Некоторые закономерности формирования подземных органов бореальных орхидей: размеры и микоризообразование

Автор: Бетехтина Анна Анатольевна, Гайсина Динара Фиргатовна, Веселкин Денис Васильевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 1, 2014 года.
Бесплатный доступ
При сопоставлении признаков строения подземных органов 14 видов бореальных орхидных показана отрицательная сопряженность между линейной протяженностью (длиной) отдельных корней и подземных органов в целом и 1) интенсивностью развития микоризы и 2) поперечными размерами подземных органов. Между развитием микоризы и поперечными размерами корней и размером центрального цилиндра наблюдается положительная зависимость. Установленные трейдоффы отражают важность варьирующего уровня специализации подземных органов орхидных к осуществлению симбиотических взаимодействий с грибами микоризных и микогетеротрофных ассоциаций для морфогенетических преобразований их подземных органов.
Орхидные, орхидеи, подземные органы, морфология, корни, микориза, функциональные свойства растений, морфогенез, структурные признаки
Короткий адрес: https://sciup.org/146116489
IDR: 146116489 | УДК: 582.594-143:581.557.24
Some conformities of boreal orchids underground organs formation: size and mycorrhiza formation
The negative correlation between the length of individual roots and that of entire underground system, from one side, and 1) the degree of mycorrhiza development, and 2) diameter of organs, from the other, has been shown on 14 species of boreal orchids. The positive correlation between the development of mycorrhiza and roots’ diameter was revealed. These tradeoffs reflect the importance of the varying level of orchids’ underground organs speciation to symbiosis with fungi from mycorrhizal and mycoheterotrophic associations.
Текст научной статьи Некоторые закономерности формирования подземных органов бореальных орхидей: размеры и микоризообразование
Введение. Уникальность многих биологических и экологических признаков представителей сем. Orchidaceae обуславливает значительный интерес к их изучению. В частности, накоплен значительный объем информации о биоморфологических, онтогенетических и экологических особенностях подземных органов орхидных, которые изучены, пожалуй, лучше, чем у большинства других дикорастущих растений. Одной из важных сторон биологии наземных Orchidaceae является облигатное для большинства видов на протяжении всего онтогенеза взаимодействие корней и других подземных органов с грибами, приводящее у разных видов к формированию эумицетных толипофаговых или эумицетных птиофаговых микориз (Селиванов, 1981). Первые характерны для фотоавтотрофных видов; вторые - для бесхлорофилльных, получающих от грибного симбионта не только воду и минеральные элементы, но и органические вещества. Для обозначения таких облигатно зависимых от
Работа выполнена при поддержке РФФИ (грант № 12-04-32055-а) и гранта
Президента РФ по поддержке ведущих научных школ (НШ-5325.2012.4)
Вестник ТвГУ. Серия "Биология и экология". 2014. № 1 грибов растений предложены термины «холомикотрофные» и «микогетеротрофные» (Bidartondo, 2005). Предполагается, что элементы микогетеротрофии свойственны многим зеленым орхидеям (Gebauer, Meyer, 2003). Микогетеротрофные ассоциации формируются также у бесхлорофилльных проростков орхидей - протокормов.
К наиболее существенным положениям, сформулированным в отношении связи жизненной формы орхидных умеренной зоны со строением, развитием и микоризацией их подземных органов, можно отнести три, изложенные Татаренко (1993): (1) существует негативная сопряженность между уровнем вегетативного развития особей разных видов и интенсивностью микоризообразования у них; (2) для автотрофных корневищных видов орхидных свойственна в среднем более низкая микоризация по сравнению с тубероидными; при этом корни у корневищных видов относительно длинные, тонкие и долгоживущие, а у тубероидных — относительно короткие, толстые и маложивущие; (3) в онтогенезе от ювенильных к генеративным особям интенсивность микоризации у многих тубероидных видов снижается, а у корневищных - возрастает.
