Неоднородность структурной организации ценопопуляций Artemisia hololeuca Bieb. ex Bess. (Asteraceae) во времени и пространстве
 во времени и пространстве")
Автор: Сидорова Л.А.
Журнал: Природные системы и ресурсы @ns-jvolsu
Рубрика: Биология и биотехнология
Статья в выпуске: 1 (1), 2011 года.
Бесплатный доступ
Исследование ценопопуляций Artemisia hololeuca Bieb. ex Bess. показало наличие скоплений различных уровней агрегированности, отличающихся целым рядом качественных признаков (возрастной состав, плотность, протяженность, дискретность). Асинхронное и волнообразное развитие скоплений A. hololeuca в континууме «время - пространство» объясняется биологией вида и целым рядом средообразующих факторов: экспозицией и крутизной склона, микрорельефом и составом почвенного субстрата, фитоценологическими особенностями растительного сообщества, а также характером и степенью антропогенного воздействия. Установлена высокая степень динамичности ценопопуляций изученного вида.
Стержнепридаточнокорневой полукустарничек, возрастные (онтогенетические) группы, ценопопуляции, скопления, уровни агрегированности, групповой тип размещения
Короткий адрес: https://sciup.org/14967323
IDR: 14967323 | УДК: 581.524.3:581.524
Heterogeneity of structure organization of cenopopulation of Artemisia hololeuca Bieb. ex Bess. (Asteraceae) during the time and in the space
The investigation of cenopopulation of Artemisia hololeuca Bieb. ex Bess. rerealed the existents of cooperations of the different levers of aggregations, different the whole ray of quality especialities such age consistence, density, alongeness, discontinuity. Non synchrony and ware resembling development of aggregation of A. hololeuca the continuum of time and space is explained the biology of the species and by the whole ray the surroundings forming factors, sail au exposition stope, microrelief and composition of earth, substrate, fitocenological especiality. of the plant cooperation and also the character and degree of anthropogenic influence. Is determined the high degree of dynamic of development cenopopulation of the tipe investigated.
Текст научной статьи Неоднородность структурной организации ценопопуляций Artemisia hololeuca Bieb. ex Bess. (Asteraceae) во времени и пространстве
Любой биологический вид существует одновременно во времени и пространстве, именно изучение биологической системы в совокупности ее пространственно-временных связей позволяет понять ее структуру и функционирование [3].
В качестве модельного объекта нашего исследования был выбран стержнепридаточнокорневой полукустарничек Artemisia hololeuca Bieb. ex Bess. Этот вид интересен для ценопопуляционных исследований во времени и пространстве по следующим причинам: вид на исследуемой территории встречается в большом обилии; в ценопопуляции легко выделить возрастные группы особей; размножается как семенами, так и вегетативно. У A. hololeuca выявлено два способа вегетативного размножения – с помощью уко-рененениия стержнепридаточных особей (в основном, v, g1) и сенильно-виргинильными особями (s-v/), которые представляют собой вегетативные партикулы распадающихся се- нильных и субсенильных особей. Последняя возрастная группа может переходить к кратковременной генерации, то есть здесь мы имеем дело с явлением частичного омоложения старческих особей [5].
Artemisia hololeuca обладает узким ареалом – произрастает вдоль течения р. Дона и его притоков; на территории Волгоградской области встречается исключительно на северо-западе области, на самой северной границе своего современного ареала распространения [2]. На протяжении всего своего ареала вид приурочен исключительно к коренным выходам мела, занимая крутые части обнажений («взлобья»), выступая доминантом в растительных сообществах [1, с. 34–36; 2].
Работа проводилась на территории Нижнехоперского природного парка (Волгоградская область, Алексеевский район, урочище «Дунина гора») на участках ассоциации Artemisia salsoloides + Artemisia salsoloides, с участием ксерофильного разнотравья (Thymus calcareus, Silene cretacea, Pimpinella titanophila, Hedysarum grandiflorum, Scabiosa isetensis, Jurinea ledebourii, Stipa pennata и др.). Площадь почвы, свободной от растительного покрова, составляет 30–50 %; обилие A. hololeuca варьирует в пределах sol. – cop3. Исследуемые ценопопуляции занимают небольшое по площади меловое плато несколько асимметричного строения южной и юго-западной экспозиции; субстрат представлен малоразвитой выветрившейся меловой породой. Территория урочища Дунина гора подвержена значительному антропогенному влиянию (разработка меловых карьеров, усиленный выпас крупного и мелкого рогатого скота).
