О питании птенцов и гнездовых повадках поползня Sitta еurораеа

Автор: Бардин А.В.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 192 т.11, 2002 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149454
IDR: 140149454
On nestling diet and nesting habits of the wood nuthatch Sitta europaea
Текст статьи О питании птенцов и гнездовых повадках поползня Sitta еurораеа
Настоящие поползни широко распространены в лесах и горах Голарктической и Индо-Малайской областей. Наиболее разнообразны они в Южной Азии, где обитает 15 из 24 выделяемых в настоящее время видов рода Sitta (см.: Harrap, Quinn 1996). Некоторые виды поползней имеют очень ограниченное распространение. Таковы открытый в 1975 году кабильский поползень Sitta ledanti, известный из четырёх лесных массивов в горных хребтах Телль-Атласа в Алжире, эндемик Корсики S. whiteheadi, а также белобровый поползень S. victoriae, обитающий только на горе Виктория в горной системе Аракан-Йома в Бирме.
Самый обширный ареал среди поползней имеет евразийский 5. еигораеа. Формы, включаемые в настоящее время в состав этого политипического вида, распространены на северо-западе Африки (системы горных хребтов Эль-Риф и Средний Атлас в Марокко) и на огромных пространствах Евразии от Пиренейского полуострова, южной половины Великобритании и Скандинавии до Камчатки, Сахалина, Курил, Японских островов и восточного Китая, включая о-в Тайвань, на полуострове Малая Азия, Кавказе, в Эльбурсе, Загросе, небольшом участке восточного Тянь-Шаня.
В пределах своего громадного ареала обыкновенный поползень образует около двух десятков географических рас, входящих в большой комплекс форм (всего их описано более 60). Разные систематики рассматривают формы этого сложного комплекса в составе разного числа видов (и даже родов — Бутурлин 1916). В последней сводке (Harrap, Quinn 1996) самостоятельными видами признаются S. еигораеа Linnaeus, 1758, S. nagaensis Godwin-Austen, 1874, S. cashmirensis Brooks, 1871 и S. castanea Lesson, 1831. Ч.Вури (Vauri 1959) рассматривает их всех как один вид. М.А.Воинственский (1954) первые три объединяет в один вид, включая в него ещё и форму yunnanensis Grant, 1900, a castanea считает самостоятельным видом. По всей видимости, систематика этого комплекса будет ещё неоднократно пересматриваться.
Здесь важно обратить внимание на то обстоятельство, что хотя обыкновенный, или евразийский поползень и считается хорошо изученной птицей, это справедливо лишь для немногих европейских рас, прежде всего из группы “caesia”. Экология же большинства остальных форм на основной части ареала этого политипического вида остаётся совершено не исследованной. В том числе и в отношении питания и трофоценотических связей, Рус. орнитол. журн. 2001 Экспресс-выпуск №192 AQQ которые, как известно, определяют положение вида в экосистеме и многие особенности его биологии (Новиков 2001; Рахилин 2001). Что же касается птенцового питания, то в этом отношении все формы поползня изучены очень плохо, что связано с труднодоступностью гнёзд в естественных дуплах и редким гнездованием этих птиц в искусственных гнездовьях. На всём Северо-Западе России этот вопрос изучал только Э.В.Гавлюк (1972), наблюдавший за одним гнездом поползня в Ленинградской области.
В данном сообщении описаны наблюдения за выкармливанием птенцов в одном гнезде поползня* на Куршской косе (Калининградская обл.) в 1982 году. Работа является частью исследований по питанию птенцов разных видов птиц, проводившихся на Биологической станции Зоологического института в пос. Рыбачий. Методика сбора и обработки материала подробно описана ранее (Бардин и др. 1991).
На Куршской косе поползень малочислен. В широкой части косы у пос. Рыбачий, где сохранились участки широколиственного леса с липой, дубом, грабом, ясенем, в 1980-1990 обитали 2-3 пары этих птиц. Хотя с 1980 года в лесах российской части косы мы начали массовую развеску дуплянок, число которых вскоре превысило 1 тыс., в дощатом искусственном гнездовье поползни гнездились лишь один раз, в 1982. Они заняли дуплянку, где до этого два года подряд гнездились мухоловки-пеструшки Ficedula hypoleuca. Гнездовье располагалось на чёрной ольхе на высоте 3 м в черно-ольшанике, подтопленном благодаря деятельности бобров. Гнездовой участок поползней захватывал также зарастающий болотистый луг, два небольших заболоченных озерка, березняк, участок елового леса с примесью осины и берёзы. Примерно в 0.7 км начинался сосновый лес.
