О целесообразном характере окраски антарктической рыбы Dolloidraco longedorsalis (Artedidraconidae; Notothenioidei; Perciformes), живущей там, куда не проникает видимый солнечный свет
, живущей там, куда не проникает видимый солнечный свет")
Автор: Балушкин А.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 4, 2017 года.
Бесплатный доступ
Выполнено описание особенностей окраски (глазчатые пятна на хвостовом плавнике, камуфляж настоящих глаз) у антарктической рыбы Dolloidraco longedorsalis (сем. Artedidraconidae) и обсуждены вопросы их целесообразности в условиях больших глубин.
Семейство artedidraconidae, окраска тела, самомимикрия, глазчатые пятна, эффект вавилова-черенкова, южный океан
Короткий адрес: https://sciup.org/147204852
IDR: 147204852 | УДК: 59
On the purposive coloration of the antarctic fish Dolloidraco longedorsalis (Artedidraconidae; Notothenioidei; Perciformes), inhabiting layers, where the visible sunlight cannot permeate
The peculiarities of coloration (false eyespots on the caudal fin, camouflage of the authentic eyes) in the Antarctic fish Dolloidraco longedorsalis (Artedidraconidae) are described and the questions of their fitness in deep water conditions are discussed.
Текст научной статьи О целесообразном характере окраски антарктической рыбы Dolloidraco longedorsalis (Artedidraconidae; Notothenioidei; Perciformes), живущей там, куда не проникает видимый солнечный свет
Длиннопёрые бородатки Долло Dolloidraco longe-dorsalis Roule, 1913 – это небольшие рыбки стандартной длиной не более 10 см, населяющие мезобен-тальные глубины шельфа и верхнего склона Антарктиды и близлежащих островов. Они относятся к самому южному в современной ихтиофауне семейству бородатковых Artedidraconidae, включающему 4 рода: Artedidraco Lönnberg, 1905 (7 видов), Pogono-phryne Regan, 1914 (29 видов) и монотипические Dol-loidraco Roule, 1913 и Histiodraco Regan, 1914 [Спо-дарева, Балушкин, 2014; Балушкин, Сподарева, 2015; Eakin, Riginella, La Mesa, 2015]. Морфологически Dolloidraco наиболее резко обособлен от других родов семейства, характеризуясь крупными и высоко посаженными на голове глазами, удлиненными хво- стовыми позвонками, смещённым к голове колючим спинным плавником, неветвистыми лучами в анальном и втором спинном плавниках и другими признаками. К числу последних относится окраска тела, анализу которой и будет посвящена настоящая работа. Самая необычная особенность окраски длиннопёрых бородаток – присутствие на хвостовом плавнике крупного глазчатого пятна.
Описанию глазчатых пятен (глазков, ложных или фальшивых глаз) в различных классах животных посвящена огромная научная, а особенно -научно-популярная литература. В дарвиновской теории эволюции и в её дериватах (неодарвинизм, синтетическая теория эволюции) их появление у видов рассматривалось в качестве созданного в ходе естественного отбора приспособления для пассивной защиты от хищников. В учебниках лож-
ные глаза принято относить к категории угрожающей или устрашающей окраски [см.: Шмальгаузен, 1969, с. 91], иногда их также считают примером самомимикрии [Cott, 1940].
Надо сказать, что не все примеры пользы ложных глаз в борьбе за выживание имели достаточное научное обоснование. На это давно обратил внимание наш выдающийся отечественный мыслитель Николай Яковлевич Данилевский – один из первых авторитетных критиков теории «естественного подбора». Если он и мог допускать возможность отпугивания хищника с помощью активной демонстрации глазков, то категорически возражал против какого-либо участия отбора в создании пассивных средств защитной окраски, проводя аналогии о бесполезности рисования китайцами драконов или других чудовищ для устрашения войск, с которыми им приходилось сражаться, или вывешивания пугал от птиц на огородах и плодо- вых садах, на которые после короткого опыта птицы преспокойно садятся [Данилевский, 1885, с. 165–166]. К настоящему времени в экспериментах, проводимых экологами, доказательная база существенно улучшена. В литературе на примерах разных животных (бабочки, рыбы, птицы) известно несколько гипотез, объясняющих пользу ложных глаз для организма: гипотезы запугивания хищника, отклонения хищника на менее жизненно важные части тела жертвы, сигнализация о статусе и как просто неиспользуемый более рудимент [Котт, 1950; Blest, 1957; Neudecker, 1989; Stevens, 2005; Lönnstedt, McCormick, Chivers, 2013]. Для рыб наиболее убедительные результаты, указывающие на пользу ложных глаз для выживания молоди, были получены в недавних экспериментах с рыба- ми-ласточками Pomacentrus amboinensis Bleeker, 1868 (сем. Pomacentridae) [Lönnstedt, McCormick, Chivers, 2013]. Ниже при обсуждении наших материалов мы немного подробнее коснёмся результатов этих опытов.
