О некоторых аспектах физиолого-биохимических процессов, обусловливающих изменчивость признаков окраски меха европейской рыжей полевки

Автор: Емельянова А.А.
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Зоология
Статья в выпуске: 5, 2007 года.
Бесплатный доступ
На примере рыжих полевок, отловленных на территории Тверской области, показано, что с понижением среднесуточной температуры и увеличением количества осадков в мехе зверьков увеличивается содержание рыжих тонов на фоне общего посветления окраски. Представляется вероятной связь этого явления с температурной обусловленностью активности фермента тирозиназы и особенностями распределения эу- и феомеланина в волосе полевок. Демонстрируется неоднозначность в проявлении признаков окраски меха, обусловленная влиянием внешних и внутренних причин.
Короткий адрес: https://sciup.org/146116093
IDR: 146116093 | УДК: 591.157:599.323.43
On some aspects of physiologico-biochemical processes determining the variability of the fur coloration in European
The increase of red tint along with whitening of the general fur coloring under decreasing of average daily temperature has been shown on a series of Bank Voles, captured in Tver region. The phenomenon appears to be connected with the temperature-dependent activity of tyrosinase as well as with distribution of eu- and phaeomelanin in the hair of the vole. The ambiguity in the appearance of fur coloring characters dependant on external and internal causes is demonstrated.
Текст научной статьи О некоторых аспектах физиолого-биохимических процессов, обусловливающих изменчивость признаков окраски меха европейской рыжей полевки
Место, время сбора материала, его объем и методика обработки приведены по ходу изложения, в табл. 1, также более ранних работах, посвященных изменчивости окраски рыжей полевки и методам ее исследования [6; 7; 9].
Окраска рыжей полевки носит название агути (agouti). Эта окраска свойственна многим диким животным и обусловлена зонарным распределением пигмента в волосе. Меланин в окрашенных структурах существует в двух формах: в форме эумеланина (коричневый или черный пигмент) и в форме феомеланина (желтый или красноватый пигмент). Волос агути имеет терминальную или субтерминальную желтую феомела-ниновую полосу, а в остальных его частях находится эумеланиновый пигмент (у домовой мыши с типичной окраской агути в зоне желтого кольца зачастую образуется смесь эу- и феомеланина) [16]. В частности, у рыжей полевки нижняя часть волоса темная, а верхняя имеет ржаво-коричневые тона различной интенсивности.
Известно, что окраска животных – признак вариабельный, для которого, как и для всех признаков полигенной природы, характерно значительное варьирование под воздействием условий среды. Изменения окраски животных тесно связаны с определенными эколого-географическими условиями. В жизни мелких млекопитающих, в частности грызунов, особую роль играют такие экологические факторы, как температура и влажность окружающей среды, что объясняется несовершенством терморегуля- торного механизма. Температура тела полевок особенно зависима от температуры среды обитания. С этим связано их активное приспособление к микроклиматическим условиям местообитания, проявляющееся в выборе места для устройства нор и гнёзд, и сложность этих убежищ [2; 10; 11; 12].
Данное теоретическое положение было подробно рассмотрено нами на примере 190 молодых рыжих полевок, отловленных в летние сезоны 1999 и 2000 гг. на территории пяти административных районов Тверской области. Сеголетки принадлежали к двум условным возрастным группам, которые имели разное происхождение в зависимости от времени рождения. Группа subadultus (sb) включала при возможности выделяемые подгруппы sb3 и sb2, т.е сеголеток, относящихся к раннелетней и летней сезонным генерациям (когорты К 2 и К 3 соответственно). Abulescens (ab) объединяла весеннюю группу сеголеток – особей первых пометов перезимовавших животных (когорта К 1 ). Путем сопоставления значений цветовых показателей с ходом среднесуточной температуры и количеством выпавших осадков в период рождения зверьков, принадлежащих к разным сезонным генерациям*, было установлено влияние на величину показателей белизны и оттенка этих экологических факторов. Оно заключалось в увеличении яркости окраски и содержания охристо-рыжих тонов при значительном понижении температуры и высокой влажности [7].
