карбоксильная группа ά-D-полигалактуроновой кислоты (СООН ПГУК), максимальные значения которой обнаружены в клеточной стенке листа у Betula nana, высокие значения имеют виды Salix рolaris и Dryas octopetala , а низкие величины отмечены у Cassiope tetragona. Основным полимером пектиновых веществ является поли-галактуроновая кислота, полимеры которой находятся в пиранозной форме. Пектиновые вещества – природные ионообменники, основной их функцией является регуляция действия ферментов за счет создания подвижной системы локального изменения рН и электрического заряда. Так, именно полигалактуроновая кислота отвечает за отрицательный заряд клеточной стенки [3].
Второй катионообменной группой клеточной стенки листа является карбоксильная группа гидроксикоричных кислот (СООН ГКК). Исследуемые виды по количеству СООН ГКК в клеточной стенке листа объединились в две группы: наибольшие значения обнаружены у Betula nana, Dryas octopetala, во вторую группу вошли Salix рolaris и Cassiope tetragona с более низкими значениями. Гидроксикоричные кислоты являются составляющей суберина, кутина и пектиновых полисахаридов. Кроме того, такие оксикоричные кислоты, как n-кумаровая, феруловая и синаповая, относят к лабильной метаболической составляющей клеточной стенки [3]. Благодаря образованию диферулатов феруловая кислота может выполнять в клеточных стенках структурную функцию, связывая между собой полисахариды, уменьшая растяжимость клеточных стенок [9], [10], [11]. Фенольные кислоты клеточных стенок могут в качестве восстановителей участвовать в окислительно-восстановительных процессах, способствовать утилизации активных форм кислорода. Они также могут выполнять функции регуляторов роста и играть сигнальную роль при взаимодействии растений с микроорганизмами [9], [19].
Третья катионообменная группа – фенольные ОН-группы могут входить в состав компонентов вторичной клеточной стенки – лигнина и суберина. К видам с большим количеством этих групп в клеточной стенке относятся Betula nana, Dryas octopetala (2700 и 2600 мкмоль/г сух. массы кл. ст. соответственно); почти в 2 раза меньше у Cassiope tetragona и еще меньше у Salix рolaris. Лигнин как структурная стабильная компонента клеточной стенки составляет до 40 % ее сухой массы. Фенольные соединения – продукты вторичного метаболизма, выполняют множество функций в растительном организме, в том числе защитную, как от биогенных, так и от абиогенных факторов.
Общее количество ионообменных групп клеточной стенки листа у видов Арктики оказалось в 2–3 раза больше, чем в клеточной стенке хвои Pinus sylvestris [8], листьев видов рода Betula , произрастающих в более южных районах Европейского Севера России (по неопубликованным данным Н. А. Галибина). Значение общего количества всех исследованных функциональных групп клеточной стенки листа показало, что оно изменяется от максимального у Betula nana (8300 мкмоль/г сух. массы кл. ст.) до минимального у Cassiope tetragona (2700 мкмоль/г сух. массы кл. ст.). Среди исследованных видов более высокие и близкие значения оказались у Betula nana, Dryas octopetala ; почти в 2–2,5 раза меньше у Salix рolaris и Cassiope tetragona. По данным Т. А. Горшковой [3], количество функциональных групп видоспецифично и зависит от происхождения вида, особенностей структуры клеточной стенки, физиологических особенностей, географического ареала. Есть данные, что количество ионогенных групп в структуре клеточной стенки органов может изменяться при действии тяжелых металлов [8; 12], засоления [15; 16].
Анализ процентных соотношений групп в клеточной стенке листа показал, что у всех видов наибольший процент приходится на карбоксильные группы гидроксикоричных кислот (от 28 % у Cassiope tetragona до 54 % у Salix рolaris) и фенольные ОН-группы (от 18 % у Salix рolaris до 35 % у Dryas octopetala), которые в основном входят в состав соединений фенольной природы. Наиболее низкие значения полу- чены по проценту аминогрупп у Dryas octopetala (7 %), а самые высокие – по процентному содержанию СООН ГКК (54 %) в клеточной стенке Salix polaris.
Одним из показателей функционального состояния клеточной стенки является константа ионизации ионообменных групп, значение которой зависит от качественного состава полимеров и степени их полимеризации.
