О некоторых закономерностях в распространении ящерицы Линдгольма (Sauria, Lacertidae) на юго-восточном побережье Крыма
 на юго-восточном побережье Крыма")
Автор: Кукушкин О.В.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 1 т.18, 2009 года.
Бесплатный доступ
Приводятся оригинальные данные о распространении эндемичной ящерицы Линдгольма (Darevskia lindholmi) на юго-восточном побережье Крыма. Современное распространение этого петрофильного и гигрофильного горного вида определяется, главным образом, наличием обширных крутосклонных обнажений горных пород и увлажнением. В условиях засушливого климата крайней восточной части Горного Крыма ареал D. lindholmi распадается на несколько некрупных изолятов. Численность локальных популяций варьирует от первых десятков до приблизительно одной тысячи особей. Фрагментация ареала относится, по всей видимости, к «ксеротермической» эпохе среднего голоцена, характеризовавшейся сокращением площади лесных массивов.
Ящерица линдгольма, распространение, аридный климат, крым, юго-восточное побережье
Короткий адрес: https://sciup.org/148314767
IDR: 148314767
Текст краткого сообщения О некоторых закономерностях в распространении ящерицы Линдгольма (Sauria, Lacertidae) на юго-восточном побережье Крыма
Ящерица Линдгольма, Darevskia lindholmi (Lantz et Cyrén, 1936) – эндемичный крымский представитель обширной группы скальных ящериц, распространенной, главным образом, на Кавказском перешейке (Ryabinin et al., 1996; MacCulloh et al., 2000) – петрофильный, гигрофильный и холодостойкий горно-лесной вид, предпочитающий скальные обнажения и обрывы. Вид весьма эвритопен (населяет разнообразные биотопы от лишенных растительности галечных пляжей до горных лесов и луговых степей яйл), обладает высокой терморезистентностью (обитает в районах с наибольшими и наименьшими в Крыму температурами) и является исключительно «заметным» элементом герпетокомплексов горной части полуострова. На большей части ареала D. lindholmi – фоновый вид со стабильной численностью, и его биогеоценотическая роль в экосистемах Горного Крыма весьма высока.
Интересный вопрос представляет распространение D. lindholmi на юговосточном побережье Крыма (южный макросклон Главного хребта от Алушты до Феодосии), характеризующемся, в сравнении с Южным берегом и западной частью Горного Крыма, более континентальным климатом. Вследствие преобладания западного направления влагонесущих атмосферных потоков пониженная юго-восточная часть Горного Крыма оказывается в тени яйлинской гряды, что определяет низкие годовые величины атмосферных осадков: обычно 310–360 мм в приморской полосе (примерно в 1,5–2 раза меньше, чем в центральной части ЮБК). В крайней восточной части района осевая часть Главного хребта теряет определенность (исчезает четкое деление на северный и южный макросклон), и возникает сочленение гор с равнинностепными и прибрежными ландшафтами, что, в свою очередь, определяет значительное разнообразие микроклиматов и растительности. В силу перечисленных причин именно на юго-восточном побережье общие тенденции биотопического распределения D. lindholmi проступают особенно четко. Только здесь этот обычнейший вид ящериц на обширных территориях может быть весьма редким. Основной блок данных о распространении D. lindholmi в восточном Крыму получен в 1997–2007 гг. (рис. 1)1. Приводящиеся в тексте геоморфологические и климатические характеристики района почерпнуты из литературных источников (Добрынин, 1938; Природа Карадага, 1989; Курорт Коктебель…, 1997; Ландшафтногеофизические условия…, 2001).
