О лесообразующей роли темнохвойных пород в районах широколиственно-темнохвойных и горнотаежных темнохвойных лесов Южного Урала

Автор: Горичев Ю.П., Давыдычев А.Н., Кулагин А.Ю.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 1 т.27, 2018 года.
Бесплатный доступ
В статье на основе анализа и обобщения результатов исследований сделаны выводы об участии темнохвойных пород ели сибирской и пихты сибирской в лесообразовательном процессе в районах широколиственно-темнохвойных и темнохвойных лесов Южного Урала
Южный урал, лесообразовательный процесс, широколиственно-темнохвойные леса, темнохвойные породы, ель сибирская, пихта сибирская
Короткий адрес: https://sciup.org/148313581
IDR: 148313581 | УДК: 574.38; | DOI: 10.24411/2073-1035-2018-10007
Текст краткого сообщения О лесообразующей роли темнохвойных пород в районах широколиственно-темнохвойных и горнотаежных темнохвойных лесов Южного Урала
Темнохвойные породы ель сибирская Picea obovata Ledeb. и пихта сибирская Abies sibirica Ledeb. участвуют в лесообразовательном процессе в двух ботанико-географических районах Южного Урала – районе горнотаежных темнохвойных лесов и районе широколиственнотемнохвойных лесов.
Стационарные и маршрутные исследования лесных экосистем проведенные нами в данных районах (Горичев и др., 2007, 2012; Давыдычев и др., 2011) позволяют сделать некоторые выводы об участии темнохвойных пород в лесообразовательном процессе. Определены экото-пические ареалы пород. Проанализированы показатели фитоценотической активности пород –
плотность древостоя, запаса древесины, плотность подроста по 5 пробным площадям (ПП) в районе горнотаежных темнохвойных лесов и по 10 ПП - в районе широколиственнотемнохвойных лесов.
Многими исследователями отмечается, что экологические и фитоценотические свойства ели сибирской и пихты сибирской во многом схожи. При этом отмечаются определенные различия. В частности, отмечается требовательность ели к условиям грунтового увлажнения (Порфирьев, 1947; Поликарпов и др., 1986), а пихты к условиям атмосферного увлажнения (Кулагин, 1978; Крылов и др., 1986).
Как показали наши исследования, активное участие темнохвойных пород в лесообразовательном процессе наблюдается в районе горнотаежных темнохвойных лесов. Лесорастительные условия района в полной мере благоприятны для пород. Темнохвойные породы являются эдификаторами коренных насаждений, где абсолютно доминируют. Экотопические ареалы ели и пихты охватывают практически весь спектр лесорастительных условий (ЛРУ), за исключением экстремальных (высокогорья и болота). В избыточно увлажненных экотопах и в высокогорьях фитоценотические позиции пихты, по сравнению с елью, более ослаблены. В данном районе исследованы 5 темнохвойных насаждений II-III класса бонитета, из которых 3 насаждения – коренные, 2 насаждения - условно-коренные. Все насаждения отнесены к одному типу – пихто-ельник травяной. В состав древостоя условно-коренных насаждений, занимающих инсолируемые покатые и крутые склоны, входят сосна обыкновенная и лиственница Сукачева. Темнохвойные породы абсолютно доминируют в древесном ярусе (8-10 единиц). Участие ели в составе древостоя составляет от 2 до 6 единиц, запас древесины -85-277 м3/га. Участие пихты в данных насажде- ниях от 3 до 8 единиц, запас древесины - 108279 м3/га. В исследованных насаждениях пихта превосходит ель по плотности древостоя (числу деревьев на 1 га) в 1,3-6,0 раз. Плотность деревьев ели составляет 100-368 шт./га, пихты – 468-924 шт./га. По запасу древесины пихта также превосходит ель в большинстве насаждений (рис. 1). Темнохвойные породы активно участвуют в возобновлении. Во всех насаждениях крупный подрост пихты более многочислен, чем, ели - соответственно 0,3-2,2 тыс. шт/.га и 0,1-0,3 тыс. шт./га (рис. 2). В 3 насаждениях (ПП-15, ПП-16, ПП-17) мелкого подроста пихты также больше, чем ели - соответственно 2,4-4,4 тыс. шт./га и 0,8-1,2 тыс. шт./га. В 2 насаждениях (ПП-18, ПП-19) мелкий подрост ели более многочислен – соответственно 3,2 тыс. шт./га и 1,2-2,0 тыс. шт./га.