Материалы
В основе настоящей работы лежит изучение музейных коллекций Зоологического музея Гамбургского университета (Zoologisches Institut und Museum, Universität Hamburg) и лаборатории ихтиологии Зоологического института РАН. Рыбы были пойманы в морях Уэдделла (глубины 210–1 168 м), Лазарева (350–370 м), Моусона (460–550 м) и в море Росса (640–700 м). Всего было исследовано 66 экз. стандартной длиной 36.9–90.8 мм.
Результаты и их обсуждение
Ложный глаз у длиннопёрой бородатки представляет собой крупное белое пятно в центральной части хвостового плавника. По периметру его окаймляют широкая тёмная полоса на основании плавника спереди, серия прерывистых тёмных пятен сверху, широкая тёмная полоса чуть впереди кончиков лучей С сзади и тёмные основания самых нижних лучей плавника снизу (рис. 1). Нам представляется, что при использовании глазчатых пятен длиннопёрые бородатки, помимо очевидной дезориентации хищника об их пространственной ориентации и возможных путях отступления, умело применили и другие стратегии защиты: камуфляж истинного глаза и отпугивание (устрашение) посредством активной демонстрации глазка.
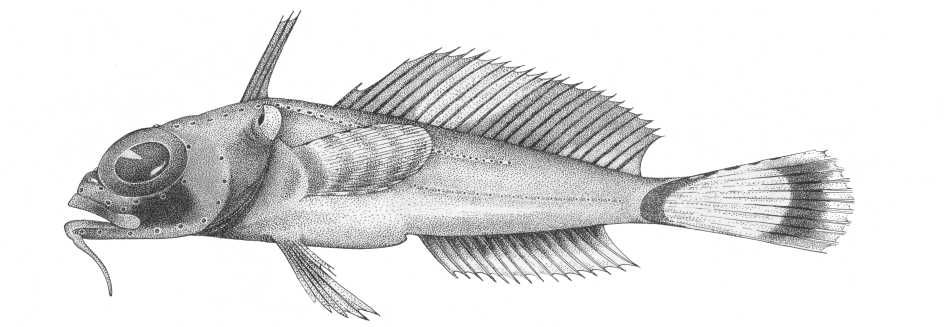
Рис. 1 . Длиннопёрая бородатка Долло Dolloidraco longedorsalis [по Балушкин, Сподарева, 2015]
Последнее, как это хорошо известно из классических примеров с глазками бражников [см. например: Шмальгаузен, 1969; Мариковский, 1977; Дольник, 2009], предусматривает необходимость предварительной маскировки глазчатых пятен, позволяющей создать эффект неожиданности. Будучи до поры до времени скрытыми от хищника, устра- шающие глаза могут стать решающим фактором при бегстве от хищника, давая возможность выиграть время в первые (и самые важные) секунды его атаки. У D. longedorsalis крупные ложные глаза чётко выявляются только на расправленном хвостовом плавнике и, таким образом, расправленный хвостовой плавник у D. longedorsalis может слу- жить одновременно и средством отступления (как движитель) и отпугивания хищника. Как это не покажется странным, надежность предварительного сокрытия ложного глаза хорошо подтверждается практикой описания ихтиологами окраски длиннопёрых бородаток. В научных коллекциях хвостовой плавник рыб зафиксирован, как правило, в сложенном виде, при котором пятно оказывается сокрытым (рис. 2А, 2Б). Отсюда при описании окраски этого вида авторы замечали на хвостовом плавнике только 2 широкие тёмные полосы [Waite, 1916; Norman, 1938; DeWitt, Tyler, 1960; Соляник, 1975; DeWitt, Hureau, 1979; Eakin, 1990; Miller, 1993].