Хорошая изученность особенностей строения подземных органов и значительная, по сравнению со многими другими группами растений, детерминированность онтогенеза делают орхидные удобным объектом для проверки ряда предположений относительно общих закономерностей организации подземных органов растений. В частности, в настоящей работе мы задались целью рассмотреть, некоторые взаимосвязи между анатомо-морфологическими особенностями корней и развитием микоризы у орхидных умеренной зоны.
Методика. Оригинальные данные о строении подземных органов получены в отношении 14 видов орхидей (таблица), представляющих биоморфологические группы видов: 1) корневищных (Cephalanthera rubra (L.) Rich.; Cypripedium guttatum Sw.; Epipactis atrorubens (Hoffm. ex Bernh.) Besser; Listera ovata (L.) R. Br.); Goodyera repens (L.) R. Br.); 2) co стеблекорневыми тубероидами (Dactylorhiza fuchsii (Druce) Soo; D. hebridensis (Wilmott) Aver.; D. incarnata (L.) Soo; D. raunsteineri (Saul.) Soo; Gymnadenia conopsea (L.) R. Br.; Platanthera bifolia (L.) Rich.); 3) бесхлорофилльных (микогетеротрофных) -короткокорневищного с запасающими корнями (Neottia nidus-avis (L.) Rich.) и коралловидно-корневищных (Corallorrhiza trifida Chatel.; Epipogium aphyllum Sw.). Растения были собраны в июне-августе 2010 г. на территории г. Екатеринбурга (Уктусский лесопарк; 5б°4б' с.ш.; 60°39' в.д.), Сысертского р-на Свердловской области (Биостанция УрФУ; 56°36' с.ш.; 61°03' в.д.) и Советского р-на Ханты-Мансийского автономного округа (государственный природный заповедник «Малая Сосьва» 60°.. .°62 с.ш.; 61°.. .66° в.д.).
Анализ морфо-анатомического строения подземных органов (корней у видов с корневищами и со стеблекорневыми тубероидам; микоризомов - у микогетеротрофных коралловидно-корневищных видов) и оценку развития микоризы в них осуществляли на поперечных срезах, приготовленных на замораживающем микротоме. Фиксировали: общий диаметр корня и размер центрального цилиндра. Определяли интенсивность развития микоризы (С, %, по: Селиванов, 1981).
Проанализировано от 2-3 (микогетеротрофные виды) до 12-15 (Platanthera bifolia, Cypripedium guttatum) особей каждого вида. У каждой особи анализировали строение 2-5 отдельных корней. Все измерения в дальнейшем были усреднены (сначала - в пределах особи; затем - между особями) для получения средней видовой характеристики признаков.
Данные о морфологических характеристиках подземных органов (длине корней и размерах микоризомов) аккумулированы из литературы (Вахромеева и др., 1993а, б, 1996, 2003; Варлыгина, 1995; Татаренко, 1995, 1996; Вахромеева, 2000; Татаренко, Кондо, 2003; Ефимов, 2004; Мамаев и др., 2004; Татаренко и др., 2005, 2006; Татаренко, Варывдина, 2006; Быченко, 2007; Железная, 2007; Vakhrameeva et al., 2008). Нами тажке использованы литературные оценки интенсивности микоризообразования у орхидных (Татаренко, 1996; Вахромеева, 2000; Вахромеева и др., 2003; Татаренко, Кондо, 2003; Татаренко и др., 2005, 2006; Татаренко, Варывдина, 2006). Учитывались сведения о строении только генеративных особей. Поскольку в большинстве случаев в источниках в качестве видовых характеристик были приведены некоторые границы варьирования признаков, в расчетах мы использовали среднюю арифметическую между минимальными и максимальными значениями.