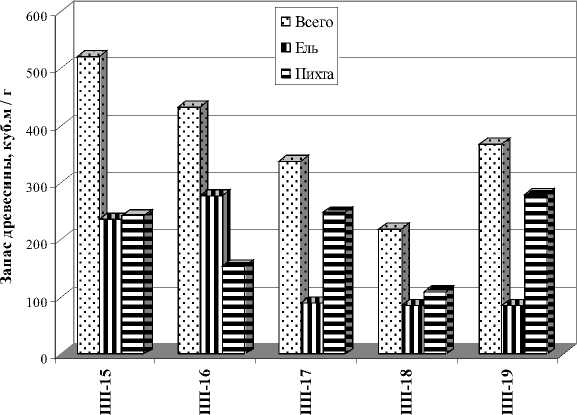
Рис. 1. Запас древостоя ели и пихты в горнотаежных темнохвойных насаждениях
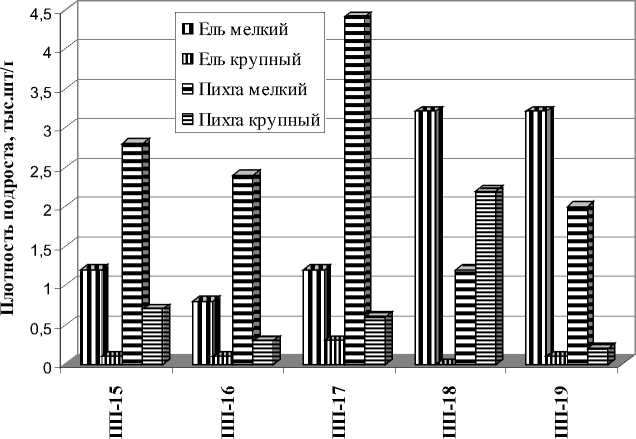
Рис. 2. Плотность подроста ели и пихты в горнотаежных темнохвойных насаждениях
В районе широколиственно-темнохвойных лесов по сравнению с районом горнотаежных темнохвойных лесов роль темнохвойных пород в лесообразовательном процессе ослабевает. Это, прежде всего, проявляется в сужении эко-топических ареалов пород. Темнохвойные породы в районе широколиственнотемнохвойных лесов доминируют в формируемых ими темнохвойных насаждениях, занимающих холодные и контрастно-холодные клима-топы. Они также входят в состав смешанных широколиственно-темнохвойных насаждений, занимающих прохладные климатопы. Экотопи-ческий ареал ели уже, чем пихты. Ареал доминирования ели охватывает экотопы со свежими и влажными ЛРУ – днища долин горных рек, ручьев и временных водотоков (где формируются т.н. приручьевые типы леса), а также пологие и вогнутые склоны, нижние части длинных склонов, куда поступает дополнительное увлажнение. Вне экотопического ареала ели находятся теплые климатопы, занимаемые широколиственными насаждениями. Экотопиче-ский ареал пихты несколько шире, чем у ели. Наибольшая активность пихты, как лесообра-зователя, наблюдается на склонах северных экспозиций со свежими ЛРУ. Пихта входит в состав некоторых типов широколиственных насаждений (кленовников).