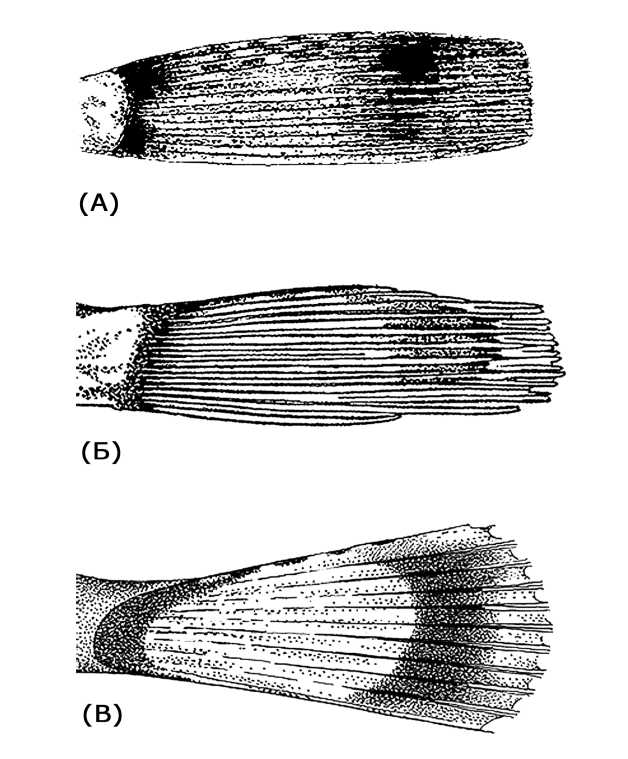
Рис. 2 . Хвостовой плавник D . longedorsalis :
А – по Roule, 1913; Б – по Miller, 1993; В – экземпляр ЗИН № 55578
Камуфляж истинного глаза достигается за счёт присутствия около него тёмных пятен или проходящих через глаз тёмных полос (это так называемый расчленяющий тип окраски). Маскирующее глаз тёмное пятно на щеке под орбитой присутствует у D. longedorsalis (см. рис. 1), его наличие отметил ещё Рул [Roule, 1913] в первоописании («Tête montrant une tache foncée sous l'orbite…», р. 17). Актуальность такой маскировки у длиннопёрой бородатки объяснима также очень крупными размерами глаз – ювенильным признаком, указывающим на возникновение рода путём педоморфоза. Как было ранее отмечено [Balushkin, 2000], этот путь эволюционного развития у бородатковых стал причиной самой масштабной среди подотряда
Notothenioidei миниатюризации его представителей, особо коснувшейся родов Artedidraco и Dol-loidraco . В русле стратегии маскировки истинного глаза можно рассматривать также скоординированное изменение размеров ложных и истинных глаз. В упомянутых выше экспериментах с рыбами-бабочками P. amboinensis [Lönnstedt, McCormick, Chivers, 2013] у молоди этого вида под прессом хищника статистически достоверно развиваются более крупные ложные глаза (они расположены у этого вида в задней части спинного плавника), а размер нормальных глаз уменьшается. У длиннопёрой бородатки Долло такой тип эпигенетического развития стал достаточно устойчивым (т.е. наследственным), так что глазные пятна у вида всегда крупнее нормальных глаз (см. рис. 1). Нельзя не отметить и ещё один важный эффект опытов с рыбами-бабочками: у находящейся под прессом хищника молоди этого вида продлевается срок существования глазных пятен (у взрослых рыб они исчезают совсем). По-моему, эти эксперименты довольно убедительно свидетельствуют о том, что морфологические преобразования происходят не в ходе неопределённого по времени ожидания подходящей генетической мутации и её дальнейшего отбора, а являются продуктом сознательных усилий самого организма по корректировке хода онтогенеза и последующего её закрепления в памяти с привлечением известных биохимических структур протоплазмы и ядра. То есть эволюция здесь идёт скорее «по Ламарку», а не «по Дарвину». Не развивая далее эту тему, оказавшуюся особенно болезненной для отечественной биологии, замечу, что уже с начала 90-х гг. прошлого века факт наследуемости приобретённых признаков был окончательно доказан с указанием конкретных молекулярно-генетических механизмов [см. обзор: Назаров, 2005].