Результаты и обсуждение
Длина корней и микоризообразование. Предположение о негативной сопряженности между длиной корней у орхидных и обилием микоризных грибов в них требует проверки. Это предположение можно сделать на основании того, что у автотрофных корневищных орхидей корни обычно длинные, но слабо заселены микоризными грибами, в противоположность тубероидным видам, корни которых короче и содержат больше грибов (Татаренко, 1996). Будет ли справедливым данное утверждение, если переформулировать его из терминов сравнительной биоморфологии в терминах анализа функциональных свойств растений?
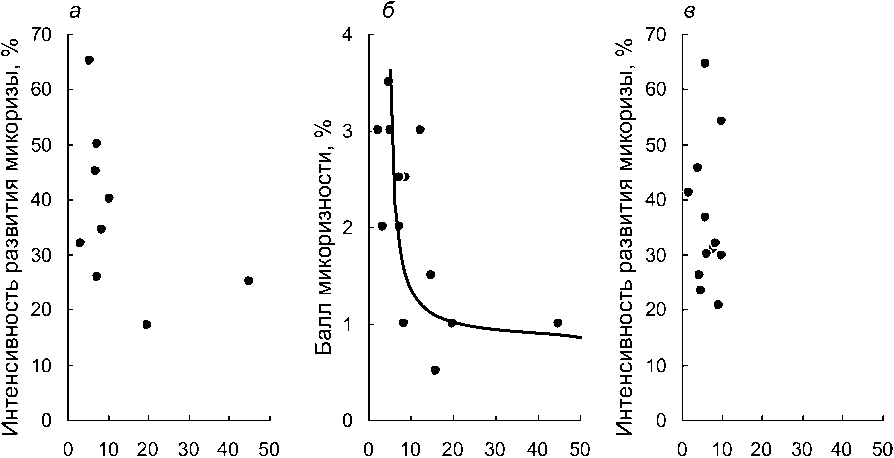
Средняя длина одного корня, см
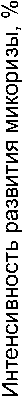
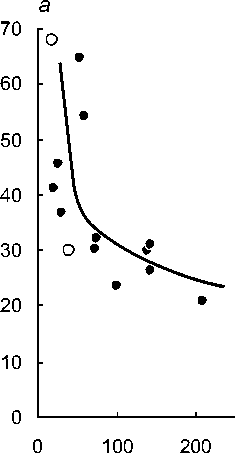
Рис. 1. Изменение интенсивности развития микоризы при увеличении длины среднего корня у разных видов орхидных по несколькими массивам данных:
а, б - литературные данные; в - длина корня - литературные данные, интенсивность развития микоризы - оригинальные оценки
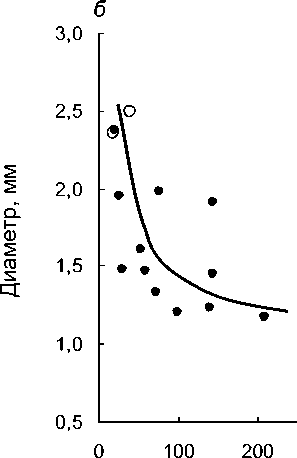
Суммарная длина подземных органов, см
Рис. 2 . Снижение интенсивности развития микоризы (а) и поперечных размеров подземных органов (б) с увеличением их линейной протяженности у разных видов орхидных:
длина подземных органов - литературные оценки, интенсивность развития микоризы и диаметр - оригинальные. Залитые точки - виды с корнями;
незалитые - с микоризомами.
