Новые данные о видовом разнообразии миксомицетов ЦЛГПБЗ (Нелидовский р-н, Тверская область)
")
Автор: Лебедев Александр Николаевич, Гмошинский Владимир Иванович, Бухтоярова Наталья Юрьевна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Биоразнообразие и охрана природы
Статья в выпуске: 1, 2017 года.
Бесплатный доступ
В мае и октябре 2014 года была проведена работа по изучению видового разнообразия миксомицетов южного лесничества Центрально-Лесного государственного природного биосферного заповедника. Был собран 621 образец, относящийся к 70 видам из 25 родов, 10 семейств и 5 порядков. При этом 45 видов были новыми для территории ЦЛГПБЗ, а 22 были новыми для Тверской области.
Миксомицеты, биота, биоразнообразие, заповедник, тверская область
Короткий адрес: https://sciup.org/146116696
IDR: 146116696 | УДК: 582.241(470.331)
New data on myxomycetes diversity in Central forest nature reserve (Nelidovo area, Tver region)
The check list of myxomycetes collected in May and October of 2014 on the territory of Central Forest Nature Reserve is presented. 621 samples have been investigated. 70 species from, 25 genera, 10 families and 5 orders have been described. 45 species were new to the territory of the Reserve, and 22 were new for the Tver Region.
Текст научной статьи Новые данные о видовом разнообразии миксомицетов ЦЛГПБЗ (Нелидовский р-н, Тверская область)
Миксомицеты — своеобразная группа почвенных амеб, насчитывающая около 962 видов (Lado, 2005-2015). Вегетативная стадия миксомицетов представлена гаплоидными миксамебами и зооспорами, а также обычно диплоидными плазмодиями. Покоящиеся стадии могут быть представлены микро- и макроцистами, а генеративная стдаия - спорофорами.
Центрально-Лесной государственный природный биосферный заповедник (ЦЛГПБЗ) находится на западе Тверской области, в Нелидовском районе. ЦЛГПБЗ расположен на западе Европейской части России, на главном Каспийско-Балтийском водоразделе Русской равнины (непосредственно на водоразделе верховьев р. Волга и р. Западная Двина), в юго-западной оконечности Валдайской возвышенности. Территория заповедника представляет собой слабо всхолмленную равнину. Абсолютные отметки высот колеблются от 220 до 280 м над уровнем моря.
В заповеднике сохранилось естественное разнообразие почв южной тайги. ЦЛГБПЗ расположен в переходной полосе между западноевропейским и восточноевропейским районами континентальной области умеренного климата. В этой зоне преобладают воздушные потоки атлантического происхождения, но часты вторжения с севера холодных арктических воздушных масс. За год выпадает в среднем 700 мм осадков, причем большая часть - в теплый период года.
Природный комплекс заповедника типичен для южно-таежной подзоны и является эталоном первичного биогеоценотического покрова обширной области моренного рельефа центральной части Русской равнины. Здесь сохранился уникальный, исторически сложившийся комплекс южнотаежных ельников, не затронутых рубками. В структуре растительного покрова он занимает доминирующее положение (47% площади). Около 40% лесного массива занимают березняки и осинники, возникшие в результате сукцессионных процессов (вывалов, пожаров). Сосняки произрастают на 10% площади заповедника и представлены заболоченными сообществами с низко продуктивными древостоями. Черноольховые леса (1-2%), расположены в долинах рек и ручьев. Около 6% площади заповедника занимают верховые сфагновые болота. Для лесного массива заповедника характерны большие площади ветровальных участков, вызванные ураганами.
На территории ЦЛГПБЗ изучение видового разнообразия миксомицетов проводилось только в конце 70-х годов XX века Юрием Капитоновичем Новожиловым (Новожилов, 1980). К началу наших исследований на данной территории сообщалось об обнаружении 37 видов из 5 порядков: Stemonitales, Liceales, Trichiales, Physarales и Ceratiomyxales.
Материал и методика. Сбор спороношений производили со 2 по 4 мая и с б по 8 октября 2014 года на территории 11 площадок: 1) Юг 96 квартала (56°27.243' с.ш., 032°59.385' в.д.). 2) Северо-восток 96 квартала (56°28.338' с.ш.; 032°59.548' в.д.). 3) Северо-восток 95 квартала (5б°28.64Г с.ш.; 032°58.762' в.д.). 4) Запад 94 квартала (56°28.035' с.ш.; 032°57.393' в.д.). 5) Север 78 квартала (56°30.170' с.ш.; 032°56.334' в.д.). 6) Север 80 квартала (56°30.208' с.ш.; 032°57.943' в.д.). 7) Юг 94 квартала (56°27.380' с.ш.; 032°57.617' в.д.). 8) Северо-восток 106 квартала (56°27.566' с.ш.; 032°55.924'). 9) Запад 91 квартала (56°27.303' с.ш.; 032°53.552' в.д.). 10) Просека 92 и 93 кварталов (56°28.265' с.ш.; 032°54.88Г в.д.). 11) Лес у северной оконечности пос. Заповедный (5б°27.679' с.ш.; 032°57.923' в.д.). Кроме того, 28 образцов обнаружено вне соответствующих биотопов. Для таких образцов приводится соответственное описание места сбора. Спороношения срезали ножом вместе с фрагментом субстрата и помещали в спичечный коробок, снабжая соответствующей этикеткой включающей в себя информацию о месте, дате сбора и типе субстрата. Образцы высушивали в теплом, сухом месте вдали от прямых солнечных лучей. Определенные образцы были переданы в коллекцию кафедры микологии и альгологии, где им были присвоены соответствующие инвентарные номера.
Определение видовой принадлежности производили с использованием световых микроскопов ScienOP ВР-52, бинокуляров Scien OP-SP30-C, а также сканирующего электронного микроскопа.
Аннотированный список видов
Общая схема описания каждого вида в аннотированном списке видов выглядит следующим образом:
Первая строка аннотированного списка видов содержит латинское наименование и полную ссылку на авторов данного таксона. Вторая строка содержит информацию об образце. Знак * обозначает, что вид впервые отмечен для территории ЦЛГПБЗ. Знак # свидетельствует о том, что вид является новым для Тверской области.
Вторая строка содержит информацию о числе собранных образцов, их субстратной приуроченности, о локалитетатах, в которых обнаружен данный вид, а также номера гербарных образцов. Цифра в начале соответствует числу обнаруженных образцов в ходе всех сборов (май и октябрь). Через знак «-» приводится общая частота встречаемости данного вида по результатам сборов в мае и октябре совместно. Далее в аналогичном формате дана информация о числе образцов и частоте встречаемости этого вида в каждый из месяцев. Следующее предложение содержит информацию о числе образцов обнаруженных на соответствующих субстратах: W (wood) - древесина, В (bark) - кора, L (litter) - листовой опад и F (fungi) - плодовые тела грибов. После знака «-» приводятся данные о числе образцов. Последнее предложение содержит информацию о местах обнаружения спороношений данного вида: цифра приведенная без скобок обозначает номер биотопа; значения в круглых скобках соответствуют числу собранных образцов в данном биотопе; в квадратных скобках приводятся присвоенные инвентарные номера в коллекции миксомицетов кафедры микологии и альгологии Биологического факультета Московского Государственного университета имени М.В. Ломоносова.
^Arcyodes incarnata (Alb. et Schwein.) O.F. Cook, Science 15:651 (1902). '
-
2 - 0,32 %: май - 0 (0%), окт. - 2 (0,6 %). W - 1, В - 1. Loc. 3 (1) [5335], 6(1) [5405].
"^Arcyria affinis Rostaf., Sluzowce monogr. 276 (1875).