Об эффективности защитной окраски у длиннопёрой бородатки Долло вполне может свидетельствовать тот факт, что в окраинных морях Антарктиды этот вид является одним из наиболее массовых среди представителей не только семейства Artedidraconidae , но и ряда других групп подотряда нототениевидных Notothenioidei [Ekau, 1990; Ekau, Gutt, 1991; Hubold, 1992; Gutt, Ekau, 1996; Eastman, Hubold, 1999]. Так, по результатам 6 немецких экспедиций в море Уэдделла (1979– 1989 гг.), численность этого вида составляла более половины (56%) от всех пойманных представителей семейства [Hubold, 1992], а согласно анализу результатов 10 тралений на судне « Nathaniel B. Palmer» в море Росса (летний сезон 1996–1997 гг.), численность длиннопёрой бородатки составляла 6.1% от численности всех пойманных рыб (это соответствовало 4-му месту по обилию после Pseudotrematomus scotti (Boulenger, 1907), Bathydraco marri Norman, 1938 и Pseudotrematomus eulepidotus (Regan, 1914)) [Eastman, Hubold, 1999] .
Длиннопёрые бородатки – обитатели мезобен-тальных глубин шельфа и верхнего склона Антарктиды, куда не проникают солнечные лучи видимого спектра. Поэтому всё, что было сказано выше о хитроумной стратегии этих рыб в использовании защитной окраски, базируется на допущении об её эффективном функционировании в условиях ультрафиолетового излучения (диапазон длины волны 380—10 нм), способного проникать на большие глубины (до 500–1500 м), биолюминесцентной подсветки, создаваемой многими обитателями больших глубин, и черенковского свечения.
О последнем факторе следует сказать особо. По мнению ряда физиков [Бабошин, Лопатников, Попов, 1986; Болотовский, 2009], его значение в жизни глубоководных рыб Мирового океана явно недооценивается биологами. Свечение воды на больших глубинах происходит в данном случае благодаря излучению света «быстрыми» электронами, имеющими скорость, превышающую фазовую скорость света (эффект Вавилова-Черенкова). Они образуются при распаде растворённых в океанской воде радиоактивных элементов, в первую очередь – изотопа калия-40. Считается, что хотя интенсивность такого свечения невелика, на больших глубинах всё же создаются световые условия (так называемые «сумерки Вавилова-Черенкова» [см. Болотовский, 2009]), в которых рыбы могут видеть друг друга. К этому нужно добавить, что длиннопёрые бородатки – донные рыбы, а у дна сумерки Вавилова-Черенкова становятся, как это не покажется странным, светлее. Связано это с тем, что в метровом слое воды, прилегающем непосредственно к океанскому дну, освещённость повышается за счёт гамма-излучения минералов дна. Последнее вызывает здесь образование дополнительных «быстрых» электронов, которые эффективно генерируют черенковские фотоны. Эффекты отпугивания и отклонения при демонстрации глазка могут, естественно, возникать только при наличии заметного контраста с основным фоном [Котт, 1950]. Видимо, по этой причине глазки длиннопёрых бородаток белого цвета, лучше заметного в сумеречных условиях глубин, а не тёмные, характерные для рыб, обитающих на небольших глубинах фотического слоя.