Сопоставление средней длины отдельных корней со средними уровнями интенсивности микоризообразования, выраженным в исходных величинах (т.е. в %), оказалось возможным сделать по литературным данным только для 8 видов (рис. 1а, п=9, так как для одного вида есть две независимые оценки). При общем просматривающемся снижении интенсивности развития микоризы с ростом длины корней, оценить эту связь, как значимую, невозможно (^(коэффициент корреляции Спирмена)=-0,58; Р=О,1ОЗ). Отрицательную зависимость между длиной отдельного корня и развитием микоризы удалось подтвердить при использовании в качестве показателя ее развития среднего балла микоризности вида (по: Татаренко, 1996) (рис. 16; «=13; rs=0,68; P=0,009). При совмещении оригинальных оценок развития микоризы с литературными данными о длине отдельных корней (рис. 1в) не удается установить какой-либо взаимосвязи между этими характеристиками (и=12; г§=—0,16; 7э=0,624). Это происходит, прежде всего, потому, что проанализированные нами виды демонстрируют низкий размах средней длины корня. Посему, на уровне отдельного корня справедливость утверждения о негативной связи между длиной корней и интенсивностью развития микоризы надежно подтвердить не удается. Другими словами, длина среднего корня какого-либо вида вряд ли может быть значительным детерминирующим фактором успешности развития симбиотических грибов.
Но с ростом средней для вида суммарной протяженности подземных органов, средняя микоризность снижается (рис. 2а). Эта закономерность устанавливается как при анализе 12 видов с настоящими корнями (rs=-0,80; P=0,002), так и 14 видов, включая бескорневые микогетеротрофные Corallorrhiza trifida и Epipogium aphyllum (rs=-0,77; P=0,001). С увеличением средней линейной протяженности подземных органов у изученных 14 видов бореальных орхидей ожидаемо снижается средний поперечный размер корней (рис. 26), что статистически подтверждается как при отдельном анализе 12 видов с настоящими корнями (rs=-0,62; Р=0,030), так и для всего комплекса из 14 видов, включающего микоризомные орхидеи (fs=-0,69; Р=0,006).
Таким образом, утверждение, что у видов орхидных с протяженной корневой системой обычно наблюдаются относительно тонкие корни с низкой встречаемостью в них грибных структур в целом справедливо. Толстые же подземные органы, как собственно корни, так и корневища, активно заселяемые микоризными грибами, ассоциируются обычно с их незначительными линейными размерами. Из рассмотренных орхидей наиболее ярким представителем первой стратегии, т.е. видом с относительно тонкими, протяженными и слабо микоризованными корнями, является Cypripedium guttatum. Выражено
Вестник ТвГУ. Серия "Биология и экология". 2014. № 1 противоположную организацию подземных органов, для которой характерны компактные, толстые и сильно микоризованные корни или микоризомы, демонстрируют два бескорневых микогетеротрофных вида и Neottia nidus-avis, Platanthera bifolia и Gymnadenia conopsea.
Общий диаметр, мм
0,2 0,4 0,6
Диаметр центрального цилиндра, мм
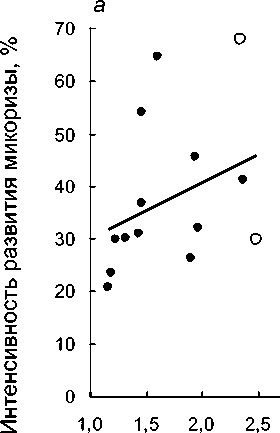
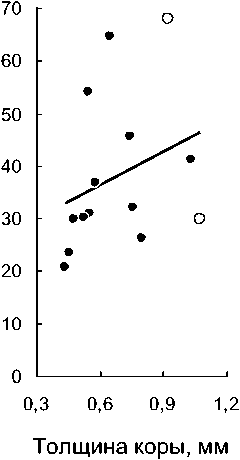
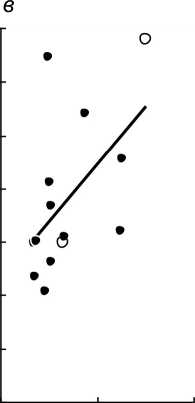
Рис. 3 . Увеличение интенсивности развития микоризы при увеличении общего поперечного размера подземных органов (а), толщины коры (б) и диаметра центрального цилиндра (в) у разных видов орхидных: залитые точки - виды с корнями; незалитые - с микоризомами
Толщина подземных органов и микоризообразование. Однонаправленная негативная сопряженность поперечных размеров и микоризности подземных органов орхидных с их линейной протяженностью заставляет предполагать возможность положительной связи между первыми двумя признаками. При этом гипотеза о положительной корреляции между микоризностью и поперечным размером корней в целом справедлива для широкой группы растений (John, 1980; Eissenstat, 1992). В связи с этим, интересно протестировать гипотезы относительно такой биоморфологически специфичной группы, как бореальные Orchidaceae. На рассматриваемых материалах данная гипотеза получает слабое подтверждение. Значимая положительная корреляция между средним диаметром корней и средней интенсивностью развития микоризы наблюдается только при рассмотрении 12 видов с настоящими корнями (rs=+0,63; Р=0,028). При одновременном рассмотрении корневых и бескорневых видов соответствующий коэффициент корреляции незначим (рис. За; rs=+0,53; Е*=0,051). Интересно, что развитие микоризы теснее коррелирует с размером центрального цилиндра (рис. Зе; и=14; rs=+0,60; Р=0,023), а не с толщиной коры (рис. 36; и=14; rs=+0,45; Р=0,105).