От самой западного пункта находок D. lindholmi на перешейке мыса Херсонес (44o34'N, 33o24'E) (Кукушкин, Свириденко, 2002) до Алушты – распространение вида практически сплошное. Далее на восток побережье расширяется, на поверхность выходят непрочные триасовые глинистые сланцы и аргиллиты, что определяет развитие характерного эрозионного лощинно-балочного рельефа. Южная граница ареала ящерицы Линдгольма отступает от берега моря вместе с Главным хребтом, сложенным верхнеюрскими известняками и конгломератами, и вплоть до окрестностей с. Морское этот вид в нижней части склона не встречается ввиду отсутствия характерных биотопов. Восточнее с. Морское, где к морю выходит ответвление Главной гряды, ареал D. lindholmi вновь вплотную приближается к побережью, и в рифовых горах между пос. Новый Свет и Судаком она почти столь же обычна, как в Западном Южнобережье. К востоку от Судака распространение D. lindholmi на побережье спорадично. Небольшая изолированная популяция населяет южное побережье небольшого (площадью около 55 га) рифового мыса Алчак (143 м н.у.м.), замыкающего Судакскую бухту с востока, окруженного холмистой степью и отделенного от близлежащих мест обитания вида полосой песчано-галечного пляжа. Вероятнее всего, ящерицы заселили этот мысок с севера по долине р. Суук-Су (Алепхор), впадающей в море близ его западных склонов. Расселение D. lindholmi вдоль лесистых речных долин представляет собой достаточно типичное явление. Например, проникновение этого вида в степное предгорье к северу от Белогорска, где он населяет сложенные палеогеновыми известняками останцы-куэсты, явно осуществлялось по долинам р. Биюк- и Кучук-Карасу. Изолированные популяции D. lindholmi известны также из долины р. Альма в западном предгорье (Щербак, 1966). Восточнее Алчака граница ареала вновь отступает от моря, и до известнякового хребта Эчкидаг ящерицы Линдгольма на побережье нет нигде, в том числе – на огромном скальном массиве полуострова Меганом (358 м н.у.м.), сложенном конгломератами и песчаниками. По-видимому, отсутствие здесь D. lindholmi обусловлено исключительной засушливостью климата этого выступающего в море низкогорного массива (годовая сумма осадков составляет здесь всего 272 мм, и климат имеет явные черты полупустынного), и (или) его изолированным по отношению к соседним возвышенностям положением (гористая часть полуострова отделена от ближайших горных кряжей овражистой степной перемычкой шириной 8,5–10 км). Далее к востоку, на побережье обширных бухт Чалки и Лисья D. lindholmi также не встречается – берег здесь сложен глинами и песчаниками, и скалистые вершины хр. Эчкидаг отделены от моря остепненными плато четвертичных континентальных террас и исполинскими бедлендами.
'45 01'Я 35т2#Ь
<14
<29,
<46
»39< <
^Феодосия ^ (Theodosia)
J5 СтарыйКрым x. (Staryi Krym)
яйла Карабй h (plateau Rarabjl 33 >32 *
Белогорск *w (Belogorsk) /^
21>^|
Судак 20 (Sudak)
ЧЕРНОЕ MOPE (BLACK SEA)
Карадагский заповедник (Karadagh Reserve)
/ 44
^Алушта
< (Alushta) 44°41'N, 34°24'E 745
45 06^N, 34 39(E X
» ?55 5^v^4
s#3 Полуостров Меганом 42^ 'I*' (Meganom Peninsula)
Условные обозначения: < пункты находок D. lindholmi; О вид не выявлен (отсутствует); 7 вид не найден, но его обитание вероятно;
х/ вероятная граница ареала;
(серым цветом выделены крупнейшие лесные массивы).
Рис. 1. Распространение Darevskia lindholmi в восточной части Горного Крыма, по данным автора2 (подчеркиванием выделены пункты обитания популяций, изолированных от основного ареала): 1 – черта г. Феодосия, генуэзская крепость ; 2 – окр. пгт. Орджоникидзе, мыс Киик-Атлама (Щербак, 1966) (?); 3 – Карадагский заповедник, Береговой хребет; 4 – Карадагский заповедник, хр. Легенер, Балвалы-Кая, Сюрю-Кая, г. Малый Карадаг ; 5 – окр. пгт. Щебетовка, г. Папас-Тепе; 6 – окр. пгт. Щебетовка, долина р. Биюк-Узень; 7 – окр. пгт. Щебетовка, хр. Курбан-Кая; 8 – окр. пгт. Щебетовка, балка Буран-Таш; 9 – окр. пгт. Курортное, хр.
