Некоторые закономерности полиморфической изменчивости одонтологических признаков европейской рыжей полевки (Clethrionomys glareolus Schreber), обитающей в верховьях Волги и на сопредельных территориях
, обитающей в верховьях Волги и на сопредельных территориях")
Автор: Емельянова Алла Александровна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Зоология
Статья в выпуске: 7, 2008 года.
Бесплатный доступ
Рассматривается популяционная изменчивость рисунка жевательной поверхности моляров рыжей полевки, обитающей на территории Тверской обл. Обнаружены некоторые отличия строения моляров рыжих полевок, отловленных в хвойных лесах от структурных особенностей коренных зубов полевок из смешанных лесов, клинальная изменчивость формы внутренней поверхности передней непарной петли параконида и зависимость встречаемости ряда фенов от степени снашивания моляров.
Моляры, рыжая полевка, тверская область, фены
Короткий адрес: https://sciup.org/146116144
IDR: 146116144 | УДК: 591.15:591.431.4:599.323.43
Текст научной статьи Некоторые закономерности полиморфической изменчивости одонтологических признаков европейской рыжей полевки (Clethrionomys glareolus Schreber), обитающей в верховьях Волги и на сопредельных территориях
В современных популяционно-морфологических исследованиях широко распространен фенетический подход. В числе важнейших общебиологических задач, решаемых с помощью данного подхода, установление уровня сходства и иерархии популяционных группировок, оценка разнообразия особей и популяций, изучение вопросов адаптации, механизмов микроэволюции и действия естественного отбора. При изучении внутривидовой и видовой изменчивости большое значение придается морфологическим параметрам, для которых возможно выявление адаптивной сущности [15]. Среди подобных признаков особое место занимает изменчивость рисунка жевательной поверхности коренных зубов грызунов, так как установлено, что форма жевательной поверхности их моляров, с одной стороны, находится под генным контролем [19; 20; 21; 22], а с другой – является признаком пищевой специализации [5].
Изучение изменчивости полиморфических признаков зубной системы рыжей полевки, обитающей в верховьях Волги и на сопредельных территориях, административно входящих в состав Тверской обл., особенно интересно. На территории Тверской обл. исторически сложился переходный ландшафт от таежных к широколиственным лесам [11; 13; 16; 17]. Здесь живут, с одной стороны, представители фауны таежной провинции, с другой – представители фауны провинции широколиственных и смешанных лесов. Многие виды млекопитающих занимают окраинные участки ареалов и относятся к периферическим популяциям. Для некоторых видов в области проходит часть контактной зоны двух подвидов (европейская норка, заяц-беляк, летяга, темная полевка, полевка-экономка) и трех подвидовых форм (белка) [3; 4]. Здесь также соприкасаются границы распространения двух подвидов рыжей полевки: Clethrionomys glareolus suecicus Miller (1900) (темная или северная рыжая полевка) и Clethrionomys glareolus glareolus Schreber (1780) (обыкновенная рыжая полевка). Граница между ареалами подвидов пересекает Тверскую обл., проходя по линиям через Холм, Белый, далее Клин, Иваново. Из этого следует, что в западных районах области обитает номинальный подвид, в то время как остальные занимает северный [1; 14].
В качестве материала нами использовалось 450 рыжих полевок, отловленных в июне–июле 1999 г. в пяти административных районах Тверской обл.: Торопецком, Нелидовском, Зубцовском, Калининском и Лесном. Районы исследования в порядке их перечисления располагаются вдоль линии наибольшей протяженности области в направлении с юго-запада на северо-восток. В данном направлении наблюдаются изме- нения зимних температур, а также переход от зоны хвойно-широколиственных лесов к таежным. Значительная пространственная разобщенность географических точек, в которых проходил сбор серийного материала (от 60 до 120 км), наличие между ними обширных обезлесенных участков, болот и развитой речной сети, выполняющих барьерно-изолирующую роль, позволяли предполагать, что отловленные особи относятся к разным популяциям. При выборе точек отлова также учитывалась возможность влияния биотопических особенностей на специфику кормовой базы разных популяций и, следовательно, на строение элементов зубной системы. Так, места обитания рыжих полевок из Торопецкого, Нелидовского и Лесного районов приурочены к коренным хвойным лесам, а зверьки из Зубцовского и Калининского р-нов были отловлены в смешанных растительных ассоциациях.
