О корреляциях в анатомической структуре корневища и придаточных корней

Автор: Петухова Людмила Владимировна, Степанова Е.Н.
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 2, 2016 года.
Бесплатный доступ
Показаны корреляции в анатомической структуре корневища и придаточных корней, формирующихся на нем. Слабая камбиальная активность корневища и первичный рост усиления (за счет разрастания апекса) соответствуют слабой активности камбия в корнях, что приводит к сохранению в них первичной структуры.
Придаточные корни, корневище, камбий, первичный рост усиления, анатомическая структура
Короткий адрес: https://sciup.org/146116644
IDR: 146116644 | УДК: 581.84
On the correlations in the anatomical structure of rhizomes and adventitious roots
We showed the correlations in the anatomical structure of rhizomes and their adventitious roots. Weak cambial activity of rhizome and primary growth (due to the expansion of the apex) correspond the weak activity of cambium in the roots. This leads to the preservation of their primary structure.
Текст научной статьи О корреляциях в анатомической структуре корневища и придаточных корней
В любом организме, в том числе и в растительном, существует сложная система связей, обеспечивающих согласованное функционирование отдельных его частей. Корнелистовая функциональная связь показана еще в середине прошлого века нашими выдающимися физиологами (Сабинин, 1949; Курсанов, 1959; Казарян, 1969). '
Физиологические связи подтверждаются и морфологическими признаками, что показано нами на примере моноподиально-розеточных растений, у которых четко прослеживается зависимость между формированием листьев и заложением придаточных корней (Петухова, 1994). Оказалось, что корнелистовое отношение для моноподиально-розеточных растений может служить одним из возрастных признаков.
Четкая морфофизиологическая связь прослеживается в соотношении лист - пазушная почка. Лист, в котором хорошо выражены ростовые процессы, тормозит формирование и развертывание почки, что хорошо демонстрируется на примере гравилата, рдеста гребенчатого и других видов (Петухова, 1994; Петухова, Лапиров, 1983).
Наши наблюдения показали, что корреляции прослеживаются не только морфологические или морфофизиологические. В анатомической структуре отдельных органов растения существуют четкие взаимосвязи, и прежде всего, в анатомических особенностях корневища и придаточных корней, на нем возникающих.
В типичном случае корни двудольных растений претерпевают вторичные преобразования за счет заложения и работы камбия. Последний возникает из прокамбиальных клеток, которые остались недифференцированными, прежде всего, под флоэмой и позднее - над ксилемой в зоне перицикла. Раньше начинает работу камбий, заложившийся под флоэмой. Он образует вторичные проводящие элементы ксилемы и флоэмы. Камбий, возникающий в перицикле, дает паренхиму радиальных лучей. Они начинаются от протоксилемы и называются первичными. По мере увеличения диаметра корня радиальные лучи закладываются в других участках вторичных проводящих тканей и называются вторичными. Ширина первичных и вторичных лучей в корнях разных растений отличается, но, как правило, первичные лучи определяются достаточно легко, отличаясь большей шириной (рис. 1). У многолетних корней часто заметны годичные кольца. При вторичных преобразованиях, как правило, первичная кора слущивается. Эти изменения в структуре корня детально описываются в обширной учебной литературе и считаются, по сути, классическими.
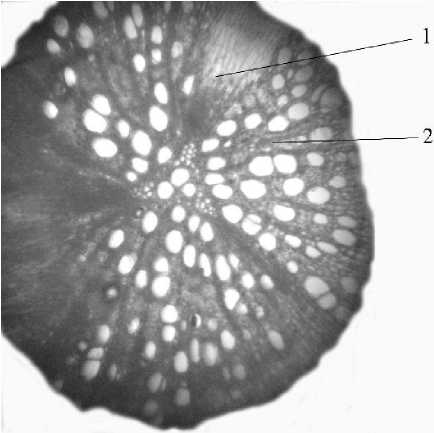
Рис. 1 . Ксилемная зона придаточного корня малины (Rubus idaeus L.): 1 - первичный радиальный луч, 2 - вторичный радиальный луч
Однако, при описании вторичных преобразований, как правило, не указывается, о каких корнях, главном или придаточных, идет речь. Наши наблюдения показали (Петухова, Степанова, 2015), что в пределах одной корневой системы главный и придаточные корни различаются по анатомическим особенностям. Эти отличия касаются и особенностей утолщения. В системе главного корня чаще всего наблюдаются типичные вторичные преобразования, в то время как в придаточных корнях наблюдаются разные варианты. Причем, следует отметить, что существует корреляция между структурой корневища и придаточными корнями, на нем возникающими.