-
18 - 2,87 %: май - 0 (0%), окт. - 18 (5,44 %). W - 15, В - 3. Loc. 1 (1) [5290], 2 (1) [5309], 3 (1) [5328], 4 (3) [5349], 5 (1) [5372], 6 (2) [5392], 8 (3) [5481], 9 (3) [5465], 11 (3) [5475].
Arcyria cinerea (Bull.) Pers., Syn. meth. fung. 1:184 (1801).
7-1,11 %: май - 0 (0%), окт. - 7 (2,11 %). W - 5, В - 1, L - 1. Loc. 3 (1) [5338], 5 (1) [5365], 6 (2) [5398], 7 (1) [5509], 8 (1) [5492], 9 (1) [5474].
Arcyria denudata (L.) Wettst., Verh. Zool.-Bot. Ges. Wien 35:535 (1886).
-
8 - 1,27 %: май - 3 (1,0 %), окт. - 5 (1,51 %). W - 7, В - 1. Loc. 1 (1) [5102], 2 (2) [5307], 6 (3) [5393], 10 (1) [4932], север 104 квартала (N 56°27.423' E 032°53.748') (1) [4991].
Arcyria incarnata (Pers, ex J.F. Gmel.) Pers., Observ. mycol. 1:58 (1796).
-
2 - 0,32 %: май - 0 (0%), окт. - 2 (0,6 %). W - 1, В - 1. Loc. 3 (1) [5337], 6(1) [5397].
*Arcyria cf. insignis Kalchbr. et Cooke, in Kalchbrenner, Grevillea 10:143 (1882). ’ ‘ '
Спорангии не полностью развитые, в результате чего невозможно установить, насколько плотно капиллиций связан с основанием чашечки спорангия. При этом микроскопические признаки полностью соответствуют типичным для A. insignis; капиллиций очень эластичный: сильно вытягивается при созревании спорангия, орнаментирован недостаточно хорошо выраженными полукольцами со скругленными верхушками; споры 7-8 мкм в диам., практически гладкие.
#Arcyria magna Rex, Proc. Acad. Nat. Sci. Philadelphia 45:364 (1893).
1 - 0,16 %: май - 0 (0%), окт. - 1 (0,3 %). В - l.Loc. 3 (1) [5344].
Arcyria obvelata (Oeder) Onsberg, Mycologia 70(6): 1286 (1979).
-
3 - 0,48 %: май - 0 (0%), окт. - 3 (0,91 %). W - 3. Loc. 3 (1) [5336], 4 (1) [5348], 5 (1) [5370].
#Arcyria occidentalis (T. Macbr.) G. Lister, in Lister, Monogr. mycetozoa, ed. 2 245 (1911). '
1-0,16 %: май - 0 (0%), окт. - 1 (0,3 %). W - l.Loc. 1 (1) [5289].
^Arcyria oerstedii Rostaf., Sluzowce monogr. 278 (1875).
-
2 - 0,32 %: май - 0 (0%), окт. - 2 (0,6 %). W - 2. Loc. 1 (1) [5301], 3 (1) [5345].
^Arcyriapomiformis (Leers) Rostaf., Sluzowce monogr. 271 (1875).
-
3 - 0,48 %: май - 1 (0,3 %), окт. - 2 ( 0,6 %). W - 3. Loc. 2 (1) [5326], 4 (1) [4909], 6 (1).
#Arcyria stipata (Schwein.) Lister, Monogr. mycetozoa, ed. 1 189 (1894).
-
4 - 0,64 %: май - 0 (0%), окт. - 4 (1,21 %). W - 4. Loc. 4 (1) [5351], 8 (1) [5498], 9 (2) [5466].
Arcyria sp.
Спорангии серовато-желтоватых тонов, но без зеленоватого оттенка. Сеть капиллиция слабо связана с основанием достаточно уплощенной чашечки, орнаментированной радиально расходящимися ребрами. Капиллиций орнаментирован полукольцевидными утолщениями. Споры практически гладкие, около 8 мкм в диам. Возможно, это сильно поврежденные спорангии Л. obvelata, однако для этого вида имеется не типичная орнаментация нитей капиллиция. У A. obvelata капиллиций орнаментирован полукольцевидными утолщениями с острыми верхушками, в результате чего они выглядят подобно крупным гребневидным образованиям или шипам. Однако в данном случае подобной орнаментации не наблюдается. Тускло-желтая окраска спороношений могла возникнуть вследствие выцветания как красных спорофоров, так и желтых. Для установления видовой принадлежности требуются дополнительные исследования.
1-0,16 %: май - 0 (0%), окт. - 1 (0,3 %). W - 1. Loe. 3 (1) [5333].
#Barbeyella minutissima Meyl., Bull. Soc. Bot. Geneve 6:89 (1914).
-
5 - 0,80 %: май -1 (0,3 %), окт. - 4 (1,21 %). W - 5. Loc. 5 (2) [5376, 5377], 6 (1) [4896], 8 (1) [5500], 9 (1) [5457]. Рис. 1.
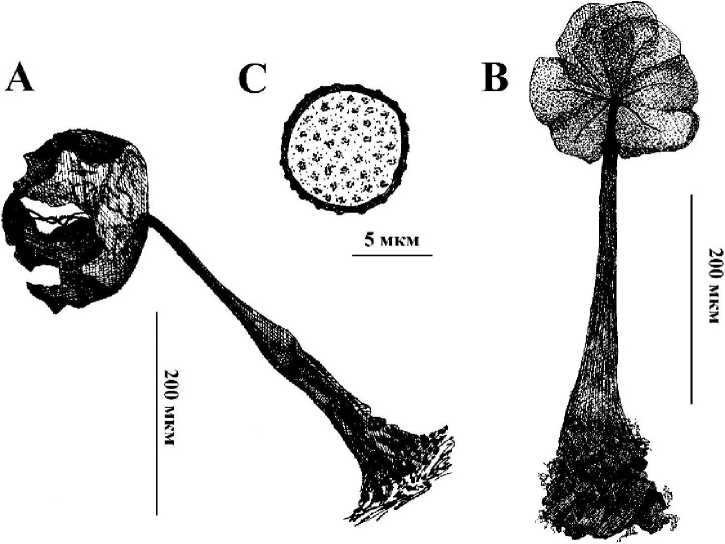
Рис. 1 . A - Внешний вид спороношений В. minutissima. В - Строение колонки, капиллиция и перидия В. minutissima в проходящем свете. С - Споры В. minutissima. Примечания: Этот редкий вид на территории России отмечен только на Урале (Башкирия и Свердловская область) (Новожилов, 2005). По состоянию на 2000 год в мире было известно всего 28 находок (Schnittler et al., 2000а). К сожалению, данные об экологии этого вида весьма недостаточны, однако нахождение его на территории ЦЛГПБЗ представляется чрезвычайно интересным. В Европе В. minutissima встречается обычно в осенние месяцы в горах на высоте от 500 до 1300 метров. В Северной Америке образцы этого вида были найдены в период с марта по сентябрь на высотах 2000-2300 м над уровнем моря. Известны еще два образца из Калифорнии, однако они были найдены в январе. Образцы из Мексики были отмечены на высотах от 3000 до 3500 м. Однако, этот вид нельзя полностью относить к экологической группе альпийских видов, поскольку известно три образца
(два из Финляндии и один из Польши), обнаруженных на небольшой высоте над уровнем моря в зоне умеренного климата (Schnittler et al., 2000а). Однако все эти образцы, были найдены в осенние месяцы: оба образца из Финляндии найдены в ноябре, а образец из Польши, предположительно, в августе или сентябре (Schnittler et al., 2000а). Таким образом, обнаружение спороношений в весенние месяцы в зоне умеренного климата является не типичным для В. minutissima. Обычно В. minutissima обнаруживают в очень влажных местах совместно с мхами, печеночниками и водорослями. Спорокарпы обычно образуются на поверхности мхов, но никогда на поверхности печеночников (Schnitller et al., 2000а). Предполагается, что протоплазмодии этого вида способны питаться полисахаридами водорослей, а при формировании спороношений предпочитают выползать на более сухое место, в результате чего они никогда не развиваются в окружении водорослевого чехла, как, к примеру, Colloderma oculatum (С. Lippert) G. Lister. Подробное описание морфологии спороношений этого вида приводится в работе М.Шниттлера с соавторами (Schnittler et al., 2000b).
