Новые подходы к решению проблемы фациальности листа с помощью молекулярно-генетических методов

Автор: Озерова Людмила Викторова, Тимонин А.К., Милютина И.А., Красникова М.С., Боброва В.К.
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Материалы лекций и устных докладов
Статья в выпуске: 9, 2008 года.
Бесплатный доступ
Развитие абаксиально-адаксиальной полярности листа у дву- и однодольных растений инициируется регулируемой активностью микроРНК (miRNA). Эти РНК, в свою очередь, регулируют накопление мРНК транскрипционных факторов, требуемых для специфической модуляции генной активности в тканях развивающегося листа. Последовательное действие двух микроРНК (miR390 и TAS3 tasiARF РНК) длиной 21 нуклеотид контролирует активность генов факторов транскрипции ARF3 и ARF4 у Arabidopsis thaliana. Нами изучено наличие генов предшественников tasiARF РНК у видов рода Senecio с би- и унифациальным типом развития листа. Оказалось, что оба типа видов кодируют такие РНК, структурно не отличающиеся от найденных у арабидопсиса. Полученные данные позволят в дальнейшем изучить регуляцию экспрессии tasiARF РНК у различных видов рода Senecio и понять вклад TAS3-подобных генов в развитие симметрии листа у растений этого рода.
Рнк
Короткий адрес: https://sciup.org/146116178
IDR: 146116178 | УДК: 581.452:
Текст научной статьи Новые подходы к решению проблемы фациальности листа с помощью молекулярно-генетических методов
-
1 Главный ботанический сад им. Н.В. Цицина РАН, Москва
-
2 Московский государственный университет им. М.В. Ломоносова, Москва 3Научно-исследовательский институт физико-химической биологии им. А. Н.
Белозерского МГУ им. М. В. Ломоносова, Москва
Развитие абаксиально-адаксиальной полярности листа у дву- и однодольных растений инициируется регулируемой активностью микроРНК (miRNA). Эти РНК, в свою очередь, регулируют накопление мРНК транскрипционных факторов, требуемых для специфической модуляции генной активности в тканях развивающегося листа. Последовательное действие двух микроРНК (miR390 и TAS3 tasiARF РНК) длиной 21 нуклеотид контролирует активность генов факторов транскрипции ARF3 и ARF4 у Arabidopsis thaliana. Нами изучено наличие генов предшественников tasiARF РНК у видов рода Senecio с би- и унифациальным типом развития листа. Оказалось, что оба типа видов кодируют такие РНК, структурно не отличающиеся от найденных у арабидопсиса. Полученные данные позволят в дальнейшем изучить регуляцию экспрессии tasiARF РНК у различных видов рода Senecio и понять вклад TAS3-подобных генов в развитие симметрии листа у растений этого рода.
Проблема фациальности листа проявляется двояко. С одной стороны, это – изучение закономерностей формирования присущей листу характерной бифациальной структуры в ходе его образования на унифациальном апексе побега. С другой стороны, это – познание специфики появления унифациальных листьев вследствие исчезновения у них адаксиальной стороны. Современная молекулярная генетика позволила перейти от чисто дискурсивного исследования фациальности листа к экспериментальному изучению глубинных факторов, определяющих адаксиальноабаксиальную полярность листового примордия. При этом в центре внимания генетиков оказалась только первая сторона проблемы, в ходе решения которой был выявлен целый ряд генов и установлены основные взаимодействия между ними [1]. Вместе с тем высокая затратность самого исследования, основанного на клонировании протяженных фрагментов генома, существенно лимитирует расширение круга изучаемых обхъектов.
Новые возможности появились с открытием роли регулируемой активности микроРНК (miRNA) в становлении адаксиально-абаксиальной полярности листового примордия у Arabidopsis thaliana (L.) Heynh. и Zea mays L. [5; 7; 12]. Эти микроРНК регулируют накопление мРНК транскрипционных факторов, необходимых для специфической модуляции генной активности в тканях развивающегося примордия [8; 9]. Последовательное действие двух микроРНК: miR390 и TAS3 tasiARF, длиной 21 нуклеотид, контролирует активность генов факторов транскрипции ARF3 и ARF4, направляя специфическую деградацию их мРНК нуклеазой РНК-индуцируемого комплекса сайленсинга (RISC) [5; 7; 12].
РНК tasiARF образуется из полиаденилированного РНК-предшественника, названного TAS3, который транскрибируется в ядре клетки ДНК-зависимой РНК-полимеразой II. Как и обычные РНК, кодирующие белки, он содержит на концах кэп и поли (А)-последовательность нуклеотидов (рис. 1). Кроме того, в нем имеются два участка узнавания специфическим белковым комплексом, содержащим нуклеазу AGO
[6; 7] и РНК miR390, но у A. thaliana только в одном из участков, расположенном со стороны поли (А)-конца, происходит первичное расщепление молекулы предшественника [5]. После того как содержащаяся в комплексе РНК miR390 комплементарно узнает РНК TAS3, последняя расщепляется с участием нуклеазы DCL, образуя tasiARF (рис. 1) [5; 7]. У A. thaliana молекула РНК TAS3 содержит две тандемно повторяющиеся последовательности нуклеотидов и при расщеплении дает соответственно две молекулы tasiARF (рис. 1). Однако у Z. mays есть четыре TAS3-подобных РНК, и одна из них дает при расщеплении единственную молекулу tasiARF, поскольку содержит только одну соответствующую последовательность нуклеотидов [12].