Для моноподиально-розеточных короткокорневищных и кистекорневых растений с эпигеогенным корневищем характерен первичный рост усиления, который обеспечивает утолщение диаметра каждого последующего годичного прироста корневища. Камбиальная деятельность в нем невелика и не приводит к серьезным изменениям в структуре. Примерами могут служить манжетка, горец змеиный, подорожник, буквица и другие.
Рассмотрим особенности анатомической структуры корневища манжетки. Конус нарастания у нее выпуклый, в отличие от ряда других розеточных растений (Василевская и др., 1973). Увеличение диаметра наблюдается на некотором удалении от апекса (рис. 2).
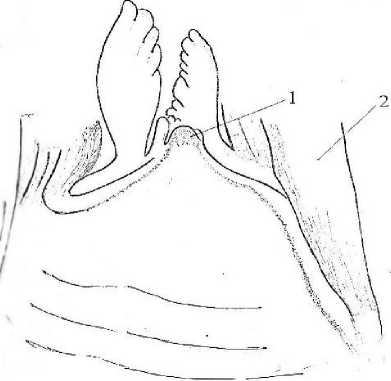
Рис. 2 . Продольный срез через апикальную часть взрослого растения манжетки (Alchemilla L.): 1 - конус нарастания, 2 - основания листьев
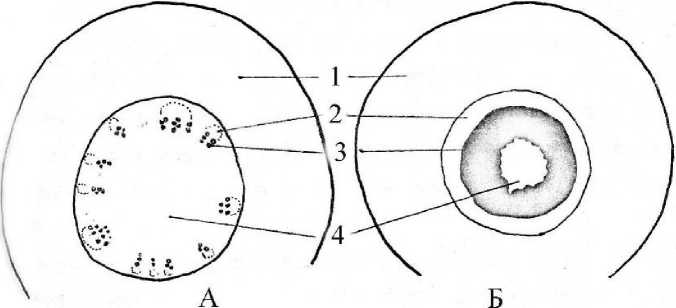
Рис. 3 . Схема строения корневища ювенильного растения манжетки (Alchemilla L.) в апикальной (А) и базальной (Б) частях:
1 - первичная кора, 2 - флоэма, 3 - ксилема, 4 - сердцевина.
У ювенильных растений корневище близ апекса имеет пучковое строение. За однослойной эпидермой следует довольно мощная первичная кора, составляющая примерно половину диаметра стебля. Сердцевина небольшая, 10 — 15 клеток по диаметру. Наблюдается образование сложных синтетических пучков и интенсивное одревеснение в зоне ксилемы, что приводит к образованию одревесневшего кольца в основании побега (рис. ЗА, Б).
В последующих годичных приростах корневища изменяется соотношение тканей. Постепенно разрастается сердцевина, достигая 75% диаметра всего корневища у взрослых генеративных растений, что составляет 100 - 120 клеток по диаметру. На долю коровой части приходится 15 - 17%. Увеличение сердцевины достигается за счет усиления апекса и благодаря делению клеток перимедуллярной зоны, которая у каждого последующего годичного прироста выражена отчетливее. Разрастание сердцевины обеспечивает первичное медуллярное утолщение корневища или «рост усиления», в результате чего каждый последующий годичный прирост чуть толще предыдущего. С возрастом пучки становятся синтетическими, однако между ними наблюдаются широкие паренхимные лучи камбиального происхождения, что объясняется подавлением здесь васкулярной деятельности камбия в связи с заложением почек или формированием цветоносных побегов (рис. 4). Сходная картина наблюдается и у других растений (Михайловская, Федотова, 1976).