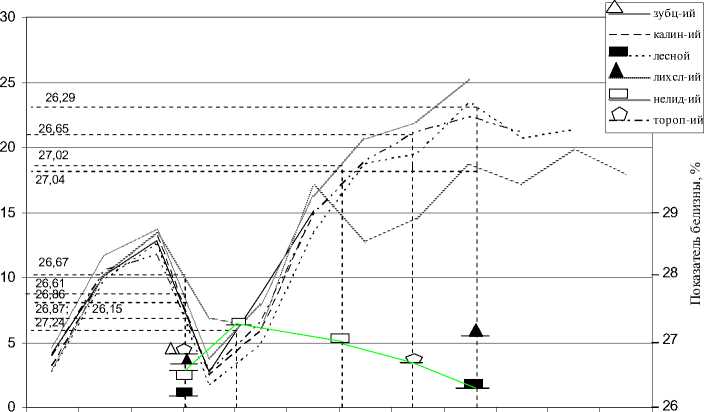
Особенно отчетливо выявились указанные закономерности при анализе зависимости цветовых характеристик меха исследованных серий полевок от хода среднесуточных температур в местах их отлова (рис. 1, 2). Наиболее тесная связь с данным фактором установлена для яркости окраски; в настоящем случае ход кривой, соединяющей значения показателя белизны последовательных пометов полевок, родившихся в 1999 г., является «зеркальным» отражением хода температурных кривых этого года (рис. 1). При этом показатель белизны ведет себя как типичная модификация, т. е. признак, находящийся в большой зависимости от условий окружающей среды.
Таблица 1
|
Районы |
Даты поимки зверьков |
Сезонная генерация |
Расчетный период появления генерации |
N |
Белизна (Б) |
Оттенок (От) |
P (Mann-Whitney test), р≤0,05 |
|||||||
|
M±m |
σ |
M±m |
σ |
К 1 /К 3 |
К 1 /К 2 |
К 2 /К 3 |
||||||||
|
Б |
От |
Б |
От |
Б |
От |
|||||||||
|
Зубцовский район |
18.06. – 21.06.99 |
К 1 |
01. – 05.05.99 |
27 |
26,860± 0,29 |
1,52 |
1,229± 0,01 |
0,07 |
– |
– |
– |
– |
– |
– |
|
Лесной район |
29.07. – 04.08.99 |
К 3 |
2 – 3 декада июня |
10 |
26,286± 0,46 |
1,45 |
1,179± 0,02 |
0,07 |
0,535 |
0,554 |
– |
– |
– |
– |
|
К 1 -К 2 |
1 – 2 декада мая |
13 |
26,146± 0,32 |
1,14 |
1,187± 0,01 |
0,03 |
||||||||
|
Лихославльский район |
31.07. – 04.08.00 |
К 3 |
2 – 3 декада июня |
12 |
27,044± 0,36 |
1,23 |
1,174± 0,01 |
0,04 |
0,843 |
0,053 |
– |
– |
– |
– |
|
К 1 -К 2 |
1 – 2 декада мая |
10 |
26,670± 0,26 |
0,83 |
1,204± 0,01 |
0,04 |
||||||||
|
Нелидовский район |
04.07. – 07.07.99 |
К 3 |
04.06. – 07.06.99 |
21 |
27,016± 0,18 |
0,85 |
1,180± 0,01 |
0,03 |
0,198 |
0,754 |
0,553 |
0,233 |
0,145 |
0,399 |
|
К 2 |
15.05. – 18.05.99 |
15 |
27,241± 0,37 |
1,43 |
1,189± 0,01 |
0,06 |
||||||||
|
К 1 |
04.05. – 07.05.99 |
19 |
26,613± 0,28 |
0,99 |
1,182± 0,01 |
0,03 |
||||||||
|
Торопецкий район |
16.07. – 21.07.99 |
К 3 |
01.06. – 06.06.99 |
26 |
26,649± 0,25 |
1,25 |
1,208± 0,01 |
0,04 |
0,557 |
0,059 |
– |
– |
– |
– |
|
К 1 |
2.05. – 07.05.99 |
37 |
26,867± 0,26 |
1,58 |
1,185± 0,01 |
0,04 |
||||||||
Особенности окраски меха разных сезонных генераций рыжей полевки из некоторых районов Тверской области
123123 декады 123123
Рис. 1. Влияние среднесуточной температуры на величину показателя белизны рыжей полевки Тверской области (подекадно с апреля месяца по июль включительно)