#Calomyxa metallica (Berk.) Nieuwl., Amer. Midi. Naturalist 4:335 (1916). '
-
3 - 0,48 %: май - 0 (0 %), окт. - 3 (0,91 %).W - 1, В - 2. Loc. 6 (2) [5382], 9(1) [5454].
*Clastoderma debaryanum X. Blytt, Bot. Zeitung (Berlin) 38:343 (1880).
-
2 - 0,32 %: май - 0 (0 %), окт. - 2 (0,6 %). W - 2. Loc. 1 (1) [5283], 5 (1).
Collaria arcyrionema (Rostaf.) Nann.-Bremek. ex Lado, Ruizia 9:26 (1991). ‘
-
11 - 1,75 %: май - 4 (1,4 %), окт. - 7 (2,11 %). W - 11. Loc. 1 (1) [4956], 2 (1) [5321], 5 (3) [4976, 4979, 5375], 6 (3), 8 (1) [5484], 9 (2) [5075, 5455].
Comatricha elegans (Racib.) G. Lister, in Lister, Guide Brit, mycetozoa, ed. 3 31 (1909).
1-0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). W - \.Loc. 2 (1) [5308]. ^Comatricha ellae Hark., Karstenia 18(1):23 (1978).
-
2 - 0,32 %: май - 0 (0 %), окт. - 2 (0,6 %). W - 2. Loc. 2 (1) [5316], 3 (1) [5340].
^Comatricha nigra (Pers, ex J.F. Gmel.) J. Schrot., in Cohn, Krypt.-Fl. Schlesien 3(1):118 (1885) ["1889"]. "
-
9 - 1,43 %: май - 3 (1,0 %), окт. - 6 (1,81 %). W - 7, В - 2. Loc. 2 (1) [5097], 3 (4) [5086, 5339], 6 (1) [5401], 7 (1), 8 (1), 9 (1) [5463].
#Comatricha pulchella (C. Bab.) Rostaf., Sluzowce monogr. suppl. 27 (1876).
-
4 - 0,64 %: май - 1 (0,3 %), окт. - 3 (0,91 %). W - 4. Loc. 1 (1) [5304], 3 (1) [5081, 5378], 5 (1), 9 (1) [5486].
^Cribraria aurantiaca Schrad., Nov. gen. pl. 5 (1797).
1 - 0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). W - l.Loc. 7 (1) [5511].
Cribraria cancellata (Batsch) Nann.-Bremek., Nederlandse Myxomyceten (Zutphen) 92 (1975). ' '
-
5 - 0,8 %: май - 1 (0,3 %), окт. - 4 (1,21 %). W - 5. Loc. 4 (1) [5359], 5 (1) [5363], 6 (1) [5400], 8 (1) [5497], 9 (1) [5069].
#Cribraria languescens Rex, Proc. Acad. Nat. Sci. Philadelphia 43:394 (1891).
1-0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). W - 1. Loc. 7 (1) [5504].
#Cribraria minutissima Schweim, Trans. Amer. Philos. Soc., new ser. 4:260(1832).
-
1 - 0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). В - l.Loc. 9 (1) [5461].
*Cribraria purpurea Schrad., Nov. gen. pl. 8 (1797).
-
10 - 1,59 %: май - 5 (1,7 %), окт. - 5 (1,51 %). В - 10. Loc. 2 (1) [5095], 6 (3) [5388], 7 (4) [4964, 5512], 8 (2) [4917].
*Cribraria rufa (Roth) Rostaf., Sluzowce monogr. 232 (1875).
-
10 - 1,59 %: май - 7 (2,3 %), окт. - 3 (0,91%). W - 10. Loc. 5 (2) [4980, 4977], 6 (2) [5395], 8 (4) [4919], 9 (1) [5070], юг 65 квартала (N 46°35.203'; E 032 °56.954') (1) [5407].
ttCribraria tenella Schrad., Nov. gen. pl. 6 (1797).
-
1 - 0,16 %: май - 1 (0,3 %), окт. - 0 (0 %). W - 1. Loc. 6 (1) [4900].
*Cribraria vulgaris Schrad., Nov. gen. pl. 6 (1797).
-
2 - 0,32 %: май - 2 (0,7 %), окт. - 0 (0 %). W - 2. Loc. 6 (2) [4891].
Cribraria sp.
Образец относится к роду Cribraria. Сеточка перидия представлена 20-30 ребрами, отходящими от основания спорангия. Спорангии желтоватого цвета. Чашечка отсутствует. От С. cancellata отличается отсутствием темных тонов в окраске спорангия и фиолетовых диктидиновых гранул. По литературным данным единственный вид, под описание морфологии спороношения которого подходит данный образец - Cribraria rut На, которая является чрезвычайно редкой и была отмечена только в Австралии (Martin, Alexopoulos, 1969). При этом определение осложняется тем, что спорангии повреждены грибами и с не до конца сформированными спорами, так что узнать их диаметр, окончательную окраску и форму не представляется возможным.
1 -0,16 %: май - 1 (0,3 %), окт. - 0 (0 %). W- l.Loc. 7 (1) [4959].
Diacheopsis sp.
Спорофоры - сидячие шаровидные спорангии, покрытые пленчатым перидием с радужным оттенком. Ножка отсутствует. Колонка отсутствует. Гипоталлус развит слабо. Капиллиций в виде отдельных светло-коричневых нитей, с небольшими темными шаровидными утолщениями, отходящих от основания спорангия, сильно ветвящихся и анастомозирующих. Споры шаровидные, 12 мкм в диам., с небольшим светлым пятнышком, равномерно орнаментированные хорошо заметными бородавочками. Спорангии сформировались на листьях мха. Среди видов рода Diacheopsis есть только 4 вида с шаровидными и отдельно стоящими спорангиями: D. nannengae G. Moreno, Iliana et Heykoop, D. mitchellii Nann.-Bremek. et Y. Yamam., D. minuta Nann.-Bremek. et Y. Yamam., D. insessa (G. Lister) Ing. Образец 5334 отличается от D. nannengae наличием темных утолщений в структурах капиллиция и более крупными спорами. D. mitchellii обладает очень крупными спорами (до 20-21 мкм в диам.), у D. minuta споры орнаментированны бородавочками собранными в группы, а у D. insessa споры также более крупные, 18-19 мкм. Для установления видовой принадлежности требуются дополнительные исследования.
' 1-0,16 %: май - 0 (0%), окт. - 1 (0,3 %). W - 1. Loc. 3 (1) [5334].
#Diderma floriforme (Bull.) Pers., Neues Mag. Bot. 1:89 (1794).
1 - 0,16 %: май - 1 (0,3 %), окт. - 0 (0 %). W - l.Loc. 9 (1) [5071].
#Diderma radiatum (L.) Morgan, J. Cincinnati Soc. Nat.Hist. 16(4): 151 (1894). "
-
24 - 3,82 %: май - 14 (4,7 %), окт. - 10 (3,02%). W - 22, В - 2. Loc. 2 (4) [5096, 5090, 5317], 4 (2) [5358, 5360], 5 (3) [4974], 6 (3) [5389], 7 (2) [4960, 5480], 8 (5) [4913, 5490], 9 (1) [5469], 10 (2) [4938], 93 квартал (N 56°28.784', E 032°56.306') (1) [4944], юг 81 квартала (N 56°28.89Г E 032°58.676') (1) [4993].