При изучении строения моляров рыжей полевки за основу были взяты признаки и их вариации, предложенные Н.И. Лариной и И.В. Ереминой, а также морфотипы (фены) по И.В. Ереминой [6; 12]. Выделение фенов проводилось по рисункам, выполненным рисовальным аппаратом РА-6. Всего было выделено 296 вариаций 109 признаков элементов жевательной поверхности моляров. При сопоставлении фенофондов использовалось 113 вариаций 39 признаков, не подверженных возрастной изменчивости [7]; для оценки уровня сходства популяций применялись показатель Животовского (r) и критерий идентичности ( J ) [8; 9]. Констатация достоверности различий между выборками происходила при р≤0,05.
Высокий уровень различий фенофондов (р≤0,001), полученный при сопоставлении серий рыжих полевок по суммарному показателю сходства для признаков всех шести коренных зубов, подтвердил принадлежность выборок к разным популяциям. Популяции из Торопецкого, Нелидовского, Зубцовского и Калининского р-нов объединялись в единый комплекс на основании сходства по совокупности признаков. При этом обнаружено бóльшее сходство рыжих полевок из Торопецкого р-на со зверьками из ближайшего района – Нелидовского (r ср.=0,983), нежели с полевками из Зубцов-ского и Калининского р-нов (r ср.=0,975). Выборка из Нелидовского р-на, в свою очередь, более сходна с выборкой из Зубцовского р-на (r ср.=0,977). Минимальное значение показателя Животовского в этой группе популяций получено при сравнении серии из Калининского р-на с популяциями из Зубцовского и Нелидовского р-нов (r ср.=0,968). Популяция из Лесного р-на выделялась своеобразным фенофондом, обнаруживая несколько большее сходство с сериями рыжих полевок, места обитания которых приурочены к сходным биотопам – хвойным лесам: при сопоставлении данной популяции с выборками из Торопецкого и Нелидовского р-ов были получены r ср. – 0,963 и 0,966, а при сравнении с популяциями из Калининского и Зубцовского р-нов – 0,960 и 0,952 соответственно.
Известно, что использование суммарной характеристики какого-либо признака или группы признаков связано с феноменом потери части информации, а сравнение серий по частотам фенов может дать более детальную характеристику [18]. Исходя из этого при исследовании особенностей строения второго и третьего верхнечелюстных и первого нижнечелюстного моляров (М2, М3 и М1), на которые падает основная нагрузка при обработке пищи, кроме обобщенных суммарных показателей, применялся анализ распределения частот отдельных вариаций признаков строения зубной системы. Особое внимание обращалось на встречаемость фенов, отражающих удлинение режущего края моляров, поскольку таковое считается приспособлением к потреблению грубых кормов [2; 5] (рис. 1–3).
В ходе анализа материала были обнаружены некоторые отличия строения моляров рыжих полевок, обитающих в хвойных лесах (серии из Торопецкого, Нелидовского и Лесного р-нов) от структурных особенностей коренных зубов полевок из смешанных лесов (серии из Зубцовского и Калининского р-нов).
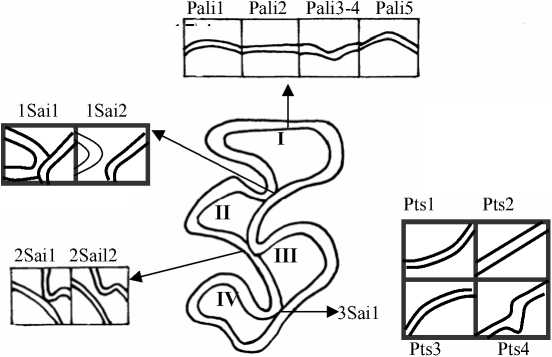
Рис. 1. Жевательная поверхность М2 и некоторые вариации её элементов .