Альтернативной идеей, объясняющей наличие глазчатых пятен у D. longedorsalis, могла быть гипотеза филогенетического атавизма, согласно которой глазки на хвостовом плавнике рассматривались бы как рудиментарные образования, доставшиеся длиннопёрой бородатке от предка, обитавшего на небольших глубинах. Слабая изученность филогенетики и исторической биогеографии боро-датковых рыб пока не предоставляет нам сколь-либо серьёзных аргументов в её пользу. К настоящему моменту можно лишь констатировать, что у более примитивных представителей нототениевид- ных рыб (семейства Pseudaphritidae, Bovichtidae, Eleginopsidae, Nototheniidae, Harpagiferidae; подробнее о родственных связях семейств [см: Балуш-кин, 1992; Balushkin, 2000]), обитающих как на литорали, так и на больших глубинах, глазчатые пятна на хвостовом плавнике отсутствуют. Вместе с тем, создание при необходимости глазчатого пятна в своей окраске не представляет, похоже, для них особого труда. К этому способу пассивной защиты прибегает, на мой взгляд, антарктическая нототения Notothenia neglecta Nybelin, у молоди которой, судя по рисунку из монографии Нормана [Norman, 1938. Fig. 13 A], появляется глазчатое пятно на конце грудного плавника (рис 3). Рас- стояние между ложным и истинным глазом здесь не столь велико, как в случае с длиннопёрой боро-даткой, но его, по-видимому, оказывается вполне достаточно для дезориентации хищника.
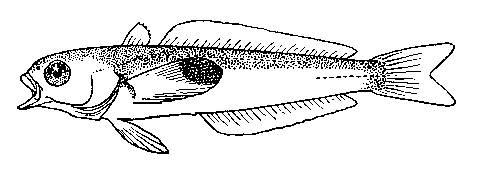
Рис. 3 . Пелагический малёк антарктической нототении Notothenia neglecta Nybelin с глазчатым пятном на конце грудного плавника [по Norman, 1938]
Труд по созданию глазка окупается и в этом случае сполна: антарктическая нототения – один из промысловых видов рыб в Южном океане. По мере роста рыб ложный глаз у N. neglecta исчезает точно так, как это происходит у рыб-бабочек P. amboinensis. Поскольку же мальки у нототении ведут пелагический образ жизни, обитая в 100метровом поверхностном слое воды, то и глазки у них формируются тёмного цвета.
Автор благодарен Маттиасу Штеманну (M. Stehmann), Хорсту Вилкенсу (H. Wilkens) и Гудрун Шульце (G. Schulze) за помощь в работе с коллекциями Зоологического музея Гамбургского университета. Автор признателен Е.А. Зиновьеву и Н.И. Литвиненко за критическое прочтение рукописи статьи, В.В. Сподаревой и М.В. Могановой за помощь в оформлении работы. Работа частично поддержана грантом Российского фонда фундаментальных исследований.
Список литературы О целесообразном характере окраски антарктической рыбы Dolloidraco longedorsalis (Artedidraconidae; Notothenioidei; Perciformes), живущей там, куда не проникает видимый солнечный свет
- Бабошин Ю.Б., Лопатников С.Л., Попов Н.И. Информативная значимость собственного черенковского свечения морской воды для глубоководных животных//Доклады АН СССР. 1986. Т. 290, № 4. С. 991-994
- Балушкин А.В. Классификация, родственные связи и происхождение семейств нототениоидных рыб подотряда Notothenioidei//Вопросы ихтиологии. 1992. Т. 32, вып. 2. C. 3-19.