Таблица 2
Константа ионизации ионообменных групп клеточной стенки листа растений Арктики
|
Вид растений |
Константа ионизации ионообменных групп, рКa |
|||
|
Аминогруппы |
СООН ПГУК* |
СООН ГКК** |
Фенольные ОН-груп-пы |
|
|
Betula nana |
4,50 ± 0,62 |
7,04 ± 0,22 |
7,95 ± 0,15 |
9,40 ± 0,08 |
|
Dryas octopetala |
3,38 ± 0,58 |
5,87 ± 0,35 |
7,79 ± 0,18 |
9,03 ± 0,09 |
|
Salix рolaris |
4,05 ± 0,45 |
6,42 ± 0,41 |
8,07 ± 0,10 |
9,73 ± 0,10 |
|
Cassiope tetragona |
3,39 ± 0,32 |
6,22 ± 0,12 |
7,62 ± 0,10 |
9,35 ± 0,11 |
Примечание. * – карбоксильные группы ά-D-поли-галактуроновой кислоты; ** – карбоксильные группы ги-дроксикоричных кислот.
Наиболее высокие значения константы ионизации (рКа) отмечены для фенольных-ОН групп и СООН ГКК, входящих в состав фенольных соединений, несколько ниже для СООН ПГУК – компонентов пектиновых веществ и еще ниже для аминогрупп, которые включаются в белки и ферменты (табл. 2). Однако различия между видами растений по диапазону варьирования рКа оказались более значимыми по СООН ГКК и фенольным ОН-группам. Это может означать, что в метаболической составляющей клеточной стенки видов, обитающих в высоких широтах, в большей степени участвуют соединения фенольной природы. Вклад более высокомолекулярных соединений, несущих аминогруппу (белки) в состав клеточной стенки, выше у Betula nana и Salix рolaris.
Для клеточной стенки как структурной единицы клетки характерно свойство набухания, которое определяет ее гидравлическую проводимость. Коэффициент Кнаб (г Н2О/г сухой массы клеточных стенок) является количественной характеристикой проницаемости полимерного матрикса клеточной стенки и зависит от степени сшивки полимеров (жесткости), общего числа функциональных групп, константы их ионизации и концентрации внешнего раствора [6].
Исследование показало, что самые высокие значения Кнаб в воде были получены на клеточной стенке Salix рolaris (2,6 г Н2О/г сухой массы клеточных стенок). Более низкие у Betula nana и Dryas octopetala (1,93 и 1,45 г Н2О/г сухой массы клеточных стенок соответственно), самые низкие (0,83 г Н2О/г сухой массы клеточных стенок) у Cassiope tetragona. Известно, что, чем выше степень сшивки полимерных цепей клеточной стенки, жестче и плотнее ее структура, тем меньше набухание. Этим свойствам отвечает клеточная стенка Cassiope tetragona, у которой высокая степень кутинизации и осмоления особенно покровных тканей, но также паренхим- ных клеток приводит к жесткости структуры. По представлениям Н. Р. Мейчик [6], у растений существует прямая связь между способностью к набуханию полимерного матрикса клеточной стенки и интенсивностью водного потока. Высокий коэффициент набухания у Salix polaris предполагает и наиболее слабую степень сшивки полимерных цепей клеточной стенки, и более интенсивный по сравнению с другими видами водный поток по системе клеточных стенок.
Из литературы известно, что процесс набухания клеточной стенки корней связан с величиной рН внешнего раствора [14], для листьев этих данных меньше.
Для растений Арктики максимальные значения коэффициента набухания клеточной стенки листа при изменении рН внешнего раствора значительно выше, чем для более южных районов.
Так, в диапазоне рН от 2–3 до 9–11 Кнаб клеточной стенки изменяется у листа Betula nana от 1,8 до 6,5; Salix polaris от 3,5 до 5; Dryas octopetala от 1,8 до 5,5; Cassiope tetragona от 2 до 3,7 г Н2О/г сухой массы клеточных стенок (см. рисунок).
Кнаб клеточной стенки листа ив г. Петрозаводска варьирует в более узком диапазоне: для Salix
alba от 1,7 до 2,8 г и Salix caprea от 1,7 до 3,5 г Н2О/г сухой массы клеточных стенок. Сходные данные были получены из этого же района для листьев Betula pendula : коэффициент набухания изменялся в диапазоне от 1,5 до 3,5 г Н2О/г сухой массы клеточных стенок (устные данные Н. А. Галибиной).