Правая сторона тела: I – передняя непарная петля (протокон); II – IV основные треугольники (гипокон, паракон, метакон)
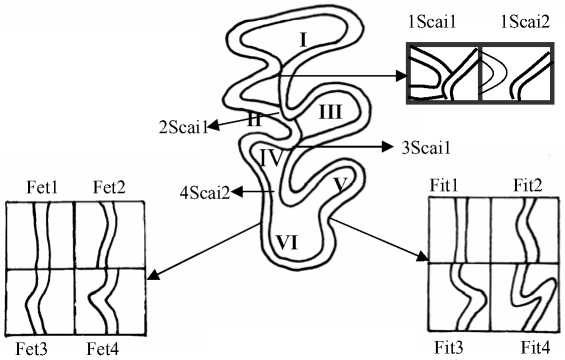
Рис. 2. Жевательная поверхность М3
и некоторые
вариации её элементов:
I – передняя непарная петля; II – IV – основные треугольники (протокон, гипокон, паракон, ме-такон); VI – талонус, или задняя непарная петля
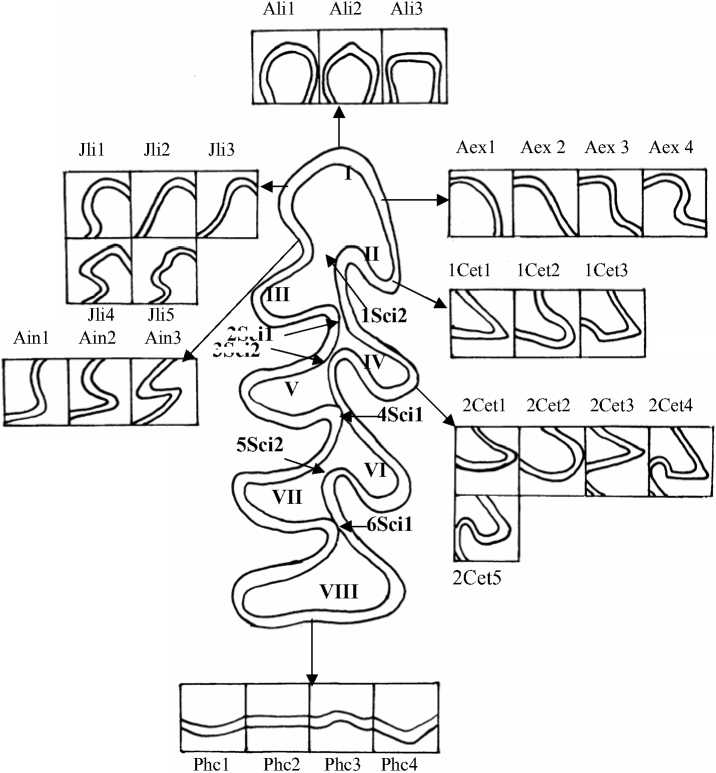
2Sci
3Sci2
Рис. 3. Жевательная поверхность М1 и некоторые вариации её элементов:
I – II – параконидный отдел (I – непарная петля, II – наружный треугольник в основании непарной петли); III – IV – треугольники в основании параконида; V – VII – основные треугольники (метаконид, протоконид, энтоконид); VIII – гипоконид (задняя непарная петля)
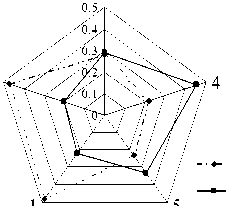
Так, угловатая форма передней непарной петли М2 (фен Pali5) ∗ более свойственна зверькам из Торопецкого, Нелидовского и Лесного р-нов – встречаемость 0,32, 0,27 и 0,30 соответственно. У рыжих полевок из Зубцовского и Калининского р-нов данный фен обнаружен с частотой 0,12 и 0,20 (рис.4, а). Уровня достоверности достигло отличие популяции из Зубцовского р-на от популяций из Торопецкого и Нелидовского р-нов (р≤0,05).
∗ Выделенные нами вариации и их обозначения не всегда соответствуют фенам, приведенным в «Каталоге основных вариаций краниологических признаков у грызунов» [12] .