Diderma sp.l
Образец соответствует роду Diderma - спорангии на ножках, шаровидные, белые в верхней части, с черным дисковидным основанием. Перидий двухслойный: наружный слой белый, скорлуповидный, обызвествленный, ломкий; внутренний - пленчатый, покрытый известью, серый. Растрескивание происходит продольной щелью. Известь в структурах перидия гранулированная. Ножка достаточно длинная, достигает половины высоты спороношения, морщинистая, желтовато-охристая, необызвествленная. Колонка присутствует, небольшая, черная. Капиллиций представлен отдельными тонкими нитями, слабо ветвящимися и не образующими сеть, фиолетово-коричневыми, практически без утолщений. Споры черные в массе, темно-фиолетово-коричневые в проходящем свете, около 9 мкм в диам., орнаментированные мелкими бородавочками. Оболочка спор равномерно утолщена. Из всех видов рода Diderma наиболее близкими по цвету спороношения, наличию двухслойного перидия и спорангиев на ножках можно считать D.montanum (Meyl.) Meyl., D. umbilicatum Pers, и D. carneum Nann.-Bremek. Однако все эти виды обладают хорошо развитой и сильно обызвествленной колонкой. Ни в одном описании приведенных выше видов не упомянуто о возможности наличия у них темного основания спорангия, а также удлиненной, сравнительно тонкой и необызвествленной ножки.
-
1 - 0,16 %: май - 1 (0,3 %), окт. - 0 (0 %). W - 1. Loc. 2 (1) [5092].
Diderma sp.2
По внешнему облику образцы напоминают виды рода Diderma. Спорофоры - спорангии на ножках, рассеянные или собранные в группы по 2-3, темно-коричневые, шоколадно-коричневые. Перидий однослойный, темно-коричневый, пленчатый, светло-коричневый - в проходящем свете. Растрескивание спорангия происходит за счет апикальной щели. Гипоталлус хорошо развит, темно-коричневый, пленчатый. Ножка красноватокоричневая, пленчатая, необызвествленная. Капиллиций представлен тонкими, практически гладкими нитями 1-2 мкм в диам., с небольшим количеством темных шаровидных утолщений. Споры шаровидные или почти шаровидные, черные в массе, фиолетово-коричневые в проходящем свете 1113 мкм в диам., орнаментированные мелкими, но хорошо заметными шипиками или бородавочками, с равномерно утолщенной оболочкой. В спорофорах, которые были использованы при приготовлении препарата, не было обнаружено хорошо развитой колонки. На основании использования современной определительной литературы (Nannenga-Bremekamp, 1991; Poulain et al., 2011), можно сделать вывод, что образец наиболее близок к D. rufum Nann.-Bremek., однако споры несколько меньшего диаметра и отсутствует хорошо выраженная колонка. Кроме того, относительная длина ножки у наших образцов несколько меньше, чем в описании D. rufum, где сказано, что ножка должна составлять половину или больше от общей высоты спороношения (Nannenga-Bremekamp, 1991).
-
2 - 0,32 %: май - 2 (0,6 %), окт. - 0 (0 %). W - 2. Loc. 6 (1) [4899], 8 (1) [4921].
Didymium melanospermum (Pers.) T. Macbr., N. Amer. Slime-moulds, ed. 1 88 (1899).
-
5 - 0,80 %: май - 0 (0 %), окт. - 5 (1,51 %). W - 2, L - 3. Loc. 6 (3) [5394], 9 (1) [5460], южная оконечность 65 квартала (N46°35.203' E 032°56.954') (1) [5406].
^Didymium minus (Lister) Morgan, J. Cincinnati Soc. Nat.Hist. 16(4): 145 (1894). * "
1-0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). W - l.Loc. 1 (1) [5293].
^Enerthenema papillatum (Pers.) Rostaf., Sluzowce monogr. suppl. 28 (1876).
-
1 - 0,16%: май - 0 (0 %), окт. - 1 (0,3 %). W - l.Loc. 9 (1) [5459].
#Fuligo leviderma H. Neubert, Nowotny et K. Baumann, Myxomyceten 2:211 (1995). " " '
-
2 - 0,32 %: май - 1 (0,3 %), окт. - 1 (0,3 %). W - 2. Loc. 2 (1) [5314], 5 (1) [4984].
#Fuligo luteonitens L.G. Krieglst. et Nowotny, in Neubert, Nowotny et Baumann, Myxomyceten 2:213 (1995).
-
3 - 0,48 %: май - 3 (1,0 %), окт. - 0 (0 %). W - 3. Loc. Юго-запад 92 квартала (N 56°27.619', E 032°58.136') (3) [4942, 4943, 4989].
Fuligo septica (L.) F.H. Wigg., Prim. fl. holsat. 112 (1780).
-
2 - 0,32 %: май - 1 (0,3 %), окт. - 1 (0,3 %). W - 2. Loc. 4 (1) [5354], 10 (1) [4933].
HHemitrichia calyculata (Speg.) M.L. Farr, Mycologia 66(5):887 (1974).
-
6 - 0,96 %: май - 5 (1,7 %), окт. - 1 (0,3 %). W - 6. Loc. 1 (1) [4952], 6 (1) [5402], 8 (1) [4916], 10 (2) [4934], Юго-запад 92 квартала (N 56°27.619', E 032°58.136')(l) [4945].
Hemitrichia clavata (Pers.) Rostaf., in Fuckel, Jahrb. Nassauischen Vereins Naturk. 27-28:75 (1873).
-
15 - 2,39 %: май - 9 (3,0 %), окт. - 6 (1,81%). W - 14, В - 1. Loc. 1 (2) [5292], 3 (2) [5080, 5343], 4 (2) [4908, 5355], 6 (1) [5384], 7 (3) [4970, 5508], 10 (4) [4935], 95 квартал (N 56°27'197" E 032°58'854") (1) [4940].
^Hemitrichia serpula (Scop.) Rostaf. ex Lister, Monogr. mycetozoa, ed. 1 179(1894). " '
-
20 - 3,18%: май - 18 (6,0 %), окт. - 2 (0,6 %). W - 16, В - 1, L - 3. Loc. 1 (5) [4954], 2 (1) [5094], 3 (1) [5088], 5 (2) [5373], 7 (1) [4968], 8 (3) [5104, 4914], 10 (6) [4936], 95 квартал (N 56°27.197' Е 032°58.854') (1) [4947].
*Lamproderma arcyrioides (Sommerf.) Rostaf, Sluzowce monogr. 206 (1874). *
1-0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). W - 1. Loc. 6 (1) [5381].
*Lamproderma columbinum (Pers.) Rostaf., in Fuckel, Jahrb. Nassauischen Vereins Naturk. 27-28:69 (1873).
-
5 - 0,80 %: май - 2 (0,7 %), окт. - 3 (0,91 %).W - 5. Loc. 1 (1) [5294], 2 (1) [5319], 8 (3) [4922, 5103, 5487].
-
3 - 0,48 %: май - 3 (1,0 %), окт. - 0 (0 %). W - 3. Loc. 2 (2) [5093], 3 (1) [5099].
Leocarpus fragilis (Dicks.) Rostaf., Sluzowce monogr. 132 (1874).
1 - 0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). W/L - 1. Loc. 2 (1) [5311].
*Lepidoderma tigrinum (Schrad.) Rostaf, in Fuckel, Jahrb. Nassauischen Vereins Naturk. 27-28:73 (1873).