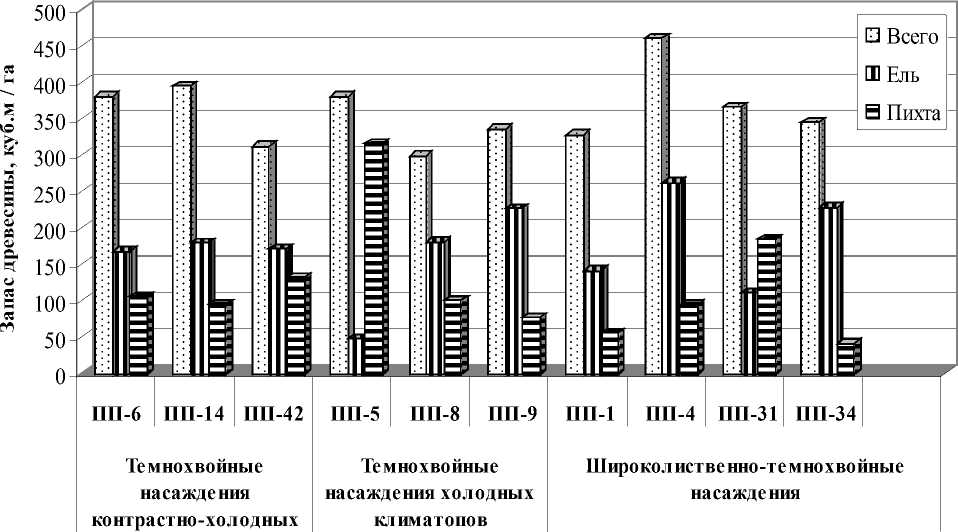
Рис. 3. Запас древостоя ели и пихты в темнохвойных и широколиственно-темнохвойных насаждениях в районе широколиственно-темнохвойных лесов
Ослабление лесообразующией роли темнохвойных пород в районе широколиственнотемнохвойных лесов можно объяснить ухудшением лесорастительных условий, и прежде всего, условий увлажнения. В районе широколиственно-темнохвойных лесов по сравнению с районом горнотаежных темнохвойных лесов количество осадков снижается с 800 до 670 мм, при некотором увеличении показателя испаряемости. Обе породы испытывают недостаток увлажнения. Режим и условия увлажнения выступают основным лимитирующим фактором, негативно сказывающимся на фитоценотиче-скую активность обеих пород и ограничивающим их экотопические ареалы. В данном районе в лесообразовательном процессе активно участвуют широколиственные породы, создающие конкуренцию темнохвойным породам.
В исследованных коренных и условнокоренных темнохвойных насаждениях холодных климатопов I-III класса бонитета участие темнохвойных пород в составе древостоя составляет 9-10 единиц. Пихта превосходит ель по плотности древостоя в 1,9-3,2 раза. Плотность деревьев пихты составляет 512-672 шт./га, ели – 192-264 шт./га, в условнокоренном насаждении превышение составляет 10,8. По запасу древесины в коренных насаждениях ель превосходит пихту в 1,8-2,9 раз. Участие ели составляет от 1 до 7 единиц, запас древесины 48-227 м3/га, участие пихты в данных насаждениях от 2 до 8 единиц, запас древе- сины 77-315 м3/га (рис. 3). Плотность крупного подроста ели и пихты примерно равная, соответственно 0,1-1,1 тыс. шт./га и 0,3-0,9 тыс.
шт./га (рис. 4). Плотность мелкого подроста ели и пихты соответственно 0,2-2,2 тыс. шт./га и 0,8-1,4 тыс. шт./га.
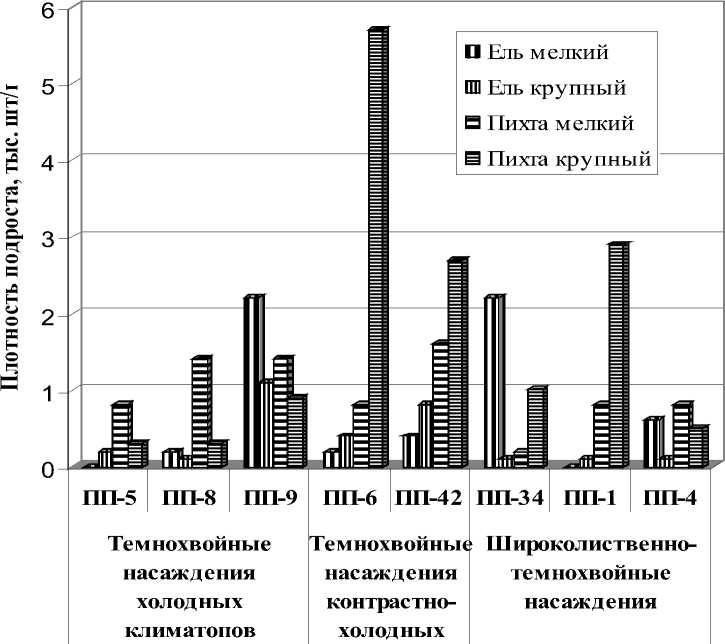
Рис. 4. Плотность подроста ели и пихты в темнохвойных и широколиственно-темнохвойных насаждениях в районе широколиственно-темнохвойных лесов
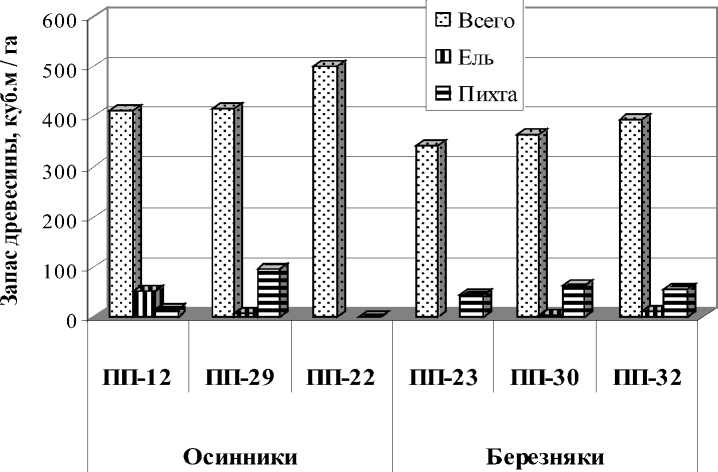
Рис. 5. Запас древостоя ели и пихты в производных насаждениях в районе широколиственно-темнохвойных лесов