Pali5
Pts4
Fet1
Fet3+Fet4
а
б
Рис. 4. Встречаемость некоторых фенов зубной системы в популяциях рыжей полевки Тверской обл.:
а – доля фенов, отражающих удлинение режущего края М 2 ; б – доля фенов, отражающих популяционные особенности формы наружной поверхности талонуса М 3 . Обозначения районов: 1 – Калининский; 2 – Зуб-цовский; 3 – Торопецкий; 4 – Нелидовский; 5 – Лесной
Выпуклая задняя сторона паракона (фен Pts1) чаще встречалась у рыжих полевок из Нелидовского, Зубцовского и Калининского р-нов – 0,66, 0,58 и 0,54. Доля М2 с прямой задней стороной паракона (Pts2) максимальна в выборке из Зубцовского р-на – 0,27, реже отмечался данный фен у зверьков из Лесного и Калининского р-нов – 0,13 и 0,12, и минимальна его встречаемость в Торопецком и Нелидовском р-нах – 0,06 и 0,05. Самая высокая встречаемость вариации Pts3 (вогнутая задняя сторона паракона) отмечена в выборке из Лесного р-на – 0,37, в сериях полевок из Торопецкого и Калининского р-нов доля этого фена одинакова и также достаточно высока – 0,22. В Нелидовском и Зубцовском р-нах фен Pts3 редок – 0,06 и 0,10. Примечательно, что задняя сторона паракона М2 с дополнительным эмалевым выступом (Pts4) наиболее обычна для рыжих полевок, обитающих в хвойных лесах: у зверьков из Торопецкого, Нелидовского и Лесного р-нов встречаемость фена соответственно 0,43, 0,23 и 0,17. У полевок из Зубцовского и Калининского р-нов доля моляров с этой вариацией – 0,05 и 0,12 (рис. 1; 4, а). По рассмотренному признаку все популяции достоверно отличались друг от друга, кроме популяций из Калининского и Лесного р-нов (р≤0,05, 0,01, 0,001).
При сопоставлении серий по признакам характера замкнутости зубных пространств М2 (Sai1, Sai2) было обнаружено уменьшение количества статистически значимых различий в направлении от передней непарной петли к заднему основному треугольнику. Так, для признака 1Sai1 получено шесть достоверных различий из десяти вариантов попарного сравнения выборок, для признака 2Sai1 – четыре и для признака 3Sai1 – три. Складывается впечатление, что для изучения фенотипических особенностей природных групп особей более информативным является первое разделение дентиновых полей. Другие два признака в большей степени связаны со степенью снашивания моляра, что делает их менее пригодными для популяционных исследований. В целях анализа различий выборок по признакам разделения дентиновых полей были получены средние доли случаев разделения дентиновых полей, рассчитанные как среднее арифметическое частот встречаемости фенов 1Sai1, 2Sai1 и 3Sai1. Для серий из Торопецкого, Зубцовского, Нелидовского, Калининского и Лесного р-нов они составили 0,67, 0,63, 0,58, 0,57 и 0,29 соответственно. Исходя из установленного нами факта, что при интенсивном стирании коронки зуба дентиновые поля вторично сливаются [7], можно предположить, что обнаруженное у зверьков из Нелидовского, Калининского и особенно Лесного р-нов уменьшение доли М2 с разделением зубных пространств обусловлено преобладанием в диете грубых кормов. Например, в питании полевок, отловленных летом 1999 г. в Калининском и Нелидовском р-нах, значительна доля зеленых частей растений, коры и корневищ – 87,5 и 84,6%. Для сравнения: у зверьков из Зубцовского и Торопецкого р-нов соответствующие доли составили 75,3 и 65,9%.