Таблица
Признаки строения подземных органов изученных видов
|
Вид |
Строение среднего корня |
Интенсивность развития микоризы, % |
Строение корневой системы у средней особи |
||||
|
длина, см |
диаметр, мм |
парциальный объем коры, % |
число корней на годичном приросте, шт. |
продолжительность жизни корней, лет [3] |
длина всех корней, см |
||
|
Корневищные виды |
|||||||
|
Cephalanthera rubra |
8,0 [7] |
1,45 |
95 |
31 |
4,5 [7] |
4,0 |
144 |
|
Cypripedium guttatum |
9,5 [3, 9] |
1,17 |
94 |
21 |
5,5 [3] |
4,0 |
209 |
|
Epipact is atrorubens |
4,5 [4, 6] |
1,91 |
95 |
26 |
8,0 [6] |
4,0 |
144 |
|
Goodyera repens |
5,0 [1] |
1,20 |
95 |
23 |
5,0 [1] |
4,0 |
100 |
|
Listera ovata |
10,0 [7] |
1,23 |
95 |
30 |
3,5 [7] |
4,0 |
140 |
|
Виды co стеблекорневыми тубероидами |
|||||||
|
Dactylorhiza fuchsii |
6,0 [5, 8] |
1,61 |
96 |
65 |
6,0 [5, 7] |
1,5 |
54 |
|
Dactylorhiza hebridensis |
6,5 [3, И] |
1,32 |
96 |
30 |
7,5 [3, И] |
1,5 |
73 |
|
Dactylorhiza incarnata |
8,5 [5, 10] |
1,98 |
95 |
32 |
6,0 [5, 10] |
1,5 |
77 |
|
Dactylorhiza traunsteineri |
10,0 [7] |
1,47 |
93 |
54 |
4,0 [7] |
1,5 |
60 |
|
Gymnadenia conopsea |
4,0 [2, 3] |
1,95 |
95 |
46 |
4,5 [2, 3] |
1,5 |
27 |
|
Platanthera bifolia |
6,0 [3, 7] |
1,47 |
96 |
37 |
3,5 [3] |
1,5 |
32 |
|
Бесхлорофилльный (микогете |
ротрофный) короткокорневищный с запас ющими корнями |
||||||
|
Neottia nidus-avis |
2,0 [3, 7] |
2,37 |
98 |
41 |
3,5 [3, 7] |
3,0 |
21 |
|
Бесхлорофилльные (микогетеротрофные) коралловидно-корневищные виды (приведены признаки строения корневищ - микоризомов) |
|||||||
|
Corallorrhiza trifida |
- |
2,35 |
95 |
68 |
- |
- |
20 [7] |
|
Epipogium aphyllum |
- |
2,49 |
98 |
30 |
- |
- |
40 [3, 7] |
Примечание. Прочерк обозначает отсутствие признака.