A. hololeuca – малолетник, длительность онтогенеза которого составляет в среднем 8– 10 лет. В онтогенезе полыни беловойлочной выделяют следующие возрастные состояния: проростки ( p ); ювенильные ( j ); имматурные ( im ); виргинильные ( v ); молодые ( g1 ), среднегенеративные ( g2 ) и старые генеративные ( g3 ); субсенильные ( ss ) и сенильно-виргинильные ( s-v/ ) (рис. 1). Календарный возраст особей: p – один вегетационный сезон, j – 4–5 недель, im – 1 год, v – 2–3 года , g1 – 1–2 года, g2 – 4– 5 лет, g3 – 1–3 года, ss – 1–2 года, s-v/ – 2– 4 года [4, с. 8–10].
Первой методической задачей при изучении ценопопуляций в континууме «пространство – время» стало выявление участков с более высокой (скопления) и с более низкой (промежутки между скоплениями) плотностью особей; вторая задача – изучение базовых параметров пространственной структуры скоплений. Для реше- ния данных задач закладывались стационарные площадки через всю территорию обнаруженной популяции с дальнейшим геоботаническим описанием и картированием [6, с. 16–68].
Во всех исследованных ценопопуляци-ях A. hololeuca в пределах стационарных участков было установлено от двух до трех уровней агрегированности (табл. 1). Уровни агрегированности (I–III) формируются на последовательно расположенных участках площадью 0,25, 0,5 и 1 кв. м соответственно и носят групповой тип пространственного размещения. Скопления на более крупных площадках (1,5, 2 кв. м и более) являются слабо дискретными, и в целом пространственное размещение может относиться к диффузному типу.
Плотность скоплений I порядка наибольшая по сравнению с аналогичными показателями скоплений II–III порядков; коэффициент плотности в пределах 2,6–4,2, что свидетельствует о явно выраженном «центре» скопления. Скопления более высоких рангов (II–III порядка) характеризуются более низкой плотностью особей, большими размерами площади и диффузно-групповым размещением. В таких скоплениях наблюдается частичное слияние взрослых особей друг с другом, образующее сплошной покров из вегетативных, генеративных и сенильно-вегетативных особей.
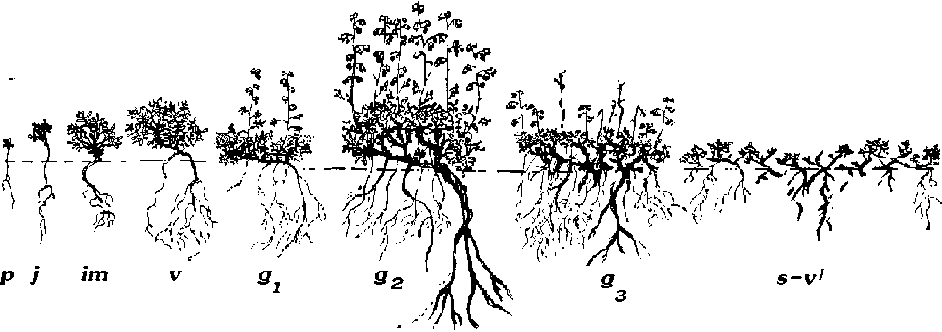
Рис. 1. Схема возрастных состояний в онтогенезе Artemisia hololeuca Bieb. ex Bess. (по Сидоровой, 2008):
p – проростки; j – ювенильные; im – имматурные; v – виргинильные; g1 – молодые генеративные;
g2 – среднегенеративные; g – старогенеративные; sv / – сенильные, сенильно-виргинильные растения
Таблица 1
Основные параметры пространственной структуры особей A. hololeuca в ценопопуляциях (урочище «Дунина гора»)
|
№ ЦП |
Уровень агрегированности |
Размер площадок, кв. м |
Плотность особей в скоплениях, шт. |
Плотность особей между скоплениями |
Средняя площадь одного скопления, кв м |
Протяженность скопления, м |
Дискретность |
|
|
D M |
D L |
|||||||
|
1 |
I |
0,25 |
2,13 ± 0,43 |
0,15 ± 0,03 |
0,06 ± 0,01 |
0,22 ± 0,03 |
0,93 |
0,45 |
|
II |
0,5 |
0,80 ± 0,17 |
0,25 ± 0,12 |
0,18 ± 0,04 |
0,49 ± 0,02 |
0,69 |
0,37 |
|
|
III |
1 |
0,51 ± 0,04 |
0,23 ± 0,06 |
0,50 ± 0,05 |
1,28 ± 0,06 |
0,55 |
0,35 |
|
|
2 |
I |
0,25 |
1,17 ± 0,18 |
0,13 ± 0,03 |