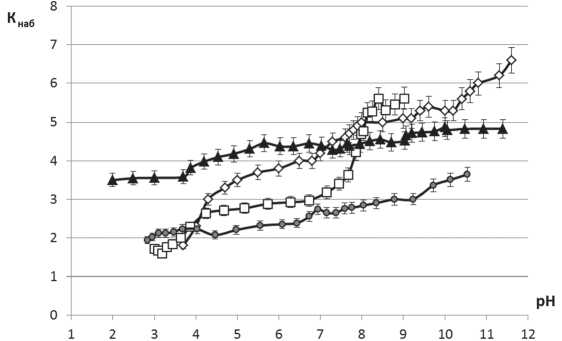
—O—Betula папа -О- Dryas octopetala —*—Salix polaris —•— Cassiope tetragona
Зависимость константы набухания (г Н2О/г сухой массы клет. ст.) клеточной стенки листа растений Арктики от рН раствора
Известно, что свойство набухания клеточной стенки лежит в основе механизма роста растения растяжением, который ингибируется в условиях Арктики [2]. Высокие значения Кнаб, полученные в настоящей работе, могут свидетельствовать о важности этого свойства клеточной стенки как потенциальной ростовой функции растений при изменении климата. Большим потенциалом роста, согласно диапазону варьирования Кнаб клеточной стенки, обладают такие виды, как Betula nana и Dryas octopetala , размеры листа могут увеличиться в 3–4 раза. Меньший ростовой потенциал можно предположить у Cassiope tetragona и значительно меньший у Salix polaris (1,5 раза). Однако на современном этапе роста и развития исследуемых видов свойство набухания клеточных стенок важно для увеличения метаболической активности, которая тесно связана с водным режимом растения в системе апопласта.
Проведенное исследование показало, что клеточная стенка листа растений Арктики является катионообменником, который представлен сходным составом ионообменных групп с клеточными стенками других изученных растений, в частности тундровой зоны. Однако особенностью растений Арктики является увеличение (в 2–3 раза) количества ионообменных групп каждого типа в структуре клеточной стенки листа по сравнению с видами других климатических зон. Кроме того, установлена ведущая роль в метаболизме компонентов клеточной стенки соединений фенольной природы, в меньшей степени – пектиновых и белковых веществ. Это хорошо соотносится с ранее полученными данными о повышенном (почти в 5 раз) содержании суммы флавоноидов у растений Арктики [5]. Сравнительный анализ исследованных видов позволяет предположить два пути адаптации на уровне такой структуры, как клеточная стенка. Это структурная адаптация (избегание), которая отмечается у Cassiope tetragona и состоит в синтезе защитных веществ (кутина, суберина), предотвращающих прямое воздействие факторов среды на растительный организм, и функциональная адаптация (увеличение количества ионообменных групп), которая свойственна клеточной стенке листа Betula nana . Если структурная адаптация свойственна широко распространенному виду на территории Западного Шпицбергена, то метаболическая – виду, спорадически представленному на этой территории, которая является крайней точкой его ареала.
Полученные данные впервые показали, что имеются функциональные различия на уровне состава ионогенных групп клеточной стенки у видов Арктики, но все виды характеризуются показателями ее высокой функциональной активности по сравнению с растениями более низких широт. Этот результат согласуется с более ранними работами о высокой фотосинтетиче- ской и дыхательной активности растений Арктики [17], [18].
В процессе эволюции одним из условий продвижения растений в Арктике было поддержание гомеостаза организма в экстремальных условиях среды. Гомеостаз внутренней среды достигался, в частности, и развитием покровных тканей и защитных структур, охраняющих организм от негативного воздействия факторов. Эволюция арктических растений сопровождалась развитием органов и механизмов, обеспечивающих селективную связь растений со средой, что могло идти через появление дополнительных функций у имеющихся структур.
В этот процесс могла быть включена и клеточная стенка, которая, помимо защитной функции, была включена и в метаболические процессы, что привело к повышению целостности организма [1]. Особенно актуально это оказалось для северных растений с их усилением роли апопластического потока, в котором большое значение отводится метаболическим процессам в свободном пространстве клеточной стенки [2]. Это совпадает и со стратегией приспособительной эволюции растений, которая идет по пути увеличения общей эффективности структур и минимизации негативного воздействия факторов внешней среды [1].