Закономерности формирования подземных органов. Негативную сопряженность между линейными размерами подземных органов бореальных Orchidaceae и их диаметром или между линейными размерами и заселенностью микоризными грибами можно трактовать как случаи конкурентных отрицательных согласованностей, возникающих при формировании разных систем или признаков организмов вследствие ограниченности имеющихся в их распоряжении ресурсов. Такие зависимости описываются термином «трейдофф» (Гиляров, 2007) и широко исследуются в экофизиологии растений. Обсуждаемые трейдоффы ожидаемы и основаны на накопленных знаниях об особенностях организации корней и корневищ орхидных разных биоморфологических групп. Они также соответствуют общим представлениям об организации вегетативных органов растений. Ясно, например, что у особи не может быть очень много толстых корней из-за очевидных значительных затрат ресурсов на их формирование. Логично также ожидать, что малая длина корней, не позволяющая эксплуатировать более или менее значительные объемы почвы, должна компенсироваться какими-либо дополнительными приспособлениями для обеспечения должного уровня снабжения ресурсами. У орхидных такие приспособления реализуются в виде симбиотических взаимодействий с микоризными грибами. В связи с этим формирование толстых корней является функционально нагруженной реакцией, поскольку при малой протяженности корней основной их функцией становится поддержание размещающихся в коре микобионтов и обеспечение процессов обмена (Brundert, 1991; Barker et al., 1998).
Стратегия редукции линейных размеров подземных органов при максимизации их толщины получает крайнее выражение у микогетеротрофных орхидей, трофически полностью зависимых от микобионтов. У части микогетеротрофных видов вместилищем грибов становятся структуры стеблевого происхождения - микоризомы, а корни полностью отсутствуют. Такой переход функций можно связывать, например, с потенциально большей долговечностью и автономностью стеблевых органов по сравнению с корнями, продолжительность функционирования которых обычно более детерминирована. Заметим, что по отношению к таким видам использование термина микориза (грибокорень) для описания симбиотических структур и самого феномена взаимодействия оправдано лишь соображениями преемственности знания. А отсюда желательным видится и коррекция использования термина «микоризом».
Если отталкиваться от признания основными функциями подземных органов орхидных функций обеспечения симбиотических взаимодействий, интересно обсудить причины наблюдаемой более тесной связи уровня их микоризности с размером центрального цилиндра, а не с размером коры. С одной стороны, размер центрального цилиндра традиционно связывают с активностью транспортной функции корня (Вахмистров, 1985; Wahl, Ryser, 2000), а размер коры - с синтетической биохимической активностью (Вахмистров, 1985) или, как в нашем случае, с осуществлением симбиотических взаимодействий (Brundert, 1991; Barker et al., 1998). Поэтому можно было бы предположить связь между развитием микоризы и размером коры. Однако, необходимо принимать во внимание специфику регуляции абсолютных (например, абсолютный объем микобионта в корнях одной особи) и относительных (интенсивность развития микоризы - доля клеток коры корня с грибными структурами) показателей развития микоризы. Возможно, что интенсивность развития микоризы - это не универсальная характеристика успешности формирования симбиоза.
Максимизация доли растительных клеток, несущих грибные структуры, не обязательно является конечной «целью» взаимодействия орхидей и микоризных грибов. Увеличение же абсолютного обилия гриба может достигаться при константности относительных величин, если предполагать увеличение абсолютных размеров вмещающей микобионта коры корня. С этих позиций наблюдаемую относительно более тесную, хотя и далекую от функциональной, связь между обилием микобиона в коре и размером центрального цилиндра можно попытаться трактовать как отражение согласованности между притоком веществ в подземные органы (вероятно пропорциональным обилию гриба в коре) и их потоком из подземных органов в надземные (пропорциональным размеру центрального цилиндра).