2Изучение каталогов коллекций D. lindholmi в фондах Зоологического музея ННПМ НАНУ (Киев) и Музея природы Харьковского национального университета не дало нам новых точек находок.
Эчкидаг; 10 – окр. пгт. Солнечная Долина, хр. Чалки; 11 – окр. пгт. Солнечная долина, г. Парсук-Кая, пер. Синор; 12 – окр. пгт. Краснокаменка, г. Сандык-Кая, Сочарчикон-Кая; 13 – окр. пгт Краснокаменка, отроги хр. Туар-Алан и Френк-Мезер; 14 – окр. г. Старый Крым, балка Караин-Дере; 15 – окр. г. Старый Крым, г. Большой Агармыш (?); 16 – окр. с. Дачное, хр. Таракташ (?); 17 – окр. г. Судак, мыс Алчак ; 18 – черта г. Судак, мыс Кыз-Кулле; 19 – окр. пгт. Новый Свет, г. Сокол; 20 – окр. пгт. Новый Свет, мыс Капчик; 21 – окр. пгт. Новый Свет, г. Караул-Оба; 22 – окр. г. Судак, г. Перчем; 23 – окр. с. Веселое, г. Вигла; 24 – окр. с. Морское, г. Папая-Кая и мыс Ай-Фока (?); 25 – окр. с. Громовка, долина р. Шелен; 26 – окр. с. Ворон, г. Плакья; 27 – окр. с. Зеленогорье, каньон р. Арпат; 28 – окр. с. Зеленогорье, г. Муэдзин-Кая и Чок-Сары-Кая; 29 – окр. с. Зеленогорье, отроги г. Построфиль; 30 – окр. с. Приветное, пер. Биюк-Капу; 31 – склоны яйлы Караби, урочище Чигинтра; 32 – плато Караби; 33 – окр. с. Генеральское, склоны Караби, долина р. Су-Ат; 34 – окр. с. Генеральское, ущелье Хапхал; 35 – черта с. Генеральское, долина р. Улу-Узень; 36 – яйла Тырке; 37 – яйла Демерджи; 38 – водопад Джурла; 39 – окр. с. Лучистое, г. Южная Демерджи; 40 – окр. с. Семидворье, каньон р. Едыфлер (Тапшан-Гя); 41 – окр. пос. Сотера, каньон р. Алака; 42 – окр. пос. Сотера; 43 – окр. пос. Семидворье; 44 – окр. пансионата «Эврика»; 45 – г. Алушта; 46 – окр. с. Пчелиное, отроги Караби-Яйлы; 47 – окр. с. Красноселовка, пер. Кокасан; 48 – окр. с. Головановка, г. Чомбай; 49 – окр. с. Карасевка; 50 – окр. с. Синекаменка, хр. Кок-Таш; 51 – окр. с. Курское, г. Бор-Кая; 52 – окр. с. Мичуринское, г. Бурундук-Кая (?); 53 – окр. с. Вишенное, г. Ак-Кая ; 54 – окр. с. Белая Скала, г. Аджилар ; 55 – окр. с. Мироновка, г. Биюк-Сарак-Кая (?)
