Непарный шелкопряд как ключевой вид грызущих филлофагов в березовых лесах Западной Сибири

Автор: Максимов Сергей Алексеевич, Марущак Валерий Николаевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: фауна
Статья в выпуске: 3-3 т.15, 2013 года.
Бесплатный доступ
В статье описаны основные особенности механизма массовых размножений непарного шелкопряда в березовых лесах Западной Сибири. Со вспышками массовых размножений непарного шелкопряда связано повышение численности как его консументов, так и нескольких видов бабочек из сем. Zygaenidae и Nymphalidae. Очагами массового размножения непарного шелкопряда становятся, как правило, насаждения, растущие на бедных почвах. В тоже время очаги филлофага выделяются своим биоразнообразием.
Береза, непарный шелкопряд, сосущие корни, погодные факторы, массовое размножение, биоразнообразие
Короткий адрес: https://sciup.org/148201920
IDR: 148201920 | УДК: 595.787:591.526+574.472
Gypsy moth as the key type of gnawing phillofags in birch forests of West Siberia
In article the main features of the mechanism of mass reproductions of gypsy moth in birch forests of West Siberia are described. Increase of number both its consuments and several species of butterflies families Zygaenidae and Nymphalidae is connected with flashes of mass reproductions of gypsy moth. The plantings growing on poor soils become the centers of mass reproduction of gypsy moth as a rule. In the same time centers of phyllofag are allocated with the biodiversity.
Текст научной статьи Непарный шелкопряд как ключевой вид грызущих филлофагов в березовых лесах Западной Сибири
Большая часть территории Урала покрыта лесами. В южной части Уральского региона в лесостепной зоне Свердловской, Тюменской, Курганской областей и даже в южной степной половине Челябинской области березовые и в меньшей степени сосновые насаждения занимают значительные площади. Березовые леса восточной половины Урала относятся к зоне Западносибирских березовых лесов. Заметным компонентом животного населения березовых насаждений являются грызущие филлофаги. К ним относятся несколько десятков видов чешуекрылых и пилильщиков, питающихся листьями березы и ведущих открытый образ жизни. Многие виды грызущих филлофагов, а возможно, и все их виды, дают вспышки массового размножения. Наиболее известным филлофа-гом, консументом березы, являются непарный шелкопряд (Lyman-tria dispar L.), который периодически дефолиирует лиственные леса умеренной климатической зоны всего Северного полушария [11]. В Зауралье вспышки численности непарного шелкопряда возникают часто. На Западном склоне Урала они случаются гораздо реже.
С непарным шелкопрядом в березовых насаждениях Урала нередко проводится борьба, в ходе которой очаги массового размножения фил-лофага обрабатываются инсектицидами на больших площадях. Так, в 1996-1997 гг. на юге Тюменской области очаги непарника были обработаны пиретроидными препаратами на площади 130 тыс. га, а в 2011 г. в Курганской области и на юге Челябинской области – на площади 400 тыс. га. На эти мероприятия в общей сложности было затрачено около 200 млн. руб. В настоящее время в работах по динамике численности непарного шелкопряда в качестве синонимов видового названия чешуекрылого употребляется слова «филлофаг» и «вредитель». Так поступают и авторы настоящей статьи.
Максимов Сергей Алексеевич, кандидат биологических наук, научный сотрудник
Однако слово «вредитель» мы используем с чувством неловкости и сильного внутреннего сопротивления, так как непарный шелкопряд, вообще говоря, не наносит вреда кормовому растению. После дефолиации непарным шелкопрядом в конце июня березы, как правило, через 2 недели восстанавливают листву и благополучно проходят нормальный вегетационный цикл. Например, в 1996 г. на юге Тюменской области объеденная непарником листва уже к июлю восстановилась, а в 1997 г. численность филлофага снизилась сама по себе [7]. В 2011 г. в Челябинской области дефолиированные непарным шелкопрядом насаждения также полностью восстановили листву, на следующий же год дефолиации они не подвергались [9]. В то же время деревья, потерявшие листву в очагах непарника в 2010 г., внезапно погибли, что и послужило основанием для планирования широкомасштабных мероприятий по борьбе с непарным шелкопрядом в Челябинской и Курганской областях в 2011 г.