0,05 ± 0,003 |
0,25 ± 0,02 |
0,89 |
0,53 |
|
II |
0,5 |
0,45 ± 0,18 |
0,10 ± 0,03 |
0,20 ± 0,01 |
0,49 ± 0,03 |
0,78 |
0,38 |
|
|
III |
1 |
Не выделен |
||||||
|
3 |
I |
0,25 |
1,33 ± 0,22 |
0,16 ± 0,03 |
0,05 ± 0,01 |
0,30 ± 0,002 |
0,89 |
0,33 |
|
II |
0,5 |
0,56 ± 0,06 |
0,15 ± 0,03 |
0,21 ± 0,03 |
0,58 ± 0,04 |
0,73 |
0,28 |
|
|
III |
1 |
0,40 ± 0,04 |
0,27 ± 0,07 |
0,53 ± 0,07 |
0,97 ± 0,02 |
0,33 |
0,25 |
|
Основными формообразующими факторами, влияющими на пространственную мозаику скоплений A. hololeuca , являются орографические, эдафические факторы и биологические особенности вида. По мере развития генеративной особи происходит старение и отмирание ее отдельных парциалей в центральной части. Дальнейшее развитие особи происходит в периферийной части преимущественно вегетативно. Появление нескольких центров в скоплениях II–III уровней связано с развитием особей как семенного, так и возникновения и развития особей вегетативного происхождения (v, s-v/). В связи с особенностями онтоморфогенеза одна среднегенеративная особь может занимать в среднем 0,25– 0,60 кв. м. Исследуя структуру скоплений A. hololeuca , можно говорить о полицентрическом варианте скоплений более высоких уровней агрегированности.
Предметом дальнейших исследований послужили возрастные спектры различных уровней агрегированности, характеризующиеся большим разнообразием и относительной вариабельностью (табл. 2). В пределах исследованных стационарных площадок удалось выявить 46 скоплений I уровня агрегирован-ности, 21 скопление II уровня агрегированно-сти и 14 скоплений III уровня агрегированнос-ти и проследить изменение возрастных спектров этих скоплений на протяжении 5 лет. В табл. 2 приведены материалы по одному скоплению каждого уровня агрегированности в каждой конкретной ценопопуляции.
Исследование возрастных спектров скоплений в течение пяти полевых сезонов показывает, что в большинстве своем скопления всех трех уровней агрегированности являются неполночленными (то есть особи некоторых возрастных групп в них не представлены). Исключение составляют скопления III уровня агрегированности в ЦП 1 в период с 2007 по 2008 год. Возрастные спектры скоплений II уровня агреги-рованности отличаются большим постоянством и стабильностью, чем скопления I и III порядков.
Возрастные спектры скоплений можно систематизировать следующим образом: 1) спектры с одним максимумом (мономо-дальные), приходящимся на im , v , g2 , s-v/ ; спектры с двумя максимумами (бимодальные), где сочетания максимумов довольно разнообразны – v и g2 , v и s-v /, g2 и s-v /.
Такое разнообразие возрастных спектров в скоплениях можно объяснить несинхронным развитием особей в скоплениях, что создает картину неоднородности скоплений по возрастному спектру при ежегодных одноразовых наблюдениях. Асинхронность развития во многом определяется онтоморфо-логическими особенностями A. hololeuca , а именно, поливариантностью развития вида, реализацией полного и неполного типа прохождения онтогенеза, наличием двух способов возобновления, где доминирующее положение занимают сенильно-виргиниль-ные особи.