Наиболее ярко закономерности в распространении вида проявляются в Карадагском заповеднике с его сложным геологическим строением, исключительно пересеченным рельефом и малым количеством осадков (в нижнем приморском поясе их сумма варьирует от года к году 199 до 683 мм, в среднем 357). Здесь же его распространение изучено наиболее детально (рис. 2). Ареал D. lindholmi в заповеднике распадается на несколько очагов площадью от 0,5–1 до нескольких сотен га, причем численность некоторых локальных группировок ограничивается первыми десятками особей (ситуация в своем роде уникальная!). Необходимо отметить, что Н.Н. Щербак (1984, 1989) оценивал общую численность «скальной» ящерицы на приморском склоне Берегового хребта в 20–30 особей, на всей территории заповедника – в 100 особей. Однако, по нашим данным, ее численность здесь многократно выше: по крайней мере, около одной тысячи особей. У моря при максимальном уровне активности на 100 м маршрута учитывали 3–12 (чаще < 5) особей, на склонах хр. Береговой, Малом Карадаге и в рифовой части заповедника – 1–4 особи (при ширине учетной полосы 3 м). На западе в сходных биотопах плотность популяций в норме значительно выше – 1 экз./ 5–10 м маршрута. Наиболее крупная популяция населяет вулканогенный (сложенный базальтами, спилитами, кератофирами и туфами) Береговой хребет, причем ящерицы тяготеют к глыбовым нагромождениям побережья, временным водотокам и самому нижнему ярусу приморских клифов, где сочатся многочисленные конденсационные родники, и в наибольшей степени сказывается увлажняющее влияние морских (адвективных) туманов. Небольшая группировка выявлена на крупном (около 35 м н.у.м.)
абразионном останце-островке Шайтан-Капу – на удалении 85 м от берега. К вершинам Берегового хребта численность вида значительно снижается, и близ его гребня обитают лишь малочисленные локальные группировки. Отсутствует в роще древовидного можжевельника ( Juniperus excelsa ) на южных обрывах западной части хр. Кара-Агач (в наиболее теплообеспеченном районе заповедника), но появляется в глубоких ущельях, прорезающих центральную часть этого хребта выше мыска Харсыз-Иван, где береговая линия поворачивает на северо-восток. В западной (нагорной) части заповедника ящерица Линдгольма обычна только на высотах 300–450 м н.у.м. – в скалах субмеридиональных известняковых хребтов Сюрю-Кая, Балалы-Кая, Легенер, Икылмак-Кая, получающих наибольшее количество осадков (плювиометрический градиент составляет 60 мм/ 100 м подъема) либо близ расположенных на меньшей высоте карстовых источников. Симптоматично, что D. lindholmi не найдена в скалах привершинья (350–450 м н.у.м.) наиболее высокой горы – Святой, но встречается в очень небольшом числе (не более 20 особей!) на меньших высотах (270–300 м н.у.м.) на одном из уступов г. Малый Карадаг близ самого мощного в вулканогенной части заповедника источника Гяур-Чешме, на склоне юго-западной экспозиции.
Мы разделяем точку зрения Н.Н. Щербака (1984, c. 24), что на Карадаге сохранились «явно угнетенные и реликтовые популяции, оставшиеся от единого горно-крымского ареала после уничтожения лесов». Однако, на наш взгляд, антропогенный фактор не играл решающей роли в формировании современного ареала D. lindholmi , и такое положение вещей объясняется значительным остепнением Карадагской горной группы в голоцене, сопровождавшимся исчезновением или сокращением численности многих мезофильных фаунистических элементов (Смирнов и др., 1959).
К востоку от Коктебеля, на невысоких (до 240 м н.у.м.) Янышарских хребтах распространены мелкогорные фриганно-степные ландшафты, малопригодные для жизни D. lindholmi (имеются лишь мелкие разрозненные скальные массивы). При поисках на мысе Киик-Атлама (в ландшафтах, весьма сходных с меганомскими) выявить этот вид также не удалось, однако он приводился для данного пункта ранее (Щербак, 1966), и нельзя исключать, что на обрывистой концевой возвышенности мыса (157 м н.у.м.), сложенной нижнемеловыми конгломератами и далеко выдвинутой в море, сохраняется малочисленная популяция (возможно, полусинантропная). На такую возможность указывает и недавняя находка ящерицы Линдгольма в генуэзской крепости Феодосии – в 7 км к северу-северо-востоку от Киик-Атламы. Поскольку на скалистом побережье близлежащего мыса Ильи, замыкающего Феодосийский залив с запада, вид не найден, можно предполагать его случайный завоз в Феодосию (Кафу), являвшуюся в позднем средневековье одним из крупнейших на Черном море портовых центров.