-
2 - 0,32 %: май - 1 (0,3 %), окт. - 1 (0,3 %).W- 1, В - 1. Loc. 9 (2) [5076, 5456].
#Licea minima Fr., Syst. mycol. 3:199 (1829).
1-0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). W - 1. Loc. 6 (1) [5404].
#Liceapygmaea (Meyl.) Ing, Trans. Brit. Mycol. Soc. 78(3):443 (1982).
1-0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). W - 1. Loc. 1 (1) [5297].
#Licea testudinacea Nann.-Bremek., Acta Bot. Neerl. 14:141 (1965).
2 - 0,96 %: май - 0 (0 %), окт. - 2 (0,6 %). W - 2. Loc. 2 (2) [5300]. Lycogala epidendrum (L.) Fr., Syst. mycol. 3:80 (1829).
-
28 - 4,44 %: май - 9 (3,0 %), окт. - 19 (5,72%). W- 24, В - 4. Loc. 1 (5) [4957, 4951, 5291], 2 (3) [5318], 3 (6) [5083, 5347], 4 (3) [4907, 5353], 6 (1) [4902], 7 (1) [4966], 8 (2) [5482], 9 (1) [5464], 10 (2) [4931, 4937], 11 (3) [5476], 95 квартал (N 56°28.919', E 032°56.124') (1) [4948].
Lycogala exiguum Morgan, J. Cincinnati Soc. Nat.Hist. 15(3-4): 134 (1893). ‘ ~
1-0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). W - 1. Loc. 5 (1) [5379].
^Metatrichia floriformis (Schwein.) Nann.-Bremek., Proc. Kon. Ned. Akad. Wetensch., C. 88(1): 127 (1985).
-
13 - 2,07 %: май - 8 (2,7 %), окт. - 5 (1,51 %). W - 13. Loc. 1 (1) [5323], 2 (2) [5322], 6 (5) [4894, 5403], 7 (5) [4971, 5506].
Metatrichia vesparia (Batsch) Nann.-Bremek. ex G.W. Martin & Alexop., Myxomycetes 143 (1969).
" 35 - 5,57 %: май - 26 (8,7 %), окт. - 9 (2,71%). W - 30, В - 5. Loc. 1 (8)
[4953, 5284, 5288], 2 (4) [5077, 5324], 3 (1) [5084], 4 (2) [4905], 5 (4) [4978, 5380], 6 (2) [4893, 5386], 7 (1) [4965], 8 (3) [4923], 9 (4) [5066], 10 (3) [4930], 11 (1) [5479], юго-запад 92 квартала (N 56°27.619', E 032°58.136') (1) [4941], (N 56°27.182', E 032°58.895') (1) [4988].
*Mucilago Crustacea F.H. Wigg., Prim. fl. holsat. 112 (1780).
-
2 - 0,32 %: май - 0 (0 %), окт. - 2 (0,6 %). W - 2. Loc. 5 (2) [5366].
Physarum album (Bull.) ChevalL, Fl. gen. env. Paris 1:336 (1826).
-
34 - 5,41 %: май - 0 (0 %), окт. - 34 (10,24 %). W- 28, В - 6. Loc. 1 (6) [5295, 5302], 2 (5) [5306], 3 (2) [5341], 4 (4) [5357], 5 (4) [5357], 6 (5) [5399], 7 (1) [5503], 8 (3) [5486, 5489], 9 (4) [5462].
^Physarum leucopus Link, Ges. Naturf. Freunde Berlin Mag. Neuesten Entdeck. Gesammten Naturk. 3:27 (1809).
1 - 0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). В - l.Loc. 2 (1) [5315].
“Physarum psittacinum Ditmar, in Sturm, Deutschl. FL, Pilze 1(4): 125 (1817). *
-
2 - 0,32 %: май - 0 (0 %), окт. - 2 (0,6 %). В - 2. Loc. 11 (2) [5477].
#Physarum pulcherrimum Berk. & Ravenel, in Berkeley, Grevillea 2:65 (1873). ' "
-
3 - 0,48 %: май - 0 (0 %), окт. - 3 (0,9 %). W - 2 + W/F - 1. Loc. 5 (2) [5368, 5369], 9 (1) [5470].
“Physarum viride (Bull.) Pers., Ann. Bot. (Usteri) 15:6 (1795).
-
3 - 0,48 %: май - 0 (0 %), окт. - 3 (0,9 %). W - 3. Loc. 2 (1) [5312], 5 (1), 8 (1) [5495].
Stemonaria sp.
Образец, скорее всего, относится к роду Stemonaria, т.к. ножка не волокнистая при основании и капиллиций рыхлый. Спорангии темные, практически черные, на ножках, собранные в группы, но без потери индивидуальности. Капиллиций относительно рыхлый, нити капиллиция ветвятся 3-4 раза, прежде чем достигают периферии спорангия. Отходят от колонки довольно часто под прямым углом, с расширением в нижней части. Во внутренней и центральной части сети капиллиций расширен, темнокоричневый, каштаново-коричневый. На периферии спорангия нити утончаются, но при этом не волосовидные, светлые, бледно-коричневатые. Споры черные, насыщенно-темно-коричневые в массе, коричневые в проходящем свете, с неравномерно утолщенной оболочкой, шаровидные или почти шаровидные, 8-9 мкм в диам., орнаментированные мелкими бородавочками. В роде Stemonaria всего 5 видов с бородавчатыми спорами: S. laxa Nann.-Bremek. et Y. Yamam., S. irregularis (Rex) Nann.-Bremek., R. Sharma et Y. Yamam, S. nannengae (T.N. Lakh, et K.G. Mukerji) Nann.-Bremek., R. Sharma et Y. Yamam., S. pallidofila Y. Yamam. et Nann.-Bremek. и S. laxiretis Nann.-Bremek. et Y. Yamam. S. irregularis обладает чрезвычайно рыхлой сетью капиллиция с грубыми утолщениями. У S. laxa капиллиций отходит от колонки под произвольными углами и не имеет расширений при оснований. S. pallidofila обладает более темной окраской и спорами большего диаметра. S. laxiretis обладает очень открытой сетью капиллиция с большим количеством мембраноподобных узелков. S. nannengae подходит по диаметру спор и по наличию светлых окончаний капиллиция, более или менее подходит под неполное описание из Poulain et al., 2011. Однако в той же работе упомянуто, что бородавочки на поверхности спор собраны в линии. При этом, имеющийся у нас образец не соответствует описанию в работе А.В.Власенко и Ю.К.Новожилова (2010): от цвета спорангиев и спор до длины ножки. Более того, имеющийся у нас образец не соответствует рисунку на стр. 597 в работе Ю. Ямомото (Yamamoto, 1998), на котором изображены споры с равномерно утолщенной оболочкой.
1 - 0,16 %: май - 1 (0,3 %), окт. - 0 (0 %). W - 1. Loc. 6 (1) [4898].
Stemonitis axifera (Bull.) T. Macbr., N. Amer. Slime-moulds, ed. 1 120 (1899). '
-
11 - 1,75 %: май - 4 (1,4 %), окт. - 7 (2,11 %). W - 11. Loc. 1 (1) [5303], 2 (2) [5313], 3 (1) [5342], 5 (1) [4985], 8 (6) [4920, 5485, 5501].
^Stemonitis fusca Roth, Bot. Mag. (Romer & Usteri) 1(2):26 (1787).
-
5 - 0,80 %: май - 3 (1,0 %), окт. - 2 (0,6 %). W - 5. Loc. 3 (1) [5332], 5 (1) [4982], 6 (1) [5385], 10 (1) [4929], 93 квартал (N 56°28.784', E032°56.306') (1) [4949].