Таблица 2
Возрастная динамика скоплений A. hololeuca разных уровней агрегированности (2006–2010 гг.)
|
№ ЦП |
Уровень агрегированности |
Сроки наблюдений * |
Возрастные группы, % |
||||||||
|
p |
j |
im |
v |
g 1 |
g 2 |
g 3 |
ss , s |
/ sv |
|||
|
1 |
I |
1 |
16,7 |
16,7 |
25,0 |
18,3 |
16,7 |
8,3 |
8,3 |
– |
– |
|
2 |
– |
– |
15,4 |
15,4 |
15,4 |
7,6 |
7,7 |
– |
38,5 |
||
|
3 |
23,5 |
– |
11,8 |
11,8 |
17,6 |
5,9 |
5,9 |
– |
23,5 |
||
|
4 |
– |
– |
8,3 |
16,7 |
16,7 |
8,3 |
– |
8,3 |
41,7 |
||
|
5 |
– |
– |
– |
33,3 |
– |
11,0 |
– |
11,1 |
44,6 |
||
|
II |
1 |
12,5 |
12,5 |
18,8 |
12,5 |
18,6 |
18,8 |
6,3 |
– |
– |
|
|
2 |
– |
– |
11,8 |
11,8 |
17,5 |
11,8 |
5,9 |
– |
41,2 |
||
|
3 |
18,2 |
9,1 |
4,5 |
9,1 |
22,7 |
9,1 |
4,5 |
– |
22,7 |
||
|
4 |
– |
– |
5,9 |
11,8 |
17,6 |
11,8 |
– |
5,9 |
47,0 |
||
|
5 |
– |
– |
– |
13,3 |
6,7 |
13,3 |
6,7 |
6,7 |
53,3 |
||
|
III |
1 |
6,7 |
6,7 |
13,3 |
20,0 |
13,3 |
33.3 |
6,7 |
– |
– |
|
|
2 |
9,3 |
4,7 |
7,0 |
16,3 |
9,1 |
28,0 |
4,7 |
2,3 |
18,6 |
||
|
3 |
13,5 |
5,8 |
1,9 |
17.3 |
11,5 |
17,3 |
7,7 |
3,8 |
21,2 |
||
|
4 |
– |
– |
5,3 |
15,8 |
15,8 |
10,5 |
5,3 |
5,3 |
42,1 |
||
|
5 |
– |
– |
– |
15,8 |
10,5 |
10,5 |
5,3 |
5,3 |
52,6 |
||
|
2 |
I |
1 |
– |
20,0 |
– |
20,0 |
20,0 |
40,0 |
– |
– |
– |
|
2 |
18,2 |
– |
9,0 |
9,1 |
9,1 |
18,2 |
– |
9,1 |
27,3 |
||
|
3 |
– |
– |
– |
18,2 |
– |
18,2 |
9,0 |
9,1 |
45,5 |
||
|
4 |
– |
– |
– |
11,1 |
22,0 |
22,4 |
11,0 |
11,2 |
22,2 |
||
|
5 |
– |
– |
– |
– |
10,0 |
30,0 |
10,0 |
10,0 |
40,0 |
||
|
II |
1 |
– |
45,4 |
18,2 |
9,1 |
9,1 |
18,2 |
– |
– |
– |
|
|
2 |
15,4 |
15,4 |
7,7 |
7,7 |
15,3 |
15,4 |
– |
7,7 |
15,4 |
||
|
3 |
– |
– |
– |
15,4 |
7,6 |
15,0 |
7,7 |
7,7 |
46,6 |
||
|
4 |
– |
– |
– |
18,2 |
18,2 |
18,2 |
9,1 |
9,0 |
27,3 |
||
|
5 |
– |
– |
– |
– |
15,4 |
23,1 |
7,7 |
7,6 |
46,2 |
||
|
III |
Не выявлен |
||||||||||
|
3 |
I |
1 |
6,8 |
18,2 |
70,5 |