На основании вышеприведенных данных можно сделать вывод, что в современных климатических условиях распространение D. lindholmi лимитируется двумя главными факторами: 1) наличием обширных
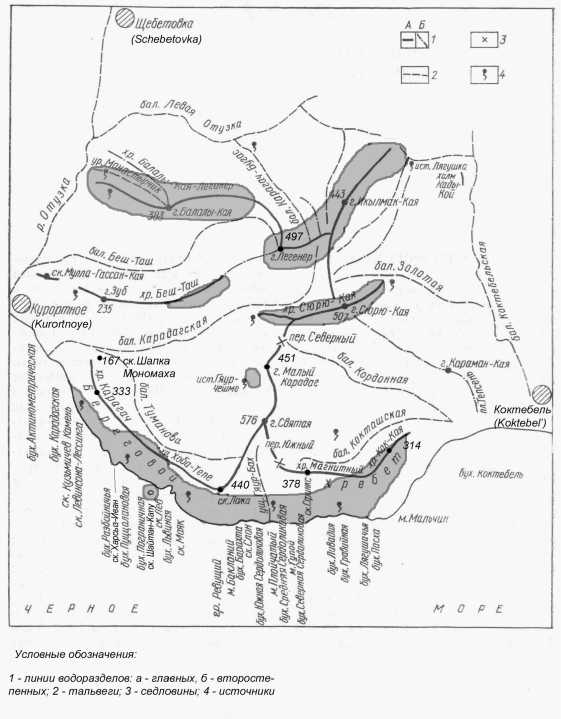
Рис. 2. Распространение Darevskia lindholmi в пределах Карадагской горной группы (районы обитания вида выделены серым цветом)
крутосклонных или отвесных обнажений трещиноватых горных пород со сложным микрорельефом поверхности (что обеспечивает наличие убежищ и сложную мозаику дневных температур, необходимую для реализации активности, направленной на поиск пищи); 2) влажностью местообитания (этот вероятно, влияет на характер распространения D. lindholmi опосредованно – через характер растительности и, в конечном итоге – кормность местообитания). Коэффициент увлажнения
Высоцкого-Иванова (отношение годовой суммы атмосферных осадков к годовой величине испаряемости) в местах обитания D. lindholmi равен или превышает 0,3 (для сопоставления, на Меганоме он составляет 0,26). О ведущей роли последнего фактора говорит тот факт, что в аридных местностях D. lindholmi проявляет себя как типичный петрофил, а в районах с условиями увлажнения, близкими к оптимальным (коэффициент увлажнения около 1), расширяет спектр занимаемых биотопов, и нередко обходится без крупных скальных выходов, населяя груды камней, щебнистые обрывы вдоль обочин дорог, шиферные склоны и стволы деревьев. Можно предполагать, что в аридных местностях на характер распространения этого вида влияет также литогенная основа местности. При прочих равных условиях ящерица Линдгольма чаще и в большем числе встречается на водорастворимых известняках, нежели на конгломератах. Данная особенность четко заметна в районах, где эти горные породы распространены равно широко (на востоке – в округе Судака, на западе – в восточных и юго-восточных окрестностях Балаклавы). Предположительно, определенные ограничения на распространение вида накладывает микроклимат убежищ, используемых для откладки яиц, в значительной мере зависящий от влагоемкости горной породы (таким образом, действие данного фактора осуществляется через успех воспроизводства популяции).