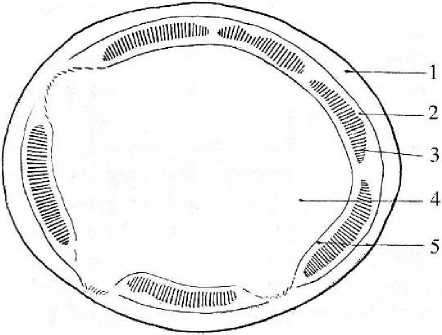
Рис. 4 . Схема строения корневища средневозрастного растения манжетки (Alchemilla L.): 1 - первичная кора, 2 - флоэма, 3 - ксилема, 4 - сердцевина; перимедуллярная зона.
В соответствии со слабой активностью камбия в корневище его деятельность не оказывает заметного влияния на диаметр.
Уже в конце первого года жизни в перицикле закладывается феллоген, образующий несколько слоев феллоидной ткани, стенки которой совершенно лишены опробковения. Интересно отметить, что феллоген может закладываться неоднократно, однако феллоидная ткань никогда не выходит на поверхность, и первичная кора сохраняется в течение всей жизни годичного прироста.
Со слабой камбиальной активностью в корневище коррелирует слабая активность камбия в корнях. Наши наблюдения показали, что она незначительна как в главном, так и в придаточных корнях. Главный корень в первичной структуре диархный. Вторичное утолщение слабое, поэтому первичная кора сохраняется, центральный цилиндр узкий. Ксилемная и лучевая паренхима одревесневают. Придаточные корни в первичной структуре трех-/четырехархные в зависимости от диаметра; в центре образуется небольшое количество клеток центральной паренхимы. Как и в большинстве придаточных корней, перицикл однослоен (рис. 5). Первичная кора довольно мощная, с системой мелких межклетников, в три раза превышает радиус центрального цилиндра. Экзодерма однорядная, эндодерма отчетливо выражена.
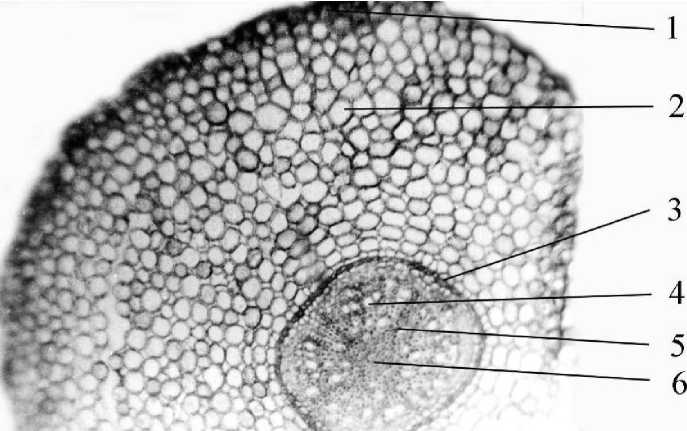
Рис. 5 . Поперечный срез придаточного корня манжетки (Alchemilla L.) первого года жизни: 1 - экзодерма, 2 - мезодерма, 3 - эндодерма, 4 - вторичная ксилема, 5 - первичная ксилема, 6 - центральная паренхима.