4,5 |
– |
– |
– |
– |
– |
|
2 |
10,9 |
13,0 |
52,2 |
15,2 |
6,5 |
2,2 |
– |
– |
– |
||
|
3 |
4,3 |
17,0 |
31,9 |
29,8 |
10,6 |
6,4 |
– |
– |
– |
||
|
4 |
– |
4,4 |
40,0 |
22,2 |
17,8 |
11,1 |
2,3 |
– |
2,2 |
||
|
5 |
– |
– |
25,0 |
16,6 |
30,6 |
11,1 |
5,6 |
2,8 |
8,3 |
||
|
II |
1 |
6,1 |
16,3 |
63,3 |
8,2 |
– |
6,1 |
– |
– |
– |
|
|
2 |
8,8 |
12,3 |
50,9 |
12,3 |
8,8 |
7,0 |
– |
– |
– |
||
|
3 |
3,5 |
14,3 |
30,4 |
25,0 |
14,3 |
10,7 |
1,8 |
– |
– |
||
|
4 |
– |
3,9 |
37,3 |
21,6 |
19,6 |
11,8 |
2,0 |
1,8 |
2,0 |
||
|
5 |
– |
– |
19,6 |
26,1 |
21,7 |
13,0 |
2,2 |
2,2 |
15,2 |
||
|
III |
1 |
4,0 |
10,7 |
72,0 |
5,3 |
4,0 |
4,0 |
– |
– |
– |
|
|
2 |
7,7 |
15,4 |
47,7 |
12,3 |
7,7 |
7,7 |
1,5 |
– |
– |
||
|
3 |
3,3 |
14,8 |
29,5 |
26,2 |
13,1 |
11,5 |
1,6 |
– |
– |
||
|
4 |
– |
5,4 |
33,9 |
19,6 |
17,9 |
12,5 |
1,8 |
1,8 |
7,1 |
||
|
5 |
– |
– |
21,4 |
31,0 |
7,1 |
14,3 |
2,4 |
2,4 |
21,4 |
||
* Сроки наблюдений: 1 – 02.07.2006; 2 – 18.06.2007; 3 – 10.05.2008; 4 – 12.08.2009; 5 – 27.08.2010.
Различия возрастных спектров одного и того же скопления в разные моменты связаны еще и по причине неравномерности процессов семенного возобновления и приживаемости проростков на разных участках ценопопуляции. Всходы полыни беловойлочной могут появляться весной и осенью при достаточной влажности почвенного субстрата и особенностей микрорельефа. Наблю- дения в течение нескольких полевых сезонов показали, что степень закрепления и дальнейшего развития проростков данного вида весьма низка.
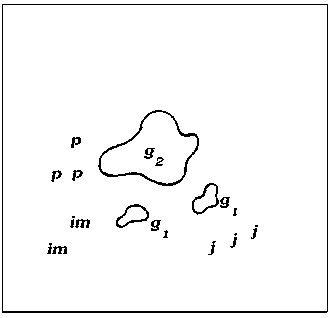
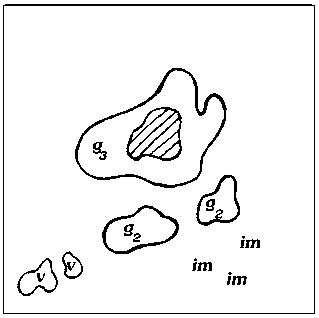
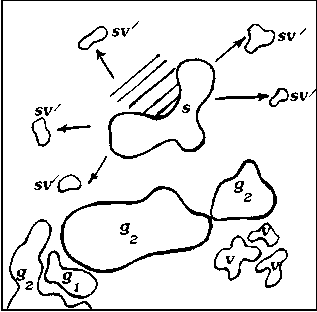
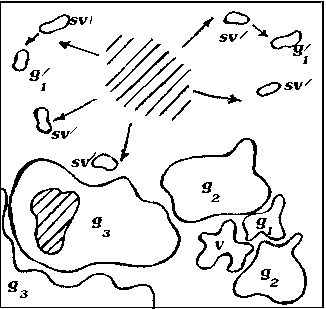
Исходя из анализа пространственно-возрастной структуры A. hololeuca , а также исходя из теоретических рассуждений, можно построить обобщенную схему, иллюстрирующую основные этапы формирования одного скопления (рис. 2).