В нашем распоряжении, к сожалению, оказалось недостаточно данных для того, чтобы быть полностью уверенными в абсолютной однозначности представленных результатов и заключений. Это обуславливает гипотетичность части представленных рассуждений, имеющих вследствие этого программный характер и требующих верификации на расширенном материале. Аргументом против абсолютизации конкретных представленных оценок и конкретных форм зависимостей является условность части фактических материалов, связанная с необходимостью использования разрозненных литературных оценок без учета таких факторов как индивидуальная, экологическая и возможная внутривидовая изменчивость строения подземных органов орхидных. Очевидным является также обоснованность замены в межвидовых сравнениях таких абсолютных размерных характеристик подземных органов как суммарная длина корней какими-нибудь индексами, отражающими или пропорциональность развития подземных и надземных органов или пропорциональность развития разных частей подземных органов.
Заключение. Основные морфологические особенности строения подземных органов орхидных бореальной зоны могут быть удовлетворительно описаны как результат реализации системы корреляций между проявлением частных признаков: отрицательных (между линейной протяженностью органов и их поперечными размерами; между линейными размерами органов и уровнем развития микоризы) и положительных (между поперечными размерами органов, особенно центрального цилиндра в их составе, и развитием микоризы). Такая согласованность указывает на то, что важным вектором морфологической диверсификации вегетативных органов бореальных орхидных является разная степень специализации и взаимодействия с микоризными грибами и грибами, участвующими в формировании микогетеротрофных ассоциаций. Орхидные оказываются удобнойя и перспективной модельной группой для верификации некоторых представлений о частных морфогенетических процессах, лежащих в основе формирования структуры подземных органов растений. Абсолютно необходимой представляетсяч строгая проверка намеченных и иных закономерностей формирования подземных органов орхидных с учетом их онтогенетической, сезонной, индивидуальной, экологической, межвидовой и биоморфологической специфики.
Список литературы Некоторые закономерности формирования подземных органов бореальных орхидей: размеры и микоризообразование
- Быченко Т.М. 2007. Онтогенез башмачка капельного (Cypripedium guttatum Sw.)//Онтогенетический атлас растений. Т. V. Йошкар-Ола: МарГУ. С. 211-219.
- Варлыгина Т.И. 1995. Род Тайник//Биол. флора Моск. обл. М.: Изд-во МГУ, Аргус. Вып. 11. С. 52-63.
- Вахмистров Д.Б. 1985. Ионный режим растений: эволюция проблемы//Новые направления в физиологии растений. М.: Наука. С. 214-230.
- Вахрамеева М.Г. 2000. Род Пальчатокоренник//Биол. флора Моск. обл. М.: Гриф и К. Вып. 14. С. 55-86.
- Вахрамеева М.Г., Блинова И.В., Богомолова Т.И., Жирнова Т.В. 2003. Пололепестник зеленый//Биол. флора Моск. обл. М.: Гриф и К. Вып. 15. С. 62-77.
- Вахрамеева М.Г., Быченко Т.М., Татаренко И.В., Экзерцева М.В. 1993. Мякотница однолистная//Биол. флора Моск. обл. М.: Изд-во МГУ. Вып. 9. Ч. 1. С. 40-50.
- Вахрамеева М.Г., Варлыгина Т.И., Баталов А.Е., Тимченко И.А., Богомолова Т.И. 1997. Род Дремлик//Биол. флора Моск. обл. М.: Полиэкс. Вып. 13. С. 50-87.
- Вахрамеева М.Г., Варлыгина Т.И., Куликов П.В. 1996. Пыльцеголовник длиннолистный//Биол. флора Моск. обл. М.: Изд-во МГУ, Аргус. Вып. 12. С. 48-59.
- Вахрамеева М.Г., Виноградова И.О., Татаренко И.В., Цепляева О.В. 1993. Кокушник комарниковый//Биол. флора Моск. обл. М.: Изд-во МГУ. Вып. 9. Ч. 1. С. 51-64.