При вторичных преобразованиях наблюдается одревеснение центральной паренхимы (Петухова, Степанова, 2015) и паренхимы в ксилемной зоне, проводящих элементов вторичной ксилемы образуется немного, вторичная флоэма хорошо заметна. Поскольку вторичный рост незначителен, первичная кора в полностью сформированном корне сохраняется. Экзодерма становится 2-3-слойной, с относительно тонкими стенками. С возрастом клетки коры буреют, периферические постепенно разрушаются, однако первичная кора сохраняется в течение всей жизни корня. Во второй — третий год жизни в перицикле закладывается феллоген, дающий несколько слоев полидермы (т.е. слоев опробковевших и неопробковевших клеток) (Эсау, 1969), причем, как и в корневище, феллоген может закладываться неоднократно, особенно в местах повреждения первичной коры, однако поверхности полидерма не достигает. Таким образом, у манжетки четко прослеживается корреляция между структурой корневища и придаточными корнями, возникающими на нем, что отражается в слабой камбиальной активности, в работе феллогена, не образующего типичной перидермы.
Такие же закономерности в анатомической структуре корневища и придаточных корней наблюдаются у горца змеиного. Диаметр каждого последующего прироста корневища больше предыдущего, поскольку, так же как и у манжетки; он обеспечивается первичным ростом усиления. Однако у горца в равной степени выражен как медуллярный, так и кортикальный рост, поэтому каждый последующий годичный прирост корневища отличается не только более широкой сердцевиной, но и более мощной коровой частью. Деятельность камбия почти незаметна, поэтому проводящие пучки более изолированы по сравнению с манжеткой. В придаточных корнях деятельность камбия не приводит к существенным перестройкам в первичной структуре, сосудов вторичной ксилемы мало (по 3 - 4 в каждой группе) и по размерам они уступают метаксилеме. В наиболее толстых корнях число архов в первичной структуре насчитывается до 5. Первичная кора сохраняется до конца жизни корня.
У других моноподиально-розеточных растений, например, у гравилата городского, наряду с первичным ростом усиления более активна и деятельность камбия в корневище, в результате проводящие пучки в базальной части корневища молодых растений расположены тесно, могут сливаться. В корневище камбий работает в течение всей его жизни, однако наряду с проводящими элементами производит широкие паренхимные участки, которые прерывают вторичную ксилему. Наряду с этим развитие цветоносов приводит к одревеснению участков этой паренхимы, образуются довольно широкие одревесневшие полосы. Следует отметить, что камбий работает, не образуя четких годичных колец. С возрастом первичная кора отмирает, с самых первых годичных приростов слущивается, на последующих приростах отмершие клетки коры сохраняются до конца жизни.
Заложившийся в перицикле феллоген образует полидерму, которая не всегда выходит на поверхность. Закладывается феллоген в первый год жизни корневища, с возрастом увеличивается число слоев клеток полидермы (т.е. чередующихся опробковевших и неопробковевших слоев). Таким образом, в корневище гравилата деятельность камбия более заметна по сравнению с манжеткой, что коррелирует с его активностью в корнях. И в главном, и в придаточных корнях первичная структура с возрастом преобразуется во вторичную за счет работы камбия. Соответственно, это приводит к слущиванию первичной коры, корни имеют типичную вторичную структуру.
А) Б)
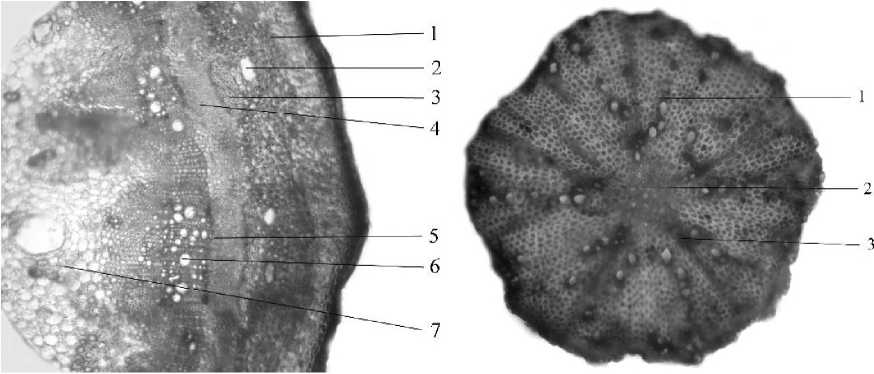
Рис. 6. Поперечный срез корневища (А) и ксилемная зона придаточного корня (Б) золотарника канадского (Solidago canadensis L.): А) 1 - первичная кора, 2 - млечный ход, 3 - флоэмная склеренхима, 4 - зона вторичной флоэмы, 5 - камбиальная зона, 6 - вторичная ксилема, 7 - сердцевина; Б)1 - вторичная ксилема, 2 - центральная паренхима, 3 - первичная ксилема.