Stemonitopsis typhina (F.H. Wigg.) Nann.-Bremek., Nederlandse Myxomyceten (Zutphen) 209 (1975) ["1974"].
" 1 - 0,16 %: май - 0 (0 %), окт. - 1 (0,3 %). В - 1. Loc. 1 (1).
*Trichia affinis de Bary, in Fuckel, Jahrb. Nassauischen Vereins Naturk. 23-24:336 (1870) ["1869"]. ’
1 - 0,16 %: май - 1 (0,3 %), окт. - 0 (0 %). W - 1. Loc. 3 (1) [5082].
Trichia botrytis (J.F. Gmel.) Pers., Neues Mag. Bot. 1:89 (1794).
-
22 - 3,5%: май - 2 (0,7 %), окт. - 20 (6,02 %). W - 16, В - 6. Loc. 1 (3) [5296, 5298], 2 (1) [5327], 3 (1) [5089], 4 (1) [5352], 5 (4) [5374], 6(4) [4897, 5396], 8 (3) [5499], 9 (5) [5472].
Trichia decipiens (Pers.) T. Macbr., N. Amer. Slime-moulds, ed. 1 218 (1899).
-
105 - 16,96 %: май - 59 (20,3 %), окт. - 46 (13,6%). W - 100, В - 4, B/L - 1. Loc. 1 (20) [4955, 5282, 5299], 2 (14) [5100, 5310], 3 (8) [5087, 5331], 4 (11) [4904, 5350], 5 (7) [4981, 4975, 5364], 6 (7) [4895, 5387], 7 (11) [4967, 5505], 8 (13) [4924, 5494, 5502], 9 (10) [5067, 5471], 10 (2) [4926], 81 квартал (N 56°28.899', E 032°58.817') (1) [4987], юг 89 квартала (N 56°28.89Г, E 032°58.676') (1) [4990].
#Trichia erecta Rex, Proc. Acad. Nat. Sci. Philadelphia 42:193 (1890).
-
2 - 0,32 %: май - 2 (0,7 %), окт. - 0 (0 %). W - 1, В - 1. Loc. 7 (2) [4963].
Trichia favoginea (Batsch) Pers., Neues Mag. Bot. 1:90 (1794).
-
84 - 13,3 8 % май - 61 (20,3 %), окт. - 23 (6,95%). W - 73, F - 1, В - 7, L - 3. Loc. 1 (13) [4950, 5287], 2 (7) [5091, 5325], 3 (4) [5078, 5330], 4 (6) [4903, 5356], 5 (10) [4973, 4983, 5362], 6 (17) [4892, 5390], 7 (6) [4972, 5507], 8 (11) [4911, 4915, 5105, 5488], 9 (6) [5065, 5473], 10 (3) [4925], 95 квартал (56°27.197', 032°58.854') (1) [4992].
#Trichia persimilis P. Karst., Not. Sallsk. Fauna Fl. Fenn. Forh 9:353 (1868).
-
10 - 1,59 %: май - 10 (3,3 %), окт. - 0 (0 %). W - 6, В - 3, L - 1. Loc. 3 (2) [5085], 4 (2) [4906], 6 (1) [4901], 7 (1) [4969], 8 (1) [4912], 9 (2) [5072], 10 (1) [4928].
*Trichia scabra Rostaf., Sluzowce monogr. 258 (1875).
-
13 - 2,07 %: май - 9 (3,0 %), окт. - 4 (1,21 %). W - 10, В - 3. Loc. 1 (2) [4958, 5286], 2 (2) [5098, 5320], 3 (3) [5079, 5346], 7 (2) [4962], 8 (1) [4910], 9 (2) [5068, 5074], 95 квартал (56°27.197', 032°58.854') (1) [4939].
tiTrichia subfusca Rex, Proc. Acad. Nat. Sci. Philadelphia 42:192 (1890).
-
2 - 0,32 %: май - 1 (0,3 %), окт. - 1 (0,3 %). W - 2. Loc. 8 (1) [5496], 9 (1) [5068].
Trichia varia (Pers, ex J.F. Gmel.) Pers., Neues Mag. Bot. 1:90 (1794).
-
24 - 3,82 %: май - 8 (2,7 %), окт. - 16 (4,83%). W - 12, В - 11, L - 1. Loc. 1 (4) [5101, 5285], 2 (2) [5305], 3 (2) [5329], 5 (1) [5371], 6 (1) [5391], 7 (2) [4961, 5510], 8 (3) [4918, 5491], 9 (5) [5073, 5467], 10 (1) [4927], 11 (2) [5478], юго-запад 92 квартала (N 56°27.619', E 032°58.136') (1) [4946].
Tubifera ferruginosa (Batsch) J.F. Gmel., Syst. nat. 2:1472 (1792) (sensu Martin, Alexopoulos, 1969).
-
2 - 0,32 %: май - 0 (0 %), окт. - 2 (0,6 %). W - 2. Loc. 6 (1) [5383], 8 (1) [5483].
He определенный вид, представитель nop. Physarales.
Спорофоры - спорангии на ножках, шаровидные, несколько поникающие, рассеянные или собранные в небольшие группы, светлофиолетового цвета. Перидий пленчатый, полупрозрачный, необызвествленный, неправильно растрескивающийся. Ножка по длине превосходит половину от общей высоты спороношения, необызвествленная, пленчатая, красновато-коричневая, морщинистая. Колонка отсутствует. Структуры капиллиция обнаружены не были. Споры 9-10 мкм в диам, черные в массе, фиолетово-коричневые в проходящем свете, орнаментированы бородавочками. Спорангии частично разрушены и, возможно, недоразвиты. Поскольку в образце отсутствуют структуры капиллиция, определить его принадлежность к тому или иному роду невозможно, однако по форме спороношения и цвету спор можно с уверенностью отнести его к представителям пор. Physarales. По цвету и форме спорангиев этот образец похож на Physarum pulcherrimum, однако подтвердить или опровергнуть это невозможно, т.к. структуры капиллиция отсутствуют. Это может объясняться недоразвитостью спороношения и/или его плохой сохранностью.
-
1 - 0,16 %: май - 1 (0,3 %), окт. - 0 (0 %). W - 1. Loc. 5 (1) [4986].
Перечень видов, о нахождении которых сообщается в работе Ю.К. Новожилова (1980), которые не были отмечены в ходе сборов в 2014 году:
Ceratiomyxa fruticulosa (O.F. Mull.) T. Macbr.; Craterium minutum (Leers) Fr.; Cribraria piriformis Schrad. Diderma niveum (Rostaf.) E. Sheld.; Didymium nigripes (Link) Fr.; Hemitrichia intorta (Lister) Lister; Perichaena corticalis (Batsch) Rostaf.; Physarum leucophaeum Fr. et
Palmquist Physarum tenerum Rex; Physarum vernum Sommerf., in Fries; Trichia verrucosa Berk., in Hooker; Tubifera microsperma (Berk, et M.A. Curtis) G.W. Martin.
Таким образом, в настоящий момент, с учетом литературных данных, на территории ЦЛГПБЗ обнаружено 82 вида миксомицетов из 6 порядков, 11 семейств и 28 родов.
Анализ таксономической структуры миксомицетов, собранных на территории ЦЛГПБЗ в 2014 году
В 2014 году на территории ЦЛГПБЗ был собран 621 образец, относящийся к 70 видам из 25 родов, 10 семейств и 5 порядков. Таким образом, список миксомицетов ЦЛГПБЗ увеличился на 45 видов. Кроме того, обнаружено 7 образцов, для которых видовая принадлежность не установлена, а еще один образец был определен только до порядка. Полная таксономическая структура биоты миксомицетов ЦЛГПБЗ по результатам сборов 2014 года приведена в таблице 1.