Ровная поверхность наружной поверхности талонуса М3 (Fet1) – преобладающая вариация в выборках из Зубцовского и Калининского р-нов, где её доли составили 0,47 и 0,48. В Торопецком, Лесном и Нелидовском р-нах встречаемость данного фена была гораздо ниже – 0,28, 0,23 и 0,22 соответственно (рис. 2; 4, б). Слабо выраженный изгиб наружной поверхности талонуса (Fet2) с одинаковой частотой встречался у зверьков из Торопецкого и Лесного р-нов – 0,43. Доля этого фена у полевок из Нелидовского, Зубцовского и Калининского р-нов также была примерно одинакова – 0,33, 0,33 и 0,30. У зверьков из Нелидовского р-на обнаружена максимальная доля М3 с дополнительным эмалевым бугорком на наружной поверхности талонуса (Fet3) – 0,20; в Торопецком и Зубцовском р-нах встречаемость данной вариации соответственно 0,11 и 0,13, а в выборках из Лесного и Калининского р-нов – 0,10 и 0,06. М3 с дополнительным острым выходящим углом на наружной поверхности талонуса (Fet4) чаще всего отмечался у полевок из Нелидовского и Лесного р-нов – 0,25 и 0,23, реже встречался фен в Торопецком и Калининском р-нах – 0,18 и 0,16, и минимальная его доля найдена в выборке из Зубцовского р-на – 0,07.
Отметим, что максимальная суммарная частота встречаемости вариаций, отражающих наибольшее усложнение (удлинение) эмалевого края наружной поверхности талонуса (сумма долей фенов Fet3 и Fet4), была зафиксирована у рыжих полевок из Нелидовского, Лесного и Торопецкого р-нов – 0,45, 0,33 и 0,29, минимальная – у зверьков из Калининского и Зубцовского р-нов – 0,22 и 0,20 (рис. 4, б). Статистически подтверждено отличие по частоте встречаемости вариаций этого признака выборки из Зубцовского р-на от серий из Лесного, Нелидовского и Торопецкого р-нов и различие серий из Нелидовского и Калининского р-нов (р≤0,05, 0,01, 0,001).
Анализ распределения в исследованных сериях вариаций строения внутренней поверхности талонуса (Fit) показал закономерности, в некотором роде обратные обнаруженным для строения наружной поверхности талонуса (рис. 2). Упрощенное строение внутренней поверхности талонуса, выражаемое суммой долей фенов Fit1 и Fit2, оказалось более характерным для выборок из Лесного и Нелидовского р-нов – 0,84 и 0,82, несколько реже оно встречалось у полевок из Торопецкого и Калининского р-нов – 0,79 и 0,78. Минимальная встречаемость талонуса с прямой или слабо изогнутой внутренней стороной отмечена в серии из Зубцовского р-на – 0,62. При этом доля самой сложной вариации Fit4 (дополнительный острый выходящий угол) максимальна у полевок из Зубцовского р-на (0,2) и минимальна у зверьков из Лесного р-на (0,03). По признаку Fit серия из Зубцовского р-на достоверно отличалась от серий из Нелидовского, Торопецкого и Лесного р-нов, также статистически подтвердилось различие выборок из Нелидовского и Торопецкого р-нов (р≤0,05, 0,01, 0,001).
Признаки Scai1, Scai2 отражают характер замкнутости зубных полей М3. По признакам 1Scai1 и 4Scai1 не обнаружено статистически значимых различий между сериями. При расчете средней встречаемости вариаций признаков 1Scai1 – 4Scai1 была получена следующая последовательность районов отлова, отражающая уменьшение доли М3 с замкнутыми дентиновыми пространствами: Торопецкий, Зубцовский, Нелидовский, Калининский и Лесной (средние доли соответственно 0,79, 0,76, 0,72, 0,71 и 0,6). Этот ряд полностью совпадает с подобной последовательностью районов, вычисленной на основании средней доли случаев разделения дентиновых полей М2, что свидетельствует о возможном влиянии на эти дискретные признаки особенностей кормовой базы.
Можно предположить, что отмеченные выше популяционные особенности распределения фенов внутренней поверхности талонуса также в той или иной степени зависят от степени снашивания коронки зуба и, следовательно, от особенностей рациона рыжих полевок. Вероятно, при совершении жевательных движений основная нагрузка падает именно на этот структурный элемент М3 и меньше – на наружную поверхность талонуса, вследствие чего дополнительные эмалевые выступы на лингвальной поверхности талонидного комплекса стачиваются вплоть до полного исчезновения.