Динамика численности непарного шелкопряда, как и других видов грызущих филлофагов, отличается непредсказуемостью. Его очаги возникают всегда неожиданно. Численность филлофага в очагах может внезапно падать, а затем возобновляет свой рост. Иногда плотность популяций не-парника поддерживается в насаждении на высоком уровне в течение 10 и более лет, а иногда она сильно возрастает в течение 1 года, а на следующий год так же быстро снижается. Известный лесной энтомолог Н.Г. Коломиец, выступая в 1996 г. перед работниками Тюменского управления лесами, высказал мнение, что вспышки массового размножения грызущих филлофагов, в том числе непарного шелкопряда, относятся к принципиально непредсказуемым явлениям. Изучая динамику популяции грызущих филлофагов Урала, мы пришли к выводу, что у каждого их вида имеется свой механизм массовых размножений. В основе механизмов массовых размножений хвое-листогрызущих вредителей лежит не описанное до сих пор явление. Если распускание почек или интенсивный рост побегов у кормовых растений совпадают по времени с началом роста нового поколения сосущих корней, а деревья в этот момент находятся в состоянии водного стресса, то рост корней нарушается, и данное поколение тонких корней развивается слабым или даже не вырастает совсем. Средний срок жизни сосущих корней у наших древесных пород составляет 4 года [4]. В насаждении, где оказалось не развившимся одно из поколений сосущих корней, на 4 года возникает их недостаток – оно становится очагом массового размножения. Личинки филлофага, питающиеся кормовым растением с недостатком сосущих корней, имеют повышенную выживаемость, что и служит причиной роста численности насекомого в очаге [6].
Знание механизма массовых размножений непарного шелкопряда позволяет объяснить все особенности динамики популяций филлофага, предсказывать возникновение его очагов и протекание вспышек массового размножения. Оно позволяет даже предсказывать гибель деревьев после дефолиации гусеницами непарника [7]. Кроме того, со вспышками массового размножения непарного шелкопряда связано повышение биоразнообразия в березовых насаждениях. Этот аспект массовых размножений непарного шелкопряда по существу оставался вне поля зрения энтомологов. Вместе с тем он позволяет взглянуть на вспышки численности филлофага другими глазами, и по нашему мнению, заслуживает внимания исследователей.
Цель работы: описать те особенности возникновения очагов массового размножения непарного шелкопряда, которые имеют отношение к повышению биоразнообразия в березовых насаждениях Урала во время массовых размножений.
Материалы и методика. Работа проводилась в 1986-2013 гг. на юге Свердловской области, а также в Челябинской, Курганской, Тюменской областях. Методика работы включала ежегодные учеты непарного шелкопряда по яйцекладкам на большом числе постоянных пробных площадей; ежегодное составление таблиц выживания филло-фага на искусственно заселенных его яйцекладками модельных деревьях; ежегодное получение проб интактных корней кормовых растений на постоянных пробных площадях; наблюдения за динамикой таяния снега и прогревания почвы в апреле-мае в березовых насаждениях в Ботаническом саду УрО РАН и в окрестностях г. Екатеринбурга; изучение факторов, определяющих тепловые свойства почвы весной; наблюдения за ростом сосущих корней у березы в Ботаническом саду УрО РАН; ежегодные фенологические наблюдения за кормовыми породами непарного шелкопряда. Количество постоянных пробных площадей составляло в общей сложности около 60. 15-17 из них располагались в березовых насаждениях около ст. Перебор в Каменск-Уральском районе Свердловской области. В 1995-2013 гг. около 10 модельных деревьев в Каменск-Уральском, Белоярском и Сысертском районах на юго-востоке Свердловской области искусственно заселялось яйцекладками непарника.
Метод составления таблиц выживания филлофага на модельных деревьях основан на том, что гусеницы непарного шелкопряда, поднявшись в 1-м возрасте в крону, остаются на модельном дереве до конца развития [3].
Особый раздел методики работы составляли учеты насекомых в очагах массового размножения непарника и в контрольных неочаговых насаждениях. Поскольку многие из видов насекомых, обитающих в березовом лесу, на той или иной стадии своего жизненного цикла встречаются на стволах берез, учеты их можно проводить параллельно с учетами непарного шелкопряда, осматривая нижние части стволов берез. Учеты наиболее важных групп насекомых, связанных с образованием очагов непарного шелкопряда, таких, как пестрянки (Zygaenidae) и нимфалиды (Nymphalidae), проводились также отдельно, на цветущих растениях.