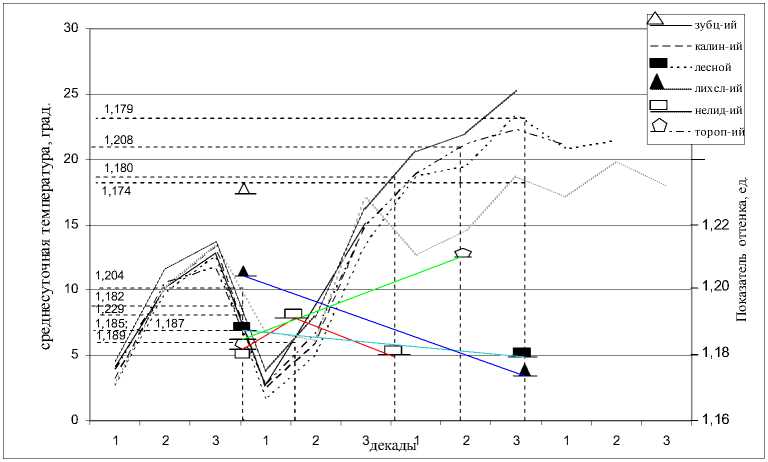
Рис. 2. Влияние среднесуточной температуры на величину показателя оттенка рыжей полевки Тверской области (подекадно с апреля месяца по июль включительно)
Степень выраженности модификации пропорциональна силе и продолжительности действия на организм вызывающего модификацию фактора [4]. Согласно этому у раннелетней генерации полевок из Нелидовского р-на наблюдается посветление окраски при значительном похолодании в первой декаде мая, а у сеголеток весенней и летней генераций в целом по Тверской области отмечается потемнение меха при резком потеплении в апреле месяце и продолжительном воздействии высоких летних температур.
Отметим, что данные закономерности наиболее наглядны при графическом анализе и менее очевидны при статистической обработке материала, поскольку различия сезонных генераций рыжей полевки по величинам цветовых показателей не достигают уровня достоверности (табл. 1).
Механизм влияния климатических факторов на животных заключается в сложном действии их на метаболизм, приводящем в конечном счете к комплексу приспособительных изменений, затрагивающих организм в целом. При химической терморегуляции, свойственной мелким млекопитающим, под влиянием пониженных температур наблюдается интенсификация окислительных процессов в организме. В эти процессы вовлекаются скелетные мышцы, а также усиливается обмен липидов – главного источника энергии. Основным местом образования жировой ткани, известной под названием подкожной жировой клетчатки, является дерма – второй, более глубокий слой кожи мезодермального происхождения. Первый слой кожи – эпидермис – участвует в образовании пигмента и имеет эктодермальное происхождение. Биосинтез меланина происходит в клетках меланоцитах, локализующихся в базальном слое эпидермиса, волосяных луковицах и ряде других структур. Волосяной фолликул имеет сложное строение: он состоит из кератинизированных эпителиальных клеток, а в основании имеет дермальный волосяной сосочек, содержащий соединительную ткань и кровеносные сосуды. Именно за счет мезодермального компонента происходит рост волос, причем для пигментации волос значим генотип только данного компонента волосяного фолликула, а генотипы эктодермального компонента роли не играют [19]. Таким образом, между упомянутыми слоями кожных покровов существует тесная генетическая, структурная и функциональная связь.