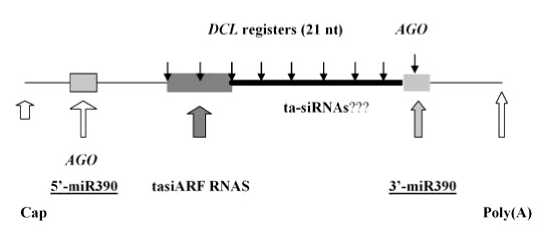
Рис. 1. Принципиальная схема организации TAS3 РНК у растений
Незаштрихованные стрелки указывают положение кэп-структуры, поли- (А) и ненасыщенного участка узнавания miR390. Заштрихованные стрелки указывают положение расщепляемого участка узнавания miR390 и тандемного повтора tasiIARF РНК. Черные стрелки указывают точки расщепления нуклеазой AGO и 21-нуклеотидную рамку разрезания нуклеазой DCI.
Важное значение генов TAS в установлении адаксиально-абаксиальной полярности листовых примордиев у A. thaliana и Z. mays , а также относительная легкость их выявления делают их удобным средством решения проблемы фациальности листа и понуждают к поиску подобных генов и у других покрытосеменных. Особый интерес в этой связи представляют виды рода Senecio – S. talinoides (DC) Sch. Bip., S. articulatus (Haw.) Sch. Bip., различающиеся по фациальности листьев [3].
У S. articulatus почка малой емкости всего с 2-3 формирующимися листьями. На периферии плоского апекса закладываются листовые примордии (рис. 2, А) в виде уплощенных бугорков. Примордии удлиняются и расширяются тангентально, приобретая уплощенную форму (рис. 2, Б), так что у них становится возможным различить ад- и абаксиальные стороны. Удлинение средней части примордия резко интенсифицируется по сравнению с его боковыми частями (рис. 2, В). Примерно на этой же стадии примордий слегка загибается над апексом и начинается утолщение медиальной области Unterblat и всего Oberblatt1. Вследствие этого верхушка примордия приобретает параболоидальную форму без границы между ад- и абаксиальные сторонами. Ниже параболоидальной верхушки примордий остается бифациальным. Рост параболоидальной верхушки примордия почти сразу прекращается, а из бифациальной его части начинает формироваться основная часть листовой пластинки. При этом не только происходит ее удлинение и расширение, но и путем локализованного роста в акропетальной последовательности закладываются по 2 главные лопасти листовой пластинки (рис. 2, В). Ниже базальных лопастей листовой пластинки примордий утолщается благодаря расширению абаксиальной стороны. В результате под листовой пластинкой возникает субунифациальный участок, дающий начало субунифациальному черешку (рис. 2, Г).

Рис. 2. Листья S. Articulatus на разных стадиях развития
А – апекс с самым молодым листовым примордием; Б – дифференциация примордиев на Unter-и Oberblatt; В – дифференциация лопастей листовой пластинки; Г – дефинитивный лист.
Условные обозначения: a – апекс; p – листовой приморди; o – Oberblatt, u – Unterblatt, ad – адаксиальная сторона; ab – абаксиальная сторона.
Таким образом, листья у этого вида почти целиком бифациальные (за исключением очень короткого остроконечия листовой пластинки).
У S.talinoides терминальная почка малой емкости. Апекс варьирует от едва выпуклого до слегка вогнутого (рис. 3, А, Б). Лист закладывается на периферии апекса в виде небольшого примордия, эллиптического в поперечном сечении (рис. 3, А, Б). Удлинение примордия сочетается с его расширением вследствие распространения меристематической активности по окружности узла в обе стороны от медианной плоскости примордия. После такого равномерного роста наступает дифференциация примордия на короткий базальный желобчатый, серповидный в поперечном сечении, Unterblatt и дистальный почти цилиндрический Oberblatt. (рис. 3, Б). Считающаяся универсальной для листьев двудольных растений [2] стадия 3-лопастного зачатка в морфогенезе листа S. talinoides отсутствует. Дифференциация листового примордия у этого вида выражается в усиленном продольном росте его медианного участка при ослаблении роста краевых участков. Одновременно с продольным ростом медианный участок существенно утолщается и округляется в поперечном сечении. Unterblatt с четко различимыми краями и адаксиальной и абаксиальной сторонами впоследствии растет слабо и становится коротким бифациальным основанием листа (рис. 3, Г).
Ставший унифациальным цилиндрический Oberblatt продолжает интенсивно расти в длину, причем этот рост довольно быстро сосредоточивается в его основании, по соседству с Unterblatt (рис. 3, В). Благодаря такому длительному интеркалярному росту возникает характерная для этого вида вальковатая унифациальная пластинка листа, сидящая на коротком бифациальном основании (рис. 3, Г). По мере роста листа и Unterblatt, и Oberblatt становятся толще, что, однако, никак не сказывается на их фациальности [4].