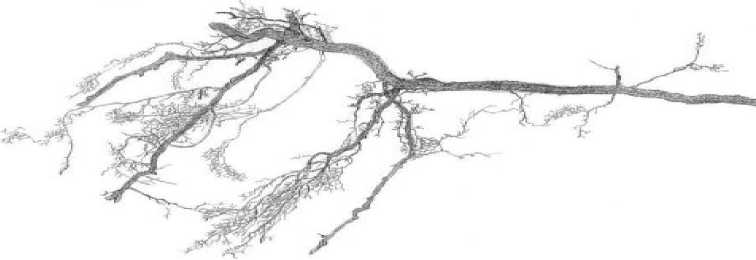
Результаты и их обсуждение. В табл. 1 и 2 представлены таблицы выживания непарного шелкопряда на модельном дереве №1 на постоянной пробной площади №4 около ст. Перебор в 20032004 гг. и 2008-2009 гг. В 2004 г. в березовых насаждениях в окрестностях ст. Перебор возник очаг непарного шелкопряда. Его возникновение проявилось в том, что гусеницы непарника перестали гибнуть в старших возрастах и почти все доживали до стадии куколок (табл. 1). Очевидно, в 2004 г. произошли какие-то изменения в физиологическом состоянии кормового растения, так что его листва стала благоприятной для питания гусениц непарного шелкопряда. Как показали наши наблюдения, изменение физиологического состояния кормового растения при возникновении очага связаны с появлением у него дефицита тонких корней. Тонкими, сосущими корнями называются периодически обновляющиеся корни кормовых растений, выполняющие у деревьев основную работу по поглощению воды и минеральных веществ [2, 12]. В 2005 г. на юго-востоке Свердловской области очаги непарного шелкопряда возникли повсеместно. По нашим данным, «очаговое состояние» насаждений, возникнув, поддерживается чаще всего 4 года [4]. 4 года – это средний срок жизни сосущих корней у древесных пород умеренной климатической зоны Северного полушария [4]. Очаги, возникшие в 2005 г., в 2009 г. должны были прекратить свое существование, так как в 2009 г. у берез должно было восстановиться нормальное количество тонких корней. В 2009 г. повсеместно в Свердловской области выживаемость гусениц непарного шелкопряда понизилась, и его численность упала (табл. 2). Однако в 2009 г. на небольшой площади (около 300 га) в березовых насаждениях между ст. Перебор и с. Покровское очаг непарного шелкопряда возник заново. На рис. 1 показаны образцы интактных корней березы, взятые на постоянной пробной площади №9 в 2008 г. (рис. 1а), 2009 г. (рис. 1б) и в 2012 г. (рис. 1в). В 2006-2008 гг. в этом месте интенсивность очага была невысокой, в 2009 г. она выросла в 3 раза (рис. 1б), а в 2012 г. насаждения вернулись к неочаговому состоянию (рис. 1в).
Таблица 1. Выживаемость непарного шелкопряда на модельном дереве №1 на постоянной пробной площади №4 около ст. Перебор в 2003-2004 гг.
|
Количество особей не-парника на дереве |
Годы |
||
|
2003 |
2004 |
||
|
гусеницы |
1-го возр. |
19 [30-40] |
3 [6-8] |
|
2-го возр. |
19 [30-40] |
2 [6-8] |
|
|
3-го возр. |
14 (дв.) [20-30] |
2 [6-8] |
|
|
4-го возр. |
3 [5] |
4 (1 дв.) [6] |
|
|
5-го возр. |
0 |
6 [4] |
|
|
предкуколки и куколки |
0 |
6 (2дв.) [4] |
|
Таблица 2. Выживаемость непарного шелкопряда на модельном дереве №1 на постоянной пробной площади №4 около ст. Перебор в 2008-2009 гг.