По результатам сборов в 2014 году как наибольшей частотой встречаемости (65,7%), так и видовой насыщенностью (27 видов) обладает порядок Trichiales. Порядки Physarales, Liceales и Stemonitales представлены 15, 14 и 12 видами соответственно. Наименьшей видовой насыщенностью обладает пор. Echinosteliales, что объясняется как мелкими размерами спороношений, которые сложно обнаружить в ходе полевых исследований, так и незначительным общим числом описанных видов. Данная таксономическая структура в значительной степени отличается от типичной для наиболее хорошо изученных территорий, где обычно по числу обнаруженных видов преобладают представители пор. Physarales (Гмошинский, 2013). Скорее всего, это объясняется тем, что исследования были проведены только ранней весной и в середине осени. В то время как многие виды миксомицетов подчас обладают достаточно хорошо выраженной сезонной приуроченностью (Власенко, Новожилов, 2011), для выявления наиболее полного видового разнообразия, а, следовательно, и таксономический структуры, требуется проведение исследований в течение всего бесснежного периода.
По частоте встречаемости образцов, доминирующее положение в биоте занимают представители пор. Trichiales, спороношения которых составляют 65,7% от всего собранного материала. Представители пор. Physarales, Liceales и Stemonitales представлены 13,9%, 10,6% и 8,7% обнаруженных образцов, соответственно.
Наименьшей частотой встречаемости опять же обладал пор. Echinosteliales - 1,1%.
На уровне семейств таксономическая структура биоты миксомицетов ЦЛГПБЗ по результатам сборов 2014 года выглядит следующим образом: Наибольшим видовым разнообразием обладают семейства Trichiaceae и Stemonitidaceae (15 и 12 видов соответственно); сем. Arcyriaceae, представлено 11 видами. Кроме того, значительной видовой насыщенностью обладают семейства Physaraceae (9 видов), Cribrariaceae (8 видов) и Didymiaceae (6 видов). Остальные семейства представлены тремя (Liceaceae и Reticulariaceae), двумя (Clastodermataceae) и одним видом (Dianemataceae).
При рассмотрении частоты встречаемости представителей отдельных семейств можно заметить, что доминирующее положение в биоте миксомицетов ЦЛГПБЗ по результатам сборов 2014 года занимают представители сем. Trichiaceae. Их частота встречаемости составляет 57%. Такое доминирующее положение в биоте можно опять же объяснить сезонными особенностями сбора материала. Среди остальных семейств, на долю которых приходится менее 43 %, доминирующее положение занимает сем. Stemonitidaceae (8,7 %), Arcyriaceae (8,1%) и Physaraceae (8,2%).
Таблица 1
Таксономическая структура биоты миксомицетов ЦЛГПБЗ по результатам сборов 2014 г.
|
Класс |
Порядок |
Семейство |
Род |
Число видов |
|
Myxomycetes (70) |
Echinosteliales (2) |
Clastodermataceae (2) |
Clastoderma |
1 |
|
Barbeyella |
1 |
|||
|
Liceales (14) |
Cribrariaceae (8) |
Cribraria |
8 |
|
|
Liceaceae (3) |
Licea |
3 |
||
|
Reticulariaceae (3) |
Lycogala |
2 |
||
|
Tubifera |
1 |
|||
|
Trichiales (27) |
Trichiaceae (15) |
Trichia |
9 |
|
|
Metatrichia |
2 |
|||
|
Hemitrichia |
3 |
|||
|
Arcyodes |
1 |
|||
|
Dianemataceae (1) |
Calomyxa |
1 |
||
|
Arcyriaceae (11) |
Arcyria |
11 |
||
|
Stemonitales (12) |
Stemonitidaceae (12) |
CoIlaria |
1 |
|
|
Comatrichia |
4 |
|||
|
Enerthenema |
1 |
|||
|
Lamproderma |
3 |
|||
|
Stemonitis |
2 |
|||
|
Stemonitopsis |
1 |
|||
|
Physarales (15) |
Didymiaceae (6) |
Diderma |
2 |
|
|
Didymium |
2 |
|||
|
Lepidoderma |
1 |
|||
|
Mucilago |
1 |
|||
|
Physaraceae (9) |
Fuligo |
3 |
||
|
Leocarpus |
1 |
|||
|
Physarum |
5 |
По результатам сборов 2014 года наибольшей видовой насыщенностью обладали следующие роды: Arcyria (12 видов), Trichia (9 видов), Cribraria (8 видов), Physarum (5 видов), Comatricha (4 вида), Hemitrichia, Lamproderma, Fuligo и Licea (по 3 вида).
С. Стефенсоном было предложено разделение биоты миксомицетов по группам частот встречаемости (Stephenson, 1988): R - редкий вид (0,5%); О - изредка встречающийся (0,5%-1,5%); С -обычный (1,5%-3%); А - вид с высокой степенью встречаемости (>3%). Виды, включенные в категорию А, составляют ядро биоты. В 2014 году таких видов было 9: Trichia decipiens (частота встречаемости 16,85%), Т. favoginea (13,35%), Metatrichia vesparia (5,56%), Physarum album (5,41%), Lycogala epidendrum (4,45%), T. varia (3,82%), Diderma radiatum (3,82%), T. botrutis (3,5%) и Hemitrichia serpula (3,18%). Таким образом, из 9 видов, составляющих ядро биоты, 4 относятся к роду Trichia, и 5 к пор. Trichiales.
Примечательным является факт того, что в список наиболее широко распространенных видов на территории ЦЛГПБЗ включен D. radiatum. Этого представителя пор. Physarales на территории Москвы и Московской области удается обнаружить достаточно редко. За все время исследований видового разнообразия этот вид был отмечен лишь 4 раза (Гмошинский, 2013). Причин этому может быть две: либо данный вид приурочен к более холодному климату, либо он чувствителен к загрязнениям и не образует спороношения в присутствии каких-либо поллютантов. В пользу первой версии может выступать факт обнаружения большого числа спороношений данного вида в регионах с более холодным климатом (по берегам Кандалакшского залива Белого моря) (Гмошинский, 2014). В пользу второй гипотезы могут выступать данные, полученные в Б.С. Плотниковым и К.А. Фефеловым. В ходе исследований, проведенных в окрестностях медеплавильного комбината на Урале (Плотников, Фефелов, 2003; 2009; Плотников, 2010), было установлено, что загрязнение атмосферы оксидами серы вызывает уменьшение видового разнообразия представителей пор. Physarales и при этом увеличение доли видов, относящихся к пор. Trichiales и Liceales. В ходе исследований, проведенных на территории г. Москвы, данную гипотезу подтвердить не удалось, однако можно предположить, что возможно существуют определенные виды, численность которых сокращается в присутствии разного рода поллютантов. Поэтому следует обратить особое внимание на численность данного вида как в северных регионах с высокой степенью загрязнения, так и в южных с относительно благополучной экологической ситуацией.
К категории С, часто встречающихся видов, относятся также 9 видов, в то время как к категории О, изредка встречающихся, относятся еще 11 видов. Остальные 41 вид и 6 неопределенных до вида морф относятся к категории редких, т.е. эти виды составляли менее чем в 0,5% от общего числа образцов. Таким образом, в ходе исследований более 50% видов было представлено менее чем 3 образцами, а, следовательно, их обнаружение в определенной степени является случайным, что в свою очередь свидетельствует о необходимости проведения дальнейших исследований видового разнообразия на территории ЦЛГПБЗ в том числе и в весенне-осенний период.