- Вахрамеева М.Г., Денисова Л.В. 1975. Гудайера ползучая//Биол. флора Моск. обл. М.: Изд-во МГУ. Вып. 2. С. 5-10.
- Гиляров А.М. 2007. Ариаднина нить эволюционизма//Вестник РАН. Т. 77. № 6. С. 508-516.
- Ефимов П.Г. 2004. Род Epipactis Zinn (Orchidaceae) на территории России//Turczaninowia. Т. 7. № 3. С. 8-42.
- Железная Е.А. 2007. Онтогенез пальчатокоренника мясо-красного (Dactylorhiza incarnata (L.) Soo)//Онтогенетический атлас растений. Т. V. Йошкар-Ола: МарГУ. С. 286-291.
- Мамаев С.А., Князев М.С., Куликов П.В., Филиппов Е.Г. 2004. Орхидные Урала: систематика, биология, охрана. Екатеринбург: УрО РАН. 124 с.
- Селиванов И.А. 1981. Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза. М.: Наука. 232 с.
- Татаренко И.В. 1995. Микориза орхидных (Orchidaceae) Приморского края//Ботан. журн. Т. 80. № 8. С. 64-73.
- Татаренко И.В. 1996. Орхидные России: жизненные формы, биология, вопросы охраны. М.: Аргус. 206 c.
- Татаренко И.В., Варлыгина Т.И., Кондо К. 2005. Биоморфологические особенности Cremastra variabilis (Orchidaceae)//Ботан. журн. Т. 90. № 3. C. 378-386.
- Татаренко И.В., Варывдина И.В. 2006. Экспериментальное изучение микоризы и морфогенеза побегов Dactylorhiza fuchsii (Orchidaceae) в зависимости от интенсивности фотосинтеза//Бюлл. МОИП. Отд. Биол. Т. 111. Вып. 4. С. 46-51.
- Татаренко И.В., Кондо К. 2003. Биоморфологические особенности Dactylostalyx ringens (Orchidaceae)//Ботан. журн. Т. 88. № 4. С. 62-67.
- Татаренко И.В., Кучер Е.Н., Попкова Л.Л., Куликов П.В. 2006. Биоморфологические особенности Dactylorhiza iberica (Orchidaceae)//Ботан. журн. Т. 91. № 7. С. 1041-1048.
- Barker S.J., Tagu D., Delp G. 1998. Regulation of root and fungal morphogenesis in mycorrhizal symbioses//Plant physiology. V. 116. № 4. P. 1201-1207.
- Bidartondo M.I. 2005. The evolutionary ecology of mycoheterotrophy//New Phytol. V. 167. № 2. P. 335-352.
- Brundert М. 1991. Mycorrhizas in natural ecosystems//Advances Ecol. Res. 1991. V. 21. P. 171-313.
- Eissenstat D.M. 1992. Costs and benefits of constructing roots of small diameter//J. Plant Nutr. V. 15. № 6-7. P. 763-782.
- Gebauer G., Meyer M. 2003. 15N and 13C natural abundance of autotrophic and myco-heterotrophic orchids provides insight into nitrogen and carbon gain from fungal association//New Phytol. V. 160. № 1. P. 209-223.
- John St T.V. 1980. Root size, root hairs and mycorrhizal infection: a re-examination of Baylis's hypothesis with tropical trees//New Phytol. V. 84. № 3. P. 483-487.
- Vakhrameeva M.G., Tatarenko I.V., Varlygina T.I., Torosyan G.K., Zagulskii M.N. 2008. Orchids of Russia and adjacent countries (within the borders of the former USSR). Ruggell (Liechtenstein): A.R.G. Gantner Verlag. 690 p.
- Wahl S., Ryser P. 2000. Root tissue structure is linked to ecological strategies of grasses//New Phytol. V. 148. P. 459-471.