* Работа выполнена при финансовой поддержке Программы стратегического развития ПетрГУ, Международного гранта Программы ЕИСП ПГС Карелия «Развитие лесных плантаций на отвалах карьеров, их облесение и фитоочистка в России и Финляндии».
Список литературы Некоторые особенности транспортной функции клеточной стенки растений Арктики
- Волков И. В. Введение в экологию высокогорных растений: Учеб. пособие. Томск: Изд-во ТГПУ, 2006. 416 с.
- Гамалей Ю. В. Транспортная система сосудистых растений. Происхождение, структура, функции, развитие, анализ разнообразия типов по таксономическим и эколого-географическим группам растений, эволюция и экологическая специализация транспортной системы. СПб.: Изд-во С.-Петерб. ун-та, 2004. 424 с.
- Горшкова Т. А. Растительная клеточная стенка как динамическая система. М.: Наука, 2007. 429 с.
- Королева Н. Е., Константинова Н. А., Белкина О. А., Давыдов Д. А., Лихачев А. Ю., Савченко А. Н., Урбанавиче И. Н. Флора и растительность побережья залива Грен-фьорд (архипелаг Шпицберген). Апатиты, 2008. 35 с.
- Марковская Е. Ф., Шмакова Н. Ю. Содержание флавоноидов в листьях сосудистых растений Западного Шпицбергена//Растительные ресурсы. 2012. Вып. 4. С. 547-554.
- Мейчик Н. Р. Ионный обмен и диффузия в клеточных стенках растений: Автореф. дис.. д-ра биол. наук. М., 2007. 48 с.
- Славинская Г. В., Хохлов В. Ю. Потенциометрическое титрование ионитов: Учеб. пособие. Воронеж, 2004. 36 с.
- Теребова Е. Н., Галибина Н. А. Структурно-функциональное состояние хвои Pinus sylvestris (Pinaceae) в условиях загрязнения диоксидом серы и тяжелыми металлами//Растительные ресурсы. 2010. Вып. 2. С. 61-73.
- Шарова Е. И. Клеточная стенка растений. СПб.: Изд-во С.-Петерб. ун-та, 2004. 156 с.
- Galibina N. A., Terebova E. N. Characterization of cell wall properties in needles from scotch pine trees of various vigor//Russian Journal of Plant Physiology. 2008. Vol. 55. № 3. P. 419-425.
- Iiyama K., Lam T., Stone B. Covalent cross-links in the cell wall//Plant physiol. 1994. Vol. 104. P. 315-320.
- Krzeslowska M. The cell wall in plant cell response to trace metals: polysaccharide remodeling and its role in defense strategy//Acta Physiol Plant. 2011. Vol. 33. P. 35-51.
- Lamport D. T. A. Cell wall metabolism//Plant physiol. 1970. Vol. 21. P. 235-270.
- Meychik N. R., Yermakov I. P. Swelling of roots cell walls as an indicator of their functional state//Biochemistry (Moscow). 2001. Vol. 66. № 2. P. 178-187.
- Meychik N. R., Nikolaeva Yu. I., Yermakov I. P. Ion-exchange properties of cell walls of Spinacia oleracea L. roots under different environmental salt conditions//Biochemistry (Moscow). 2006. Vol. 71. № 7. P. 781-789.
- Meychik N. R., Yermakov I. P., Khonarmand S. D., Nikolaeva Y. I. Ion-Exchange properties of cell walls in Chickpea cultivars with different sensitivities to salinity//Russian Journal of Plant Physiology. 2010. Vol. 57. № 5. P. 620-630.
- Muraoka Н., No da H., Uch id a M. Photosynthetic characteristics and biomass distribution of the dominant vascular plant species in a high Arctic tundra ecosystem, Ny-Alesund, Svalbard: implications for their role in ecosystem carbon gain//J. Plant Res. 2008. Vol. 121. P. 137-145.
- Semichatova O. A., Ivanova T. I., Kirpichnikova O. V. Сomparative study of dark respiration in plants inhabiting arctic (Wrangel Island) and temperate climate zones//Russian Journal of Plant Physiology. 2007. Vol. 54. № 5. P. 582-588.
- Wallace G., Fry S. C. Phenolic components of the plant cell wall//Int. Rev. Cyt. 1994. Vol. 151. P. 229-267.