А
Б
В
Г
Рис. 2. Обобщенная схема основных этапов развития скоплений A. hololeuca :
А – Г – основные этапы формирования скопления: А – скопление I уровня агрегированности; Б – В – скопления II уровня агрегированности, Г – скопление III уровня агрегированности – границы взрослого растения; – место и степень отмирания особи
Образование скоплений самого низкого уровня у A. hololeuca связано непосредственно с типом и характером возобновления. Полынь беловойлочная типичный анемохор, у которой семена рассеиваются на достаточно дальнее расстояние от материнской особи (g1 – g3) и прорастают либо в непосредственной близости от материнской особи, либо на некотором отдалении от нее. На первых этапах своего развития скопления A. hololeuca представлены ювенильными, имматурными и незначительным количеством вегетативных особей и генеративных особей (рис. 2, А). Формирование такого скопления занимает в среднем 1– 3 года и соответствует I уровню агрегирован-ности. Скопления такого рода характеризуются высокой нестабильностью, что связано с невысокой приживаемостью и значительной гибелью всходов. Затем, по мере онтогенетического развития вида, структура скоплений зна- чительно усложняется: скопления становятся разновозрастными или взрослыми за счет перехода большей части прегенеративных особей в генеративную стадию. Скопления незначительно стареют (g2→g3; g3→ss; ss→s) (рис. 2, Б, В). Отличительной особенностью такого скопления является быстрое формирование большой наземной биомассы за счет вегетативных и генеративных особей. На данном этапе развития возможны процессы частичного разрушения генеративных особей, образование субсенильных и сенильных особей, дающих сенильно-виргинильные особи. Скопления соответствуют II уровню агрегированнос-ти и формируются в течение 3–6 лет. Последний этап формирования скопления – полная или частичная дегенерация старых особей, образование большого количества сенильно-вир-гинильных парциалей. Такие скопления носят диффузно-групповой характер, занимают боль- шую площадь и частично сливаются с другими скоплениями (рис. 2, Г).
Таким образом, изучение отдельных скоплений в ценопопуляциях A. hololeuca позволяет сделать следующие выводы:
-
1. В пределах ценопопуляций Artemisia hololeuca выделяются скопления особей, которые характеризуются плотностью, протяженностью, степенью дискретности, площадью и возрастным спектром. Скопления разного уровня агрегированности различаются целым рядом количественных параметров. Выделяются от двух до трех уровней агрегированности.
-
2. Тип размещения особей в ценопопуляциях носит групповой и диффузно-групповой характер.
-
3. Изменение возрастного спектра отдельных скоплений во времени носит тот же характер и формы, что и изменения строения скоплений в пространстве. Здесь можно говорить о динамике скоплений ценопопуляций в едином континууме «пространство – время».
-
4. Возрастные спектры разных скоплений несходны друг с другом, что объясняется асинхронным развитием их во времени и пространстве. Асинхронность развития скоплений зависит от целого ряда факторов: особенностей биологии вида, эдафических и орографических условий природных местообитаний.
-
5. Природные ценопопуляции A. hololeuca весьма динамичны во времени и пространстве, что позволяет виду быстро освоить новые местообитания, долго существовать на уже за-
- нятых территориях, повышая тем самым адаптационный потенциал вида.
Список литературы Неоднородность структурной организации ценопопуляций Artemisia hololeuca Bieb. ex Bess. (Asteraceae) во времени и пространстве
- Володина, Н. Г. Флора меловых обнажений Волгоградской области/Н. Г. Володина//Флора степей и полупустынь (На примере Нижнего Поволжья): сб. ст. -Волгоград, 1982. -С. 34-46.
- Литвинова, Н. П. Об ареалах трех эндемичных видов меловых обнажений Русской равнины/Н. П. Литвинова, О. С. Горшкова//Проблемы экологии, геоботаники, ботанической географии и флористики. -Л.: Наука, 1977. -С. 151-155.
- Мазинг, В. В. Что такое структура биогеоценоза/В. В. Мазинг//Проблемы биогеоценологии. -М.: Наука, 1973. -С. 148-157.
- Сидорова, Л. А. Биоценопопуляционные исследования Artemisia hololeuca Bieb. ex Bess. (Asteraceae) и Silene cretacea Fisch. ex Spreng. (Caryophyllaceae) на территории Волгоградской области: автореф. дис.... канд. биол. наук: 03.00.05/Сидорова Людмила Алексеевна. -Астрахань, 2008. -23 с.
- Сидорова, Л. А. Особенности жизненной формы и биологии вегетативного размножения полыни беловойлочной Artemisia hololeuca Bieb. ex Bess. (Asteraceae) на меловых обнажениях Волгоградской области/Л. А. Сидорова//Современные проблемы ботаники: конф., посвящ. памяти В.В. Благовещенского (Ульяновск, 28 февр. -1 марта 2007 г.): сб. науч. ст. -Ульяновск: УлГПУ, 2007. -С. 305-311.
- Ценопопуляции растений (развитие и взаимоотношения). -М.: Наука, 1977. -131 с.