|
Количество особей не-парника на дереве |
Годы |
||
|
2008 |
2009 |
||
|
гусеницы |
1-го возр. |
120 [150] |
50 [60] |
|
2-го возр. |
139 [130] |
56 [60] |
|
|
3-го возр. |
- |
29 [30] |
|
|
4-го возр. |
23 (6дв.) [20] |
17 (4дв., 3Ар.) [20] |
|
|
5-го возр. |
9 (2Ар.) [10] |
3 [6] |
|
|
предкуколки и куколки |
4 (2дв.) [6] |
0 |
|
Примечание: цифры в столбцах таблиц означают число особей непарника, обнаруженных на стволе модельного дерева; цифры в круглых скобках: дв. – число гусениц с яйцами или предкуколок и куколок с личинками двукрылых, Ар. – число гусениц, зараженных Apanteles sp.; цифры в квадратных скобках – приблизительно число особей вредителя в кроне
Каким же образом у берез возникает дефицит тонких корней? Как это ни странно, образование очагов непарного шелкопряда всегда можно связать с определенной датой. Так, очаги филлофа-га между ст. Перебор и с. Первомайское в 2009г. возникли 12 мая. Об этом свидетельствуют данные измерений температуры почвы на постоянных пробных площадях. По нашим наблюдениям, корни берез на юге Свердловской области начинают рост, когда почва прогреется до приблизительно + 60С. 12 мая 2009 г. после потепления, происходило быстрое распускание почек у берез, и температура корнеобитаемого слоя почвы достигла + 60С и выше (табл. 3). Однако в березовых насаждениях между ст. Перебор и с. Покровское у деревьев в это время на глубине 30 см около стволов почва еще не оттаяла, что создавало трудности для водоснабжения, то есть растения испытывали водный стресс в момент перехода к высоким температурам воздуха и резкого ускорения развития листьев. Очевидно, именно этот фактор предопределил возникновение очага массового размножения в 2009 г. около ст. Перебор и с. Покровское. По нашей классификации, данный тип сценария возникновения очага относится к 4-му из 4-х типов погодных сценариев, благоприятствующих началу вспышек массового размножения непарного шелкопряда [6]. Он включает в себя следующую цепь событий. После влажного конца осени в начале зимы выпадает большое количество снега и начинаются морозы. Замерзание почвы происходит от стволов деревьев, поскольку толстый слой снега предохраняет почву от замерзания с поверхности. После превращения жидкой воды в лед в почвенных капиллярах падает давление водяного пара, и вода начинает перемещаться по капиллярам к зоне замерзания. В результате вокруг оснований стволов берез формируются кольца насыщенной льдом почвы, медленно оттаивающей весной вследствие высокой теплоемкости льда (табл. 3). Таким образом, сущность образования очагов массового размножения непарного шелкопряда заключается в следующем. Если в момент быстрого прогревания основного корнеобитаемого слоя почвы весной до +60С и выше у деревьев происходит распускание почек, и они находятся под воздействием водного стресса (табл. 3), то развитие нового поколения сосущих корней нарушается, и в дальнейшем оно остается недоразвитым. В результате у кормовых растений филлофага на время, равное среднему сроку жизни сосущих корней, возникает их недостаток (рис.). Гусеницы чешуекрылого, питающиеся листвой кормового растения с недостатком нитевидных сосущих корней, имеют повышенную выживаемость в старших возрастах (табл. 1), что служит причиной роста численности филлофага.
Одной из загадок динамики популяций непарного шелкопряда является приуроченность его очагов к определенным насаждениям [1]. Например, в березовых насаждениях около ст. Перебор мы наблюдали за возникновением очагов массового размножения филлофага в 1991, 1996, 2000, 2004, 2005, 2009 гг. С 1996 г. по 2012 г. численность непарника здесь непрерывно поддерживалась на повышенном уровне (в среднем около 100 гусениц на дерево). В то же время в березовых насаждениях в долине р. Сысерти в 30 км западнее, служивших контролем, плотность популяции непарного шелкопряда была постоянно низкой. Такая разница объясняется разницей в строении корневых систем деревьев. В окрестностях ст. Перебор березовые насаждения растут на бедных почвах, и основная масса тонких корней у берез сосредоточена в верхнем слое почвы толщиной 6 см, а корни берез в долине р. Сысерти рассредоточены в довольно толстом слое почвы до 30 см. Кроме того, в насаждениях около ст. Перебор хорошо выражена смена поколений тонких корней [8].