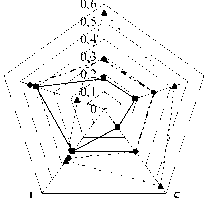
При сравнении серий по частоте встречаемости морфотипов М3 обращалось внимание на степень их сложности (рис. 5, б). Из данных, представленных в таблице, следует, что у полевок, отловленных в Торопецком, Лесном и Нелидовском р-нах, преобладали сложные фены e и f, суммарная встречаемость которых составила 0,55, 0,54 и 0,42. В выборке из Калининского р-на примерно в равных долях были представлены простые фены, средней и высокой степени сложности – 0,34, 0,30 и 0,36 соответственно. Серия из Зубцовского р-на характеризовалась преобладанием моляров простого строения (0,44) и редкостью М3 со сложным рисунком жевательной поверхности (0,16) (см. таблицу; рис.6, а). Различия большинства выборок статистически достоверны и достигли высокого уровня значимости (р≤0,01, 0,001).
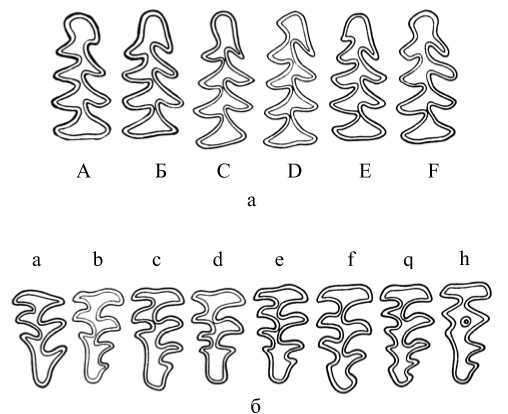
Рис. 5. Фены, выделяемые по форме жевательной поверхности моляров : а – первого нижнечелюстного (М 1 ); б – третьего верхнечелюстного (М 3 )
При отнесении морфотипов М1 к группам «простых» или «сложных» мы встретились с определенными трудностями, так как варианты форм зубов не четко альтернативны, а связаны многочисленными переходами в непрерывный ряд. Примером такого ряда являются фены B, C и F, которые достаточно сложно отделить друг от друга (рис. 5, а). Учитывая особенности строения параконидного комплекса и количество выступающих углов на наружной и особенно внутренней стороне зуба, по степени сложности морфотипы М 1 были распределены следующим образом: к «простым» отнесены А и В, к фенам средней степени сложности – C и F, «сложные» варианты строения М 1 – фены D и E.
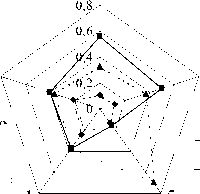
При подобном разнесении фенов анализ особенностей строения М 1 у зверьков из удаленных друг от друга районов Тверской обл. показал, что в сериях рыжих полевок из Калининского и Зубцовского р-нов преобладали моляры с упрощенным строением параконидного отдела, доля которых составила 0,24 и 0,22. У зверьков из Торопецкого, Нелидовского и Лесного р-нов фены А и В встречались редко – 0,11, 0,12 и 0,13 соответственно. При этом выборки из Торопецкого и Нелидовского р-нов характеризовались повышенным содержанием фенов средней сложности (C и F) – 0,56 и 0,50. Доля сложных фенов была максимальна у полевок из Лесного р-на – 0,70 (см. таблицу;
рис.6, б). Статистически подтвердилось различие большинства выборок (р≤0,01, 0,001), кроме серий из Нелидовского и Торопецкого р-нов, различия между которыми не достоверны.