В исследованных темнохвойных насаждениях контрастно-холодных климатопов II класса бонитета участие темнохвойных пород в составе древостоя коренных насаждений достигает 10 единиц, условно-коренных насаждений – 7 единиц. Участие ели в составе древостоя составляет от 4 до 6 единиц, запас древесины 167180 м3/га. Участие пихты в данных насаждениях от 2 до 4 единиц, запас древесины 96-133 м3/га. Плотность деревьев ели в коренном насаждении 196 шт./га, пихты – 368 шт./га, в условно-коренном насаждении (ПП-14) - соответственно 360 и 620 шт./га. Пихта превышает ель по плотности древостоя в коренном насаждении в 1,7-1,9 раза, в условно-коренном насаждении (ПП-6) превышение составляет 3,2. По запасу древесины ель превосходит пихту в коренном насаждении в 1,9 раз, в условнокоренных насаждениях – 1,7-3,2 раза. Плотность крупного подроста пихты значительно выше, чем ели - соответственно 2,7-5,7 тыс. шт./га и 0,2-0,4 тыс. шт./га. Плотность мелкого подроста пихты ели также выше, чем ели - соответственно 0,8-1,6 тыс. шт./га и 0,2-0,4 тыс. шт./га.
В исследованных широколиственнотемнохвойных насаждениях II класса бонитета, занимающих прохладные климатопы, участие темнохвойных пород в составе древостоя достигает 8 единиц, в условно-коренных насаждений – 6-8 единиц. Участие ели в составе древостоя в коренных насаждений составляет 6-7 единиц, условно-коренных насаждений – 3-4 единицы, запас древесины соответственно 227-262 и 111-141 м3/га. Участие пихты в данных насаждениях составляет соответственно 12 и 2-5 ед., запас древесины 42-94 и 56-184 м3/га. Плотность деревьев ели в коренных и условно-коренных насаждениях 108-172 шт./га, плотность деревьев пихты в коренных и насаждениях – 372-452 шт./га, в условно-коренных насаждениях она выше – 448-832 шт./га. Пихта превышает ель по плотности древостоя в коренных насаждениях в 2,2-4,2 раза, в условнокоренных насаждениях превышение составляет 3,4-7,7 раза. По запасу древесины ель превосходит пихту в коренных насаждении в 2,8-5,5 раз, в условно-коренном насаждении (ПП-31) запас древесины ели в 1,7 раза меньше, чем пихты. Плотность крупного подроста пихты во всех исследованных насаждениях значительно выше, чем ели – соответственно 0,5-2,9 тыс. шт./га и 0,1 тыс. шт./га. Плотность мелкого подроста ели варьирует в значительных пределах – от 0,6 до 2,2 тыс. шт./га, плотность пихты более стабильна - 0,2-0,8 тыс. шт./га. В услов- но-коренном насаждении отмечен «взрыв возобновления» пихты до 16 тыс. шт./га.