Таблица 3. Температура почвы на постоянной пробной площади №9 около ст. Перебор и №2 в окрестностях ст. Марамзино 12 мая 2009 г. на расстоянии 15 и 80 см от ствола модельных деревьев
|
Название учетной площади |
Глубина, на которой измерялась температура почвы |
Температуры почвы на расстоянии от ствола (0С) |
|
|
15 см |
80 см |
||
|
№2 около ст. Марамзино |
10 см |
6,3 |
6,1 |
|
20 см |
5,0 |
4,9 |
|
|
30 см |
3,0 |
+2,8 |
|
|
№3 около ст. Перебор |
10 см |
3,5 |
6,1 |
|
20 см |
1,1 |
5,2 |
|
|
30 см |
0,0 |
+3,6 |
|
Работая в очагах массового размножения непарного шелкопряда, мы обратили внимание на то, что кроме большого количества паразитоидов, хищников и сапрофагов в них неизменно наблюдается повышенная численность нескольких видов пестрянок (Zygaenidae). Позднее в очаге непарного шелкопряда было обнаружено повышение численности также видов из сем. Nymphalidae, гусеницы которых питаются на фиалках. В табл. 4 приведены данные учетов насекомых на стволах берез, сделанные в 2011 г. в очаге непарного шелкопряда и в неочаговых насаждениях. Насекомых, обнаруженных во время учетов, можно разделить на 3 группы: другие виды грызущих филлофагов; парази-тоиды, хищники и сапрофаги; виды бабочек из сем. пестрянок и нимфалид. Численность остальных видов грызущих филлофагов не связана с численностью непарного шелкопряда (исключения из этого правила встречаются, но в других регионах). Повышение в очагах непарника численности пара-зитоидов, сапрофагов и хищников, которые, как правило, также играют роль сапрофагов, ожидаемо. Наиболее, интересным, на наш взгляд, является повышение плотности популяции пестрянок и нимфалид в очагах непарного шелкопряда. Судя по данным наших учетов, оно определяется тем, что в очагах уменьшается густота сосущих корней берез (рис. 1) По-видимому, кормовые растения пестрянок и нимфалид (бобовые и фиалковые) вследствие ослабления конкуренции с корнями березы изменяют свой химический состав и становятся более благоприятными для питания гусениц бабочек данных семейств. По существу, мы имеем дело со вспышками массового размножения, являющимися в некотором смысле зеркальным отражением вспышек численности непарного шелкопряда. Вероятно, повышение численности пестрянок и ним-фалид аналогично подъемам плотности популяций у небольших растительноядных млекопитающих и птиц.
Как это видно из приведенных примеров, борьба с непарным шелкопрядом в 1996-1997 гг. в Тюменской области и в 2011 г. в Челябинской и Курганской областях была не нужна. И подобные ситуации встречаются часто. Мы хотели рассмотреть методы оптимизации борьбы с непарным шелкопрядом, однако статья на данную тему требует привлечения дополнительного материала, что значительно расширяет ее объем. Поэтому мы решили рассмотреть те аспекты динамики численности непарного шелкопряда, которые характеризуют его значение в экосистеме березовых лесов. Для видов, играющих важную роль в функционировании экосистем или поддержании биоразнообразия, было предложено понятие «ключевые виды» [10]. Существует очень мало видов животных, которым можно было бы приписать ключевое значение в наземных экосистемах [10]. На наш взгляд, на роль ключевого вида в березовых насаждениях Урала и Западной Сибири, может претендовать непарный шелкопряд [5].
а
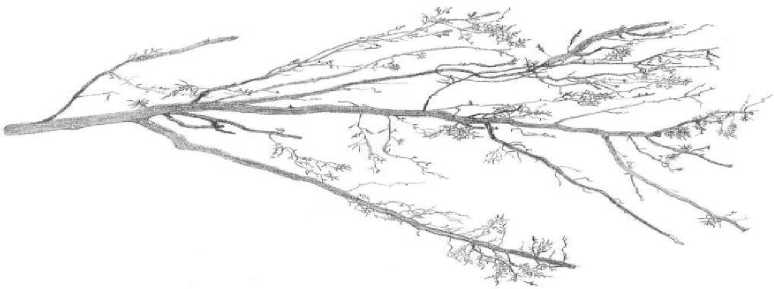
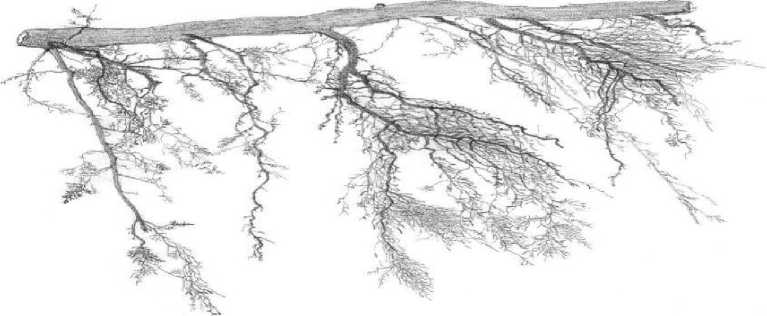
Рис. 1. Образцы интактных корней березы, взятые на постоянной пробной площади № 9 в окрестностях ст. Перебор осенью в 2008 г. (а) и 2009 г. (б) и 2012 г. (в)
Таблица 4. Результаты учетов беспозвоночных на стволах деревьев в очаге непарного шелкопряда около ст. Перебор (пробные площади№3 и №4) и в неочаговых насаждениях, проведенных в июне 2011 г.