а
простые средние сложные
простые средние
б
сложные
Доля морфотипов разной степени сложности в популяциях рыжей полевки Тверской области
|
Районы исследования |
N |
Морфотипы М3 |
Морфотипы М1 |
||||
|
Простые (a+b) |
Средняя степень сложности (c+d) |
Сложные (e+f) |
Простые (A+B) |
Средняя степень сложности (C+F) |
Сложные (D+E) |
||
|
Лесной |
50 |
0,30 |
0,13 |
0,54 |
0,13 |
0,16 |
0,70 |
|
Калининский |
100 |
0,34 |
0,30 |
0,36 |
0,24 |
0,38 |
0,38 |
|
Зубцовский |
100 |
0,44 |
0,40 |
0,16 |
0,22 |
0,41 |
0,37 |
|
Нелидовский |
100 |
0,30 |
0,19 |
0,42 |
0,12 |
0,50 |
0,38 |
|
Торопецкий |
100 |
0,28 |
0,17 |
0,55 |
0,11 |
0,56 |
0,33 |
Рис. 6. Доля морфотипов разной степени сложности в популяциях рыжей полевки Тверской обл.:
а – доля морфотипов М 3 ; б – доля морфотипов М 1 (обозначения районов см. рис. 4)
Интересен факт наличия клинальной изменчивости формы внутренней поверхности передней непарной петли параконида: обнаружено увеличение встречаемости угла на внутренней поверхности непарной петли параконида в направлении с юго-запада на северо-восток области (Jli4) (рис. 3). Доля этого фена в выборках из Торопецкого, Нелидовского, Зубцовского, Калининского и Лесного р-нов составила 0,25, 0,36, 0,41, 0,45 и 0,51. В этом же направлении отмечено соответственное уменьшение суммарных долей вариаций, отражающих упрощенное строение рассматриваемого элемента жевательной поверхности М 1 (Jli1, Jli2 и Jli3 – округлая, прямая и вогнутая форма). По данному признаку уровня достоверности достигло отличие полевок из Торопецкого р-на от зверьков из Нелидовского, Зубцовского и Лесного р-нов (р≤0,05, 0,01). При анализе распределения на территории Тверской обл. вариаций строения остальных элементов жевательной поверхности М 1 (признаки Ali, Aex, Ain, 1Cet, 2Cet, Phc, 1 Sci1-5Sci1 и др.) определенных закономерностей обнаружено не было.
В ранних исследованиях географической изменчивости состава и частоты встречаемости морфотипов М3 и М1 рыжей полевки было отмечено, что для популяций более специфичным является рисунок жевательной поверхности М 1 [6]. В свете этого особый интерес представляло определение уровня сходства рассматриваемых нами популяций по встречаемости морфотипов М 1 . Полученные данные свидетельствуют, 86
что полевки из Лесного р-на наиболее сходны со зверьками из Нелидовского (r=0,88) и Торопецкого (r=0,82) р-нов. При сопоставлении с выборками из Калининского и Зуб-цовского р-нов получены показатели сходства r=0,77 и 0,72.
В целом по признакам, описывающим рисунок жевательной поверхности М1, все выборки достоверно отличались друг от друга. Наибольшее сходство обнаружено между сериями из Зубцовского, Нелидовского и Торопецкого р-нов (r=0,98 и 0,97). Рыжие полевки из Калининского р-на наиболее сходны со зверьками из Зубцовского р-на (r=0,96). Для выборки из Лесного р-на несколько бóльшие значения показателя сходства получены при сопоставлении с сериями из Нелидовского и Торопецкого р-нов (r=0,96 и 0,93), и мéньшие – при сравнении с сериями из Калининского и Зубцовского р-нов (r=0,92).
Таким образом, подтверждается сходство строения моляров рыжих полевок, обитающих в хвойных лесах, с одной стороны, и структурных особенностей коренных зубов полевок из смешанных лесов – с другой.
Из результатов анализа фенетических особенностей выборок европейской рыжей полевки из разных точек Тверской обл. следует, что специфика строения некоторых элементов жевательной поверхности коренных зубов у данного вида обусловлена био-топическими особенностями местообитания. Обнаружено значительное сходство фенофондов у рыжих полевок, места обитания которых приурочены к хвойным лесам, находящимся в зоне елово-широколиственных и таежных лесов. В данных районах наблюдается увеличение складчатости боковых сторон коренных зубов, приводящее к удлинению их режущего края. Особенно ярко выражается эта закономерность в строении талонидного комплекса М3 и параконидного отдела М 1 . Наибольший уровень различий фенофондов получен при сравнении зверьков, обитающих в мелколиственных и смешанных лесах, где увеличена встречаемость моляров упрощенного строения, с рыжими полевками, отловленными в хвойных лесах.