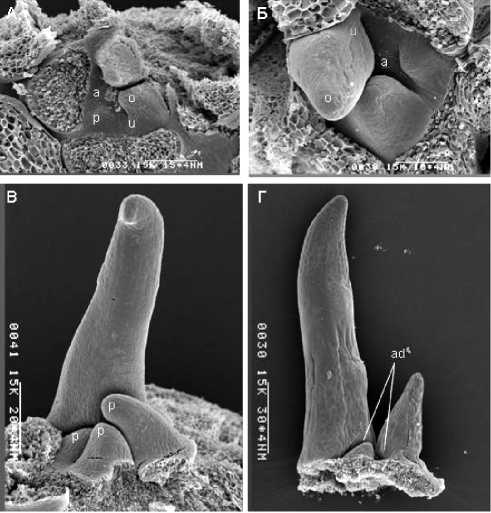
Рис. 3. Листья S.talinoides на ранних стадиях развития
А – апекс с самым молодым листовым примордием; Б – дифференциация примордиев на Unter-и Oberblatt; В – лист на стадии интеркалярного роста; Г – дефинитивный унифациальный лист. Условные обозначения: a – апекс, p – листовой примордий, o – Oberblatt, u – Unterblatt, ad – адаксиальная сторона
Следовательно, листья у этого вида почти целиком унифациальные, и лишь их очень короткое основание сохраняет бифациальное строение.
Для выявления TAS-подобных генов у обоих видов Senecio был применен новый, не описанный в литературе подход: вместо клонирования протяженных фрагментов генома, используя консервативные участки узнавания miR390, мы провели прямую амплификацию этих генов. Судя по выявленным нуклеотидным последовательностям, у обоих видов Senecio есть TAS-подобные гены, структурно сходные с типичными TAS3 генами A. thaliana – они содержат такие же тандемные повторы и потенциально способны продуцировать tasiARF РНК.
Таким образом, к настоящему времени TAS3 гены обнаружены у A. thaliana [5]., Z. Mays [12]., видов Senecio , а также у Nicotiana tabacum L. и Polygonum arenastrum Boreau (наши неопубл. данные), которые соответственно представляют Rosidae в узком смысле (= «eurosidae II»), Liliopsida, обе основные клады Asteridae («euasteridae I» и «asteridae II») и базальные Rosidae. Такая встречаемость TAS3 генов позволяет предположить, что они представляют собой универсальный компонент генома 184
покрытосеменных растений, регулирующий становление фациальности листа в онтогенезе и, вероятно, в эволюции. При этом обращает на себя внимание то, что S. spiculosus с унифациальными листьями имеет те же гены TAS3, что и S. articulatus и другие изученные виды, которым свойственны бифациальные листья. Поэтому развитие унифациальных листьев S. talinoides отнюдь не обусловлено простой делецией генов регуляторных tasiARF РНК. Очевидно, что подобная делеция была бы попросту невозможна, поскольку листовые примордии у этого вида проходят стадию полностью бифациальной структуры. Их последующее преобразование в унифациальный орган, вероятнее всего, обусловлено различиями в транскрипционной активности генов TAS3 в разных участках развивающегося листа.
На следующем этапе работы мы предполагаем изучить транскрипционную активность TAS генов в развивающемся листе, что, возможно, позволит оценить вклад модуляции накопления miRNA в механизм развития унифациальных листьев. Другие подходы будут включать прямое сравнение накопления в би- и унифациальных листьях регуляторых РНК miR390 и miR166.
Благодарности
Мы глубоко признательны всем сотрудникам Лаборатории электронной микроскопии биологического факультета Московского государственного университета им. М.В. Ломоносова, зав. лабораторией генной инженерии вирусов Научно-исследовательского института физико-химической биологии им. А.Н. Белозерского МГУ им. М. В. Ломоносова проф., д.б.н. С.Ю. Морозову.
Исследование поддержано Федеральным агентством по науке и инновациям (Роснаука), государственный контракт № 02. 518.11.7090, а также грантом РФФИ 06-04-49493.
Abaxial-adaxial symmetry during leaf development of flowering plants is known to be controlled by microRNAs (miRNAs). These RNAs influence accumulation of mRNAs encoding transcription factors required for specific modulation of gene expression in tissues of developing leaf. Sequential activity of miR390 and TAS3 tasiARF RNAs (21 nucleotides in length) controls synthesis of transcription factors ARF3 and ARF4 in Arabidopsis thaliana. We have studied the presence of tasiARF RNA genes in the representatives of genus Senecio having either unifacial or true bifacial leaves. It was found that both types of species encode RNA precursors principally similar to those found in Arabidopsis. The data obtained will be used for further studies of expression regulation of Senecio TAS3-like genes and understanding the potential contribution of these RNAs in development of plant organ abaxial-adaxial symmetry .