|
Названия семейств, родов и видов |
Количество особей на 150 деревьев в очаге непарного шелкопряда |
Количество особей на 150 деревьев в неочаговых насаждениях |
|
Костянки сем. Lithobidae |
5 |
1 |
|
Щелкуны – Elateridae: Selatosomus aeneus L. Prosternon tesselatum Latr. Melanotus rufipes Hbst Elater cinnabarinus Eschz. |
8 |
2 |
|
3 |
2 |
|
|
1 |
- |
|
|
1 |
- |
|
|
Кожееды - Dermestidae |
12 |
- |
|
Мягкотелки-Canthuridae Cantharis rustica Fall. |
1 |
|
|
Малашки – MelyridaeMalachius bipustulatus l. |
1 |
- |
|
Ктыри - Asilidae |
3 |
- |
|
Бекасницы - Rhagionidae |
5 |
- |
|
Тахины – Tachinidae: Parasetigena siilvestris R.D. |
9 |
- |
|
Серые мясные мухи – Sarcophagidae Sarcophaga sp. |
3 |
- |
|
Бабочки пестрянки – Zygaenidae (гусеницы) |
10 |
- |
|
Бабочки нимфалиды – Nymphalidae (гусеницы и куколки) |
3 |
- |
|
Пилильщики – Tenthredinidae (личинки) |
9 |
15 |
|
Клопы щитник и древесные - Acanthosomatidae |
10 |
9 |
Список литературы Непарный шелкопряд как ключевой вид грызущих филлофагов в березовых лесах Западной Сибири
- Воронцов, А.Н. Лесная энтомология. -М.: Высшая школа, 1982. С. 282.
- Колесников, В.А. Методы изучения корневой системы древесных растений. -М.: Лесн. пром-сть, 1972. С. 7.
- Максимов, С.А. Таблицы выживания непарного шелкопряда в неочаговых и очаговых насаждений/С.А. Максимов, В.Н. Марущак, А.Н. Тишечкин//Известия С-Петербургской лесотехнической академии. 2008. Вып. 182. С. 190-200.
- Максимов, С.А. Новый метод определения срока жизни сосущих корней у древесных пород/С.А. Максимов, В.Н. Марущак//Ботанические сады в 21 веке: сохранение биоразнообразия и инновационные решения. -Белгород, 2009. С. 252-257.
- Максимов, С.А. Ключевые виды и биологическое разнообразие в березовых лесах Западной Сибири/С.А. Максимов, В.Н. Марущак//Сохранение биоразнообразия Камчатки и прилегающих морей. Матер. XI конференции, 2010. С. 106-109.
- Максимов, С.А. К теории динамики популяций непарного шелкопряда на Урале/С.А. Максимов, В.Н. Марущак//Аграрный вестник Урала. 2011а. 88 (9). С. 13-16.
- Максимов, С.А. К причинам гибели деревьев в очагах массового размножения непарного шелкопряда/С.А. Максимов, В.Н. Марущак//Экология России: на пути к инновациям. Межвуз. сборн. научн. трудов. 2011б. С. 108-115.
- Максимов, С.А. Мониторинг динамики численности грызущих филлофагов как метод биоиндикации лесных экосистем (на примере непарного шелкопряда)/С.А. Максимов, В.Н. Марущак//Биологический мониторинг природно-техногенных систем. Матер. Всеросс. научно-практ. конф. Ч. 1. -Киров, 2011в. С. 77-81.
- Максимов, С.А. О механизме массовых размножений непарного шелкопряда на Южном Урале/С.А. Максимов, В.Н. Марущак//Фундаментальные и прикладные проблемы науки. Т. 3. Матер. VII Междунар. симпозиума. -М.: РАН, 2012. С. 152-159.
- Bond, W.J. Keystone species/Biodiversity and Ecosystem Function. -Berlin: Sp-Ver., 1994. P. 237-253.
- Elkinton, I.S. Population dynamics of gypsy moth in North America/I.S. Elkinton, A.V. Liebhold//Annu. Rev. Entomol. 1990. V. 35. P. 571-596.
- Thomas, P. Trees: their natural history. -Cambridge: Univer. Press, 2003. P. 78-84.