Выявлена клинальная изменчивость формы внутренней поверхности передней непарной петли параконида в пределах исследованной области, соответствующая переходу от зоны хвойно-широколиственных лесов к таежным лесам: встречаемость М 1 с упрощенным строением данного элемента жевательной поверхности в пределах Тверской обл. уменьшается в направлении с юго-запада на северо-восток. По данному признаку, в проявлении которого высока доля генотипической изменчивости [10], статистически значимо отличие серии полевок из крайнего западного района области (Торопецкий) от остальных выборок.
Вероятна зависимость ряда дискретных признаков зубной системы европейской рыжей полевки, таких, как число замкнутых дентиновых полей М2 и М3 и строение внутренней поверхности талонуса М3, от степени снашивания моляров.
СПИСОК ЛИТЕРАТУРЫ полевки (Clethrionomys glareolus Schreber) // Вестн. Твер. гос. ун-та. Сер. Биология и экология. 2005. Вып. 1, № 4 (10). С. 88–96.
-
8. Животовский Л.А. Показатель сходства популяций по полиморфным признакам // Журн. общ. биол. 1979. Т.40, №4. С. 587–602.
-
9. Животовский Л.А. Показатель внутрипопуляционного разнообразия // Журн. общ. биол. 1980. Т.41, №6. С. 828–836.
-
10. Котляров О.Н. Возрастная изменчивость морфологических признаков, используемых в фенетике популяций грызунов // Фенетика популяций: Материалы III Всесоюз. совещания. М., 1985. С. 220–222.
-
11. Кузнецов Б.А. Очерк зоогеографического районирования СССР. М., 1950. Нов.серия. Вып. 20 (35).
-
12. Ларина Н.И., Ерёмина И.В. Каталоги основных вариаций краниологических признаков у грызунов // Фенетика природных популяций. М., 1988. С. 8–52.
-
13. Невский М.Л. Леса Калининской области // Природа и хозяйство Калининской области. Калинин, 1960. С. 323–390.
-
14. Огнев С.И. Звери СССР и прилежащих стран // Грызуны. М.; Л., 1950. Т.7.
-
15. Соколов В.Е., Суханов В.Б. Задачи и программа раздела «Морфология» в монографиях серии «Виды фауны СССР и сопредельных стран» // Материалы VII (XV) и VIII (XVI) заседаний Рабочей группы по проекту №8б(18) «Вид и его продуктивность в ареале». Вильнюс, 1978. С. 20–24.
-
16. Шапошников Л.В. Звери Вышневолоцкого района // Учен. зап. КГПИ. Калинин, 1956. Т. 20. С. 159–183.
-
17. Шапошников Л.В. О зоогеографическом районировании территории Калининской области // Материалы III совещания по естественно-историческому и экономикогеографическому районированию СССР для целей сельского хозяйства. М., 1959. С. 120–121.
-
18. Яблоков А.В. Фенетика. Эволюция, популяция, признак. М., 1980.
-
19. Steven D.M. Recent evolution in the genus Clethrionomys // Symp. Soc. Exp. Biol. 1953. V. 7. Evolution. P. 310–319.
-
20. Zimmermann Z. Zur Evolution der molaren Struktur der Erdmaus, Microtus agrestis (L.) // Zool. Jahrb. Syst. 1956. Bd. 84. S. 269–274.
-
21. Zimmermann Z. Selektionswart der Simplex – Jahrform bei der Feldmaus (Eine Engegnung) // Zool. Jahrb. Syst. 1958. Bd. 86. S. 35–40.
-
22. Zimmermann Z. Über eine Krezung von Unterarten der Feldmaus Microtus arvalis // Zool. Jahrb. Syst. 1959. Bd. 87. S. 1–12.
SOME TRENDS IN POLYMORPHIC VARIATION OF ODONTOLOGICAL CHARACTERS OF EUROPEAN BANK VOLE (CLETHRIONOMYS GLAREOLUS SCHREBER) FROM UPPER VOLGA AND ADJACENT AREAS
Tver State University
Article deals with variations in molars’ occlusive surfaces of European Bank Vole, inhabiting Tver region. Some differences between voles from coniferous forests and those from mixed ones have been found. A cline variation in morphology of the first unpaired loop of paraconid along with dependant occurrence of some characters during progressive wear have been found.