В заключение отметим, что индикаторами наличия D. lindholmi могут выступать относительно влаголюбивые виды флоры. Так, самая западная точка встреч D. lindholmi близ гребня хр. Кара-Агач совпадает с появлением рощ ясеня (Fraxinus excelsior, F. oxycarpa) и каркаса голого (Celtis glabrata) и крупных куртин резухи кавказской (Arabis caucasica) на скалах. Последняя отмечает также места обитания вида на г. Малый Карадаг. В основных очагах распространения D. lindholmi в западной части заповедника широко распространены редколесья можжевельника колючего (Juniperus oxycedrus). На мысе Алчак ящерицы найдены лишь на южных склонах, где произрастает самая восточная на побережье Крыма популяция асфоделины желтой (Asphodeline lutea).
По молекулярным данным, ящерица, близкая к анцестральной для D. saxicola (Eversmann, 1834) форме, проникла на территорию Крыма с Кавказа в позднем плиоцене или раннем плейстоцене (Ryabinin et al., 1996), когда между этими горными системами, по-видимому, существовала континентальная связь (Новосад, 1992; Шнюков и др., 1997), либо они находились на достаточно малом удалении друг от друга, чтобы обеспечить успех пассивной миграции ящериц на плавнике, выносимом кавказскими реками. Последнее предположение выглядит достаточно вероятным. Так, крупные обломки экзотических для Крыма горных пород, выявляемые в корневых системах выброшенных на берег деревьев и в отложениях пляжей, по сей день не представляют редкости на черноморском побережье Керченского полуострова (Голенко, 2006). Можно предполагать, что ареал D. lindholmi , приобрел близкие к современным очертания в раннем плейстоцене – в период, характеризовавшийся интенсивным поднятием Крымских гор, погружением их южной части под уровень моря и коренной перестройкой речной сети (Добрынин, 1937; Природа Карадага, 1989). В эпохи плейстоценовых похолоданий ареал ящерицы Линдгольма в зоне Главного хребта, где предполагается развитие слабого оледенения (Ена и др., 1991), вероятно, несколько сокращался, однако ничто не препятствовало ее расселению вдоль залесенных речных долин в предгорье. При аридизации климата в среднем голоцене, сопровождавшейся сокращением площади лесных массивов, ареал D. lindholmi на юго-восточном побережье и в северном предгорье был фрагментирован и приобрел реликтовые черты. О недавнем дроблении единого ареала ящерицы Линдгольма косвенно свидетельствует невысокий уровень межпопуляционных морфологических отличий. Оригинальная окраска тела ящериц в популяциях Сердоликовых бухт Карадага и г. Ак-Кая в восточном предгорье может быть обусловлена не только их нынешней изоляцией, но и своеобразием геохимических условий данных локалитетов. В пользу данного предположения свидетельствуют аналогичные тенденции в окраске ящериц, проявляющиеся в западнокрымских популяциях, обитающих на близких по микроэлементному составу породах (Шарыгин, 1999).
Список литературы О некоторых закономерностях в распространении ящерицы Линдгольма (Sauria, Lacertidae) на юго-восточном побережье Крыма
- Голенко В.К. Древний Киммерик и его округа. Симферополь: СОНАТ, 2006. 408 с.
- Добрынин Б.Ф. Характер берегов восточного Крыма от Меганома до Карадага (По данным экспедиции КИМП 1937 г.) // Уч. зап. МГУ. Вып. XIX. М., 1938. С. 7-24.
- Ена В.Г., Кузнецов В.Г., Лысенко Н.И. Охрана природопроявлений плейстоценовых ландшафтов Горного Крыма // Экологические аспекты охраны природы Крыма: Сб. науч. тр. Киев, 1991. С. 11-14.
- Кукушкин О.В., Свириденко Е.Ю. Находки меланистических особей скальной ящерицы (Darevskia lindholmi) в Крыму // Вестн. зоол. 2002. 36, № 3. С. 98.
- Курорт Коктебель. Природные ресурсы и урбоэкологический прогноз / Под ред. А.А. Вронского. Киев: Наукова думка, 1997. 134 с.