Сравнение видового разнообразия миксомицетов ЦЛГПБЗ в весенний и осенний периоды
В ходе проведенных сборов спороношений миксомицетов на территории ЦЛГПБЗ в 2014 году было обнаружено 628 образцов, таксономическое положение которых удалось определить до рода. В мае собрано 297 образцов, в октябре -331 образец. Таким образом, в каждый из сезонов собрано приблизительно равное число образцов, а, следовательно, можно проводить сравнение видовой насыщенности биоты в каждый из сезонов.
При сравнении видового разнообразия в каждый из сезонов можно заметить, что осенью выявляемое видовое разнообразие значительно выше, чем весной: 64 и 39 видов соответственно.
Таблица 2
Ядро биоты миксомицетов по результатам сборов в мае и октябре
|
Вид |
май |
октябрь |
||
|
Число образцов |
Частота встречаемости |
Число образцов |
Частота встречаемости |
|
|
Trichia favoginea |
61 |
20,5% |
23 |
6,9% |
|
Trichia decipiens |
59 |
19,9% |
45 |
13,6% |
|
Metatrichia vesparia |
26 |
8,8% |
9 |
2,7% |
|
Hemitrichia serpula |
18 |
6,1% |
2 |
0,6% |
|
Diderma radiatum |
14 |
4,7% |
10 |
3,0% |
|
Lycogala epidendrum |
10 |
3,4% |
19 |
5,7% |
|
Trichia persimilis |
10 |
3,4% |
0 |
0,0% |
|
Hemitrichia clavata |
9 |
3,0% |
6 |
1,8% |
|
Trichia scabra |
9 |
3,0% |
4 |
1,2% |
|
Trichia varia |
8 |
2,7% |
16 |
4,8% |
|
Trichia botrytis |
2 |
0,7% |
20 |
6,0% |
|
Arcyria affinis |
0 |
0,0% |
18 |
5,4% |
|
Physarum album |
0 |
0,0% |
34 |
10,3% |
Ядро биоты миксомицетов в мае и октябре представлено в таблице 2. Только 4 вида: Trichia favoginea, Т. decipiens, Diderma radiatum и Lycogala epidendrum составляют ядро биоты и в мае и в октябре. При этом три вида, входящие в ядро биоты в один из периодов, не обнаружены в другой. Так, Trichia persimilis, представленный 10 образцами в мае, не был отмечен в октябре, в то время как Arcyria affinis и Physarum album, представленные в октябре 18 и 34 образцами, в мае обнаружены не были.
При рассмотрении таксономической структуры биоты на уровне семейств можно сделать вывод, что их относительная видовая насыщенность в значительной степени различается в зависимости от времени года. При сопоставлении видовой насыщенности семейств миксомицетов можно заметить, что в октябре она снижается у представителей сем. Trichiaceae, Cribrarariaceae и Didymiaceae, в то время как число видов представителей сем. Arcyriaceae, Physaraceae, Reticulariaceae и Liceaceae в октябре в значительной степени возрастает.
По частоте встречаемости на уровне семейств можно также заметить значительные различия в зависимости от сезона года. Так семейство Trichiaceae, частота встречаемости представителей которого в мае составляет 73,7%, в октябре представлено лишь 40,5% от общего числа образцов. Сходная динамика наблюдается и для представителей сем. Didymiaceae (6,4% в мае против 4,8% в октябре) и Cribrariaceae (5,7% против 4,5%). Остальные семейства в осенние месяцы представлены большим числом образцов. Наиболее выражена подобная динамика для представителей сем. Arcyriaceae (1,3% в мае против 14,2% в октябре) и Physaraceae (1,7% в мае против 14,8% в октябре).
Интересно отметить, что, несмотря на различия в таксономической структуре, на уровне видов, родов и семейств в мае и в октябре, относительные значения видовой насыщенности оказались схожими, чего нельзя сказать об относительной частоте встречаемости представителей этих порядков. Частота встречаемости в значительной степени снижается для представителей пор. Trichiales, в то время как для представителей остальных порядков она в значительной степени возрастает. Особенно хорошо это выражено для представителей пор. Physarales, доля которых в осенние месяцы возрастает более чем в два раза (8,1% против 19,6%).
Авторы выражают благодарность Н.И. Киреевой за выполненный ей рисунок B.minutissima, а также сотрудникам Меж факультетской лаборатории электронной микроскопии МГУ за предоставление приборной базы для получения фотографий на сканирующем электронном микроскопе, использованных в ходе определения.
Список литературы Новые данные о видовом разнообразии миксомицетов ЦЛГПБЗ (Нелидовский р-н, Тверская область)
- Власенко А.В., Новожилов Ю.К. 2010. Редкие и новые для России виды миксомицетов (Myxomycetes) из сосновых лесов правобережья верхнего приобья//Микология и фитопатология. Т. 44. №. 2. С. 99-108.
- Власенко А.В., Новожилов Ю.К. 2011. Фенологические особенности миксомицетов сосновых лесов правобережной части верхнего Приобья//Растительный мир Азиатской России. № 2. Вып. 8. С. 3-8.
- Гмошинский В.И. 2013. Миксомицеты Москвы и Московской области. Дисс. на соиск. степ. канд. биол. наук. 698 с.
- Новожилов Ю.К. 1980. Миксомицеты Центрально-лесного государственного заповедника и Ленинградской области//Микология и фитопатология. Т. 14. Вып. 3. С. 198-201.
- Новожилов Ю.К. 2005. Миксомицеты (класс Myxomycetes) России: Таксономический состав, экология и география: дис. … д-ра биол. наук. СПб., БИН РАН. 377 c.
- Плотников Б.С. 2005. Систематическая структура сообществ миксомицетов в градиенте техногенного загрязнения//Экология: от генов до экосистем: мат-лы конф. молодых ученых. Екатеринбург. С. 206-211.
- Плотников Б.С., Фефелов К.А. 2003. Таксономический состав миксомицетов города Екатеринбурга//Проблемы глобальной и региональной экологии: Матер. конф. молод. ученых 31 мар.-4 апр. 2003. Екатеринбург: «Академкнига». 372 с.
- Плотников Б.С., Фефелов К.А. 2009. Миксомицеты южной тайги среднего Урала в градиенте промышленного загрязнения выбросами медеплавильного комбината//Микология и фитопатология. Том. 43. Вып. 1. С. 33-44.
- Lado C. 2005-2015. An on line nomenclatural information system of Eumycetozoa. Real Jardín Botánico, CSIC. Madrid, Spain. 2005-2015. http://www.nomen.eumycetozoa.com (дата обращения 02.04.2015).
- Martin G.W., Alexopoulos C.J. 1969. The Myxomycetes//Iowa City: Univ. of Iowa Press. 561 p.
- Nannenga-Bremekamp N.E. 1991. A guide to temperate myxomycetes. Biopress Ltd. 409 p.
- Poulain M., Meyer M., Bozonnet J. 2011. Les Myxomycétes. Tome 1, guide de détermination. mycologique et botanique Dauphiné-Savoie: Sévrier France. 568 p., 15 pl.
- Schnittler M., Stephenson S.L., Novozhillov Yu.K. 2000а. Ecology and world distribution of Barbeyella minutissima (Myxomycetes)//Mycological Research. Vol. 104. No. 12. P. 1518-1523.
- Schnittler M., Stephenson S.L., Novozhillov Yu.K. 2000b. Ultrastructure of Barbeyella minutissima (Myxomycetes)//Karstenia. Vol. 40. P. 159-166.
- Stephenson S.L. 1988. Distribution and ecology of Myxomycetes in temperate forests I. Patterns of occurrence in the upland forests of southwestern Virginia//Canad. J. Bot. Vol. 66. P. 2187-2207.
- Yamamoto Y. 1998. The Myxomycete biota of Japan. Tokyo: Toyo Shorin Publishing Co., Ltd. 700 p.