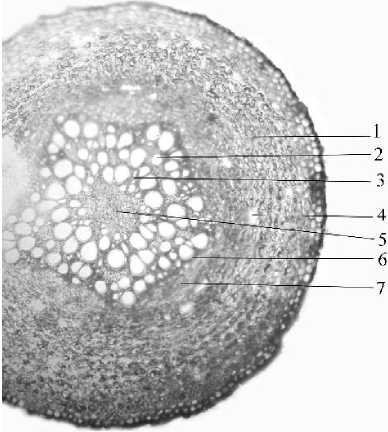
Рис. 7 . Поперечный срез придаточногго корня посконника пятнистого (Eupatorium maculatum L.): 1 - вторичная кора, 2 - вторичная ксилема, 3 - первичная ксилема, 4 - млечный ход, 5 - центральная паренхима, 6 - камбиальная зона, 7 - вторичная флоэма.
У трав с гипогеогенным корневищем надземные побеги монокарпические, чаще и моноциклические, должны быть механически прочными, поэтому у двудольных растений камбий активен, образуется довольно много вторичной ксилемы, обеспечивающей в основном прочность стебля. С возрастом подвергается одревеснению паренхима сердцевинных лучей в области вторичной ксилемы, что приводит к формированию сплошного кольца одревеснения. В корневищах сердцевина хорошо выражена, однако первичный рост усиления отсутствует; с возрастом диаметр сердцевины не увеличивается Придаточные корни, формирующиеся на корневище в этом случае, имеют типичное вторичное строение. Это соответствие можно показать на многих примерах (рис. 6, 7).
Петухова Л.В. О корреляциях в анатомической структуре корневища и придаточных корней / Л.В. Петухова, Е.Н. Степанова // Вести. ТвГУ. Сер.: Биология и экология. 2016. № 2. С. 44-52.
Список литературы О корреляциях в анатомической структуре корневища и придаточных корней
- Василевская В.К., Баранов М.П., Борисовская Г.М. 1973. Строение розеточного растения Plantago major L. в первый год жизни//Бот. журн. Т. 58. № 1. С. 33-42.
- Казарян В.О. 1969. Старение высших растений. М.: Наука. 280 с.
- Курсанов А.Л. 1959. Корневая система растений как орган обмена веществ//Изв. АН СССР. Сер. биол. № 6. С. 689-705.
- Михайловская И.С., Федотова Т.А. 1976. Анатомическая структура корневища Potentilla alba в онтогенезе//Бюлл. МОИП. Отд. биол. Т. 81. Вып. 2. С. 99-107.
- Петухова Л.В. 1994. К вопросу о корреляциях в теле растений//Флора и растительность Тверской области. Тверь. С. 68 -75.
- Петухова Л.В., Лапиров А.Г. 1983. Некоторые биоморфологические особенности рдеста гребенчатого (Potamogeton pectinatus)//Взаимоотношения компонентов биоценозов в южнотаежных ландшафтах. Калинин. С. 77-83.
- Петухова Е.Н., Степанова Е.Н. 2015. Разнообразие анатомической структуры корней в пределах корневой системы//Вестн. ТвГУ. Сер.: Биология и экология. № 4. С. 167-172.
- Сабинин Д.А. 1949. О значении корневой системы в жизнедеятельности растений//Тимирязевские чтения. Вып. 9. С. 1-48.
- Эсау К. 1980. Анатомия семенных растений. М.: Мир. Т. 2. 558 с.