В исследованных производных насаждениях , возникших на месте коренных широколиственно-темнохвойных насаждениях после сплошных рубок – осинниках и березняках участие темнохвойных пород в составе древостоя достигает 2-3 единиц. Участие ели в составе древостоя в осинниках достигает 1 единицы, запас древесины от 7 до 51 м3/га, в березняках участие ели менее единицы, запас древесины от 5 до 13 м3/га (рис. 5). Участие пихты в осинниках и березняках достигает 2 единиц, запас древесины составляет соответственно - от 16 до 95 м3/га и от 44 до 63 м3/га. В 2 насаждениях – осиннике (ПП-22) и березняке (ПП-23) ель в составе древостоя отсутствует. По плотности деревьев, как в осинниках, так и в березняках (256-332 шт./га и 404-428 шт./га), пихта многократно превосходит ель (соответственно 8-96 шт./га и 4-12 шт./га). В насаждениях, где отсутствует ель, пихта единично встречается в осиннике (4 шт./га), в березняке ее больше (88 шт./га). Редкий подрост ели встречен лишь в 2 насаждениях – березняках. В одном из них (ПП-32) учтен крупный подрост (0,1 тыс. шт./га), в другом (ПП-30) – мелкий (0,2 тыс. шт./га). Во всех насаждениях встречен подрост пихты, плотность крупного подроста составляет от 0,1 до 2,5 тыс. шт./га, мелкого – от 0,1 до 1,2 тыс. шт./га. В березняке отмечен взрыв возобновления (9,2 тыс. шт./га).
В широколиственных насаждениях , занимающих теплые климатопы ель отсутствует. Пихта входит в состав древостоя некоторых типов насаждений (кленовники) (до 1 единицы), запас древесины 10-32 м3/га. Встречен редкий крупный подрост (до 0,1 тыс. шт./га).
ВЫВОДЫ
Как показали исследования, более активное участие темнохвойных пород в лесообразовательном процессе наблюдается в районе горнотаежных темнохвойных лесов. Экотопические ареалы ели и пихты охватывают практически весь спектр лесорастительных условий. Темнохвойные породы являются эдификаторами коренных насаждений, доминируют в древостое, активно участвуют в возобновлении. В районе широколиственно-темнохвойных лесов роль темнохвойных пород в лесообразовательном процессе ослабевает. Сужаются экотопические ареалы пород. Темнохвойные породы доминируют в темнохвойных насаждениях, занимающих холодные и контрастно-холодные клима-топы. В смешанных широколиственнотемнохвойных насаждениях, занимающих про- хладные климатопы в состав древостоя, наряду с темнохвойными, входят широколиственные породы, оказывающие определенную конкуренцию. Возобновление темнохвойных в этих насаждениях затруднено. Экотопический ареал ели занимает узкий спектр лесорастительных условий. Ареал пихты несколько шире, пихта входит в состав некоторых типов широколиственных насаждений (кленовников), где ель отсутствует.
Список литературы О лесообразующей роли темнохвойных пород в районах широколиственно-темнохвойных и горнотаежных темнохвойных лесов Южного Урала
- Горичев Ю.П., Давыдычев А.Н., Кулагин А.Ю., Алибаев Ф.Х., Юсупов И.Р. Горнотаежные темнохвойные леса Южно-Уральского заповедника, состояние и особенности возобновления // Вестн. Оренбург. гос. ун-та. Спец. вып. (75). 2007. С. 84-87.
- Горичев Ю.П., Давыдычев А.Н., Алибаев Ф.Х., Кулагин А.Ю. Широколиственно-темнохвойные леса Южного Урала: пространственная дифференциация, фитоценотические особенности и естественное возобновление. Уфа: Гилем, 2012. 176 с.
- Давыдычев А.Н., Горичев Ю.П., Кулагин А.Ю., Сулейманов Р.Р. Лесовозобновительные процессы под пологом древостоя в широколиственно-темнохвойных лесах Южного Урала // Лесоведение. 2011. №1. С. 39-48.
- Крылов Г.В., Марадудин И.И., Михеева Н.И., Козакова Н.Ф. Пихта. М.: Агропромиздат, 1986. 239 с.
- Кулагин Ю.З. Хвойные лесообразователи и экологическое прогнозирование // Экология хвойных / БФАН СССР. Уфа, 1978. С. 5-21.
- Поликарпов Н.П., Чебакова Н.М., Назимова Д.И. Климат и горные леса Южной Сибири. Новосибирск: Наука, 1986. 226 с.
- Порфирьев В.С. К познанию пихтовых ценозов Татарской АССР // Учен. зап. Казан. пед. ин-та. 1947. Вып. 5. С. 17-55.