Можно предположить, что биохимические реакции в дерме также затрагивают надлежащий слой кожи, воздействуя, таким образом, на синтез пигментов. Оттенок окраски определяется интенсивностью и продолжительностью окислительного процесса ряда веществ – предшественников меланина. Главную роль в данном процессе биосинтеза пигментов играет фермент тирозиназа, активность которого и определяет некоторые особенности окраски. У животных генотипа агути в одной и той же волосяной луковице образуется и феомеланин и эумеланин, но в определенной последовательности. Появление феомеланиновой полосы совпадает с двумя обстоятельствами: 1) подавлением активности тирозиназы; 2) повышением скорости роста волос в связи с повышенной митотической активностью клеток [16]. Кроме этого существуют данные, свидетельствующие об усилении активности тирозиназы с повышением температуры среды, и связанной с этим гиперпигментацией [1]. По всей видимости, в последнем случае пигментация вызывается усилением синтеза эумеланина. Можно предположить, что тирозиназа является одним из основных механизмов переключения биосинтеза двух форм меланина, которые, таким образом, являются антагонистами относительно активности данного фермента. Активность тирозиназы, в свою очередь, находится под влиянием условий внешней среды.
Указанная температурная обусловленность показателя белизны может объясняться тесной связью ее величины с интенсивностью биохимических процессов в организме животных. Связь же показателя оттенка с климатическими факторами не столь явная, однако в пределах исследованных серий рыжих полевок были зарегистрированы вышеупомянутые тенденции в изменении этой цветовой характеристики как реакция на совокупность рассматриваемых экологических факторов. Интересно, что при графическом анализе влияния хода среднесуточных температур на окраску меха весенней и летней групп сеголеток отмечается разнонаправленный характер изменчивости показателей белизны и оттенка в пределах каждой точки отлова. В частности, при изучении реакции цветовых показателей на воздействие высоких летних температур во всех исследованных нами популяциях, кроме серии из Торопецкого района, обнаружено увеличение средних значений показателя белизны и уменьшение величин показателя оттенка. Так, температурным условиям в Лесном, Лихославльском и Нелидовском р-нах в период появления весенней группы сеголеток (конец апреля – начало мая – 12,6 – 13,70 С) соответствуют следующие значения показателя белизны: 26,146, 26,670 и 26,613%. В июне, при повышении среднесуточной температуры воздуха до 18,6 – 23,40С, яркость окраски летних пометов полевок в этих районах незначительно увеличивается: 26,286, 27,044 и 27,016% соответственно (р – 0,535, 0,843 и 0,198). Для весенней и летней групп сеголеток зафиксированы следующие изменения величин пока- зателя оттенка, также не достигающие уровня достоверности: в Лесном районе – с 1,187 до 1,179 ед. (р – 0,554), в Лихославльском – с 1,204 до 1,174 ед. (р – 0,053) и Нелидовском – с 1,182 (К1) до 1,180 ед. (группа К3) (р – 0,754). При сопоставлении окраски меха весенней и летней генераций полевок из Торопецкого района отмечается увеличение содержания в окраске охристо-рыжих тонов с 1,185 до 1,208 ед. (р – 0,059) и уменьшение яркости окраски с 26,867 до 26,649% (р – 0,557). То есть, несмотря на возможную специфическую реакцию полевок данной популяции, сохраняется разное направление изменчивости цветовых характеристик (рис. 1, 2).