- Балушкин А.В., Сподарева В.В. Морфологическая характеристика длиннопёрой бородатки Долло Dolloidraco longedorsalis (Artedidraconidae, Notothenioidei) из окраинных морей Антарктиды//Вопросы ихтиологии. 2015. Т. 55, № 5. С. 499508
- Болотовский Б. М. Излучение Вавилова -Черенкова: открытие и применение//Успехи физических наук. 2009. Т. 179, № 11. Р. 1161-1173
- Данилевский Н.Я. Дарвинизм: критическое исследование. СПб: Издание Меркурия Елеазарови-ча Комарова, 1885. Т. 1, ч. 2. 530 с
- Дольник В.Р. Непослушное дитя биосферы: Беседы о поведении человека в компании птиц, зверей и детей. СПб.: Петроглиф, 2009. 350 с
- Котт Х. Приспособительная окраска животных. М.: Иностранная литература, 1950. 544 с
- Мариковский П.И. Насекомые защищаются. М.: Наука, 1977. 200 с
- Назаров В.И. Эволюция не по Дарвину. Смена эволюционной модели. М.: КомКнига, 2005. 520 с
- Соляник Г.А. Донные и придонные рыбы Антарктики и сопредельных вод. Симферополь: Таврия, 1975. 182 с
- Сподарева В.В., Балушкин А.В. Описание нового вида жабовидной бородатки рода Pogonophryne (Perciformes: Artedidraconidae) из пролива Брансфилд (Антарктика) с ключом для определения видов группы «marmorata»//Вопросы ихтиологии. 2014. Т. 54, № 1. С. 3-8
- Шмальгаузен И.И. Проблемы дарвинизма. Л.: Наука, 1969. 494 с
- Balushkin A. V. Morphology, classification, and evolution of notothenioid fishes of the Southern Ocean (Notothenioidei, Perciformes)//Journal of Ichthyology. 2000. Vol. 40, suppl. 1. P. S74-S109
- Blest A.D. The function of eyespot patterns in the Lepidoptera//Behaviour. 1957. V. 11, № 2/3. P. 209-256
- Cott H. Adaptive Coloration in Animals. Oxford University Press, 1940. 373 p
- De Witt H.H., Hureau J.-C. Fishes collected during «Hero» cruise 72-2 in the Palmer Archipelago, Antarctica, with the description of two new genera and three new species.//Bull. Mus. natn. Hist. nat. Paris. 1979. Sect. A. Ser. 4. Vol. 1, № 3. P. 775-820
- DeWitt H.H., Tyler J.C. Fishes of the Stanford Antarctic Biological Research Program, 1958-1959//Stanford Ichthyol. Bull. 1960. Vol. 7, № 4. P. 162-199
- Eakin R.R. Artedidraconidae//Fishes of the Southern Ocean (O. Gon and P.C. Heemstra, eds.) J.L.B. Smith Institute of Ichthyology, Grahamstown, South Africa. 1990. P. 332-356
- Eakin R.R., Riginella E., La Mesa M. A new species of Artedidraco (Pisces: Artedidraconidae) from the Weddell Sea, Antarctica//Polar Biology. 2015. Vol. 38(10). P. 1597-1603
- Eastman J.T., Hubold G. The fish fauna of the Ross Sea, Antarctica//Antarct. Sci. 1999. Vol. 13, № 3. P. 293-304
- Ekau W. Demersal fish fauna of the Weddell Sea, Antarctica//Antarct. Sci. 1990. Vol. 2. № 2. P. 129137
- Ekau W., Gutt J. Notothenioid fishes from the Weddell Sea and their habitat, observed by underwater photography and television//Proc. NIPR Symp. Polar Biol. 1991. Vol. 4. P. 36-49
- Gutt J., Ekau W. Habitat partitioning of dominant high Antarctic demersal fish in the Weddell Sea and Lazarev Sea//J. Exp. Mar. Biol. Ecol. 1996. Vol. 206. P. 25-37
- Hubold G. Zur Okologie der Fische im Weddellmeer//Ber. Polarforsch. 1992. № 103. S. 1-157
- Lonnstedt O.M., McCormick M.I., Chivers D.P. Predator-induced changes in the growth of eyes and false eyespots//Science Reports. 2013. Vol. 3. P. 2259-2263
- Miller R.G. History and Atlas of the Fishes of the Antarctic Ocean. Foresta Institute for Ocean and Mountain Studies, Carson City, Nevada, 1993. 792 p
- Neudecker S. Eye camouflage and false eyespots: chaetodontid responses to predators//Environ. Biol. Fish. 1989. Vol. 25, № 1-3. P. 143-157
- Norman J.R. Coast fishes. Pt. III. The Antarctic zone//Discovery Reports. Cambridge. 1938. Vol. 18. P. 1-105
- Roule L. Description d'un genre nouveau et de deux espece nouvelle de poisons antarctique (Expedition Antarctique Francaise (1908-1910) comm. par M. J. B. Charcot)//Bull. Mus. Hist. Nat. 1913. Vol. 19, № 1. Р. 4-7
- Stevens M. The role of eyespots as anti-predator mechanisms, principally demonstrated in the Lepidoptera//Biological Reviews. 2005. Vol. 80, № 4. P. 573-588
- Waite E.R. Fishes//Australasian Antarct. Exp. 19111914. Sci. Repts. Ser. C. 1916. Vol. 3, Pt. 1. P. 1-92