По-видимому, оптические свойства меха, выражающиеся стандартными цветовыми показателями, соответствуют содержанию в остевом волосе двух форм меланина. В этом случае величина показателя оттенка может отражать ширину феомеланино-вой полосы и интенсивность цвета данного пигмента, а величина показателя белизны – насыщенность цвета остевых волос, зависящую от присутствия обеих форм меланина, но в большей мере – от относительного количества эумеланина. Ранее упоминалось, что темный и желтый пигмент являются антагонистами относительно активности фермента тирозиназы. Этим может объясняться разнонаправленная реакция цветовых характеристик на изменение температурных условий. Сам характер наблюдаемых реакций цветовых показателей также вполне согласуется с предполагаемым соответствием их величины относительному количеству определенного пигмента. Так, при повышении температуры активность тирозиназы увеличивается, что сопровождается усилением синтеза эумеланина и, как следствие, потемнением окраски. При снижении среднесуточной температуры происходит подавление активности данного фермента, что приводит к переключению с синтеза эумеланина на синтез феомеланина. В этом случае на волосе образуется более широкая рыжая полоса, количество эумеланина в зоне кольца (в случае его присутствия в этой зоне) уменьшается, а величина показателя оттенка свидетельствует об увеличении содержания в окраске меха рыжих тонов. Последнее положение хорошо иллюстрируется влиянием резкого майского похолодания на раннелетнюю генерацию полевок из Нелидовского района, а именно увеличивается содержание охристо-рыжих тонов до 1, 189 ед. на фоне общего посветления окраски до 27,241 % (рис. 1, 2).
Ширина феомеланиновой зоны и интенсивность цвета данного пигмента определяются также генетически (существует темный и светлый феомеланин) [16]. С этим вполне согласуются результаты эксперимента, полученные при попытке сближения окраски двух подвидовых форм узкочерепной полевки ( Microtus gregalis gregalis и M. g. major ), которые обладают резкими различиями по данному признаку. Анализ колориметрических характеристик особей, отклонившихся за пределы варьирования окраски исходной лабораторной популяции M. g. major , показал, что ликвидация разрыва между подвидами происходила практически только за счет показателя белизны [18]. На основании наших и литературных данных, свидетельствующих о различной степени зависимости от факторов внешней среды признаков окраски рыжей полевки, можно заключить, что показатель белизны характеризуется в своем проявлении значительной долей модификационной изменчивости, в то время как показатель оттенка, хоть и испытывает влияние климатических факторов, в основном формируется под влиянием генотипа. Это подтверждается проведенным ранее сопоставлением окраски меха разных возрастных групп в пределах изученных популяций: практически во всех популяциях сеголетки и взрослые полевки достоверно различаются величиной показателя белизны, одновременно представляя некое единство по величине показателя оттенка [9].
В заключение отметим, что, несмотря на многообразие физико-химических и гуморальных факторов, оказывающих влияние на процесс меланообразования в клетках, наличие известной зависимости уровня активности фермента тирозиназы от температуры среды позволяет считать именно этот механизм ведущим при изменении окраски мелких млекопитающих под воздействием факторов внешней среды. Обнаруженные закономерности действия климатических факторов – температуры и влажности – на признаки окраски меха европейской рыжей полевки, выражающиеся показателями белизны и оттенка, объясняются изменением интенсивности биохимических процессов в организме животных и активности тирозиназы, влияющей на особенности рас- пределения эу- и феомеланина в остевом волосе. При этом изменения яркости окраски под воздействием данных факторов имеют явно направленный характер, т. е. являются модификационными. Показатель оттенка менее зависим от факторов внешней среды и лучше отражает генотипические особенности популяций.
ON SOME ASPECTS OF PHYSIOLOGICO-BIOCHEMICAL PROCESSES DETERMINING THE VARIABILITY OF THE FUR COLORATION
Tver State University
The increase of red tint along with whitening of the general fur coloring under decreasing of average daily temperature has been shown on a series of Bank Voles, captured in Tver region. The phenomenon appears to be connected with the temperature-dependent activity of tyrosinase as well as with distribution of eu- and phaeomelanin in the hair of the vole. The ambiguity in the appearance of fur coloring characters dependant on external and internal causes is demonstrated.