Весьма своеобразно при размножении в скворешнице проявились характерные повадки поползня, связанные с выбором и обустройством гнездового дупла. Как известно, эти птицы чаще всего занимают старые дятловые дупла, предпочитая те, леток которых уже “заплыл” и стал узким. При этом они почти всегда поселяются там, где в одном или нескольких рядом стоящих деревьях имеется не одно, а сразу несколько дупел. Поэтому гнездящиеся пары нередко оказываются в тесном соседстве с другими дуплогнездниками, обычно опережая их в сроках размножения. В наблюдавшемся случае поползни тоже поселились на участке, где в пределах видимости было вывешено пять дуплянок, а одна висела в 3 м от занятой ими, и в ней гнездились скворцы Sturnus vulgaris.
Из всех наших дуплогнездников, поползень предпочитает дупла с самым узким летком. В занятых им дуплах средний диаметр входного отверстия, по наблюдениям в разных регионах, составляет 2.8-3.1 см (Lohrl 1957, 1958; Нечаев 1991; Cramp, Perrins 1993), а в отдельных случаях может быть и ещё меньше — 2.3 (Султанов, Мухтаров 1995) и даже 2.2 см (Бардин 1983). Сами поползни в такие летки пролезают с видимым трудом. Поэтому для них важно, чтобы края входа были гладкими, чего они,— способные как долбить, так и “штукатурить”,— могут добиться сами.
Находившаяся под наблюдением пара заняла просторное дощатое гнездовье с узким летком (дно 12x12 см, расстояние от нижнего края летка до дна 20 см, диаметр летка 3.0 см). Края летка дуплянки были подолблены. По моим наблюдениям на Северо-Западе, такое ощипывание краёв летка типично для поползней и наблюдается даже в тех случаях, когда леток частично замазывается глиной. При необходимости поползни могут значительно расширить леток, но даже если этого не требуется, его края они всегда немного раздалбливают и ощипывают, как это делают многие синицы. Возможно, такая маркировка имеет демонстративное значение (сигнал, что дупло занято)*. Как и многие синицы, поползни способны не только расширять леток, но и расчищать полость дупла, выбрасывая остатки старых гнёзд, удаляя гнилую древесину. В рассматриваемом случае из дуплянки был тщательно удалён материал прошлогоднего гнезда мухоловки-пеструшки. Иногда поползни настолько значительно расширяют полость дупла в трухлявой древесине, что можно говорить даже о самостоятельном его изготовлении (Воинственский 1954; Lohrl 1957, 1958, 1988). Однако обыкновенные поползни никогда не начинают долбить дупло “с нуля”, они используют уже имеющиеся углубления на месте выгнившего сучка, начатого дятлового дупла и т.п. К тому же значительное расширение полости дупла у S. еигораеа — случай исключительно редкий (как, например, у Parus major). В то же время среди поползней, как и среди синиц, есть виды, для которых самостоятельное выдалбливание дупел является правилом, например, S. pusilia, S. pygmaea, S. krueperi, S. canadensis, S. whiteheadi.
Другая особенность гнездостроительного поведения поползней — использование глины, навоза, мякоти ягод, смолы и млечного сока деревьев в качестве замазки для сужения летка и заделки щелей. Виды, выдалбливающие дупла самостоятельно, “замазку” совсем не используют или делают это очень редко, тогда как S. пеитауег, гнездящийся на скалах, практически все стены гнездового укрытия лепит из глины, которой может натаскать несколько килограммов. У 5. еигораеа использование “замазки” проявляется в разной степени. В Псковской и Ленинградской областях S. е. еигораеа редко используют глину в существенном количестве, и обмазка носит скорее такой-же ритуальный характер, как и ощипывание краёв летка. Например, в начале апреле 1976 я наблюдал за парой поползней в окрестностях Печор. Она поселилась в старой дуплистой осине в сосновом лесу, выбрав старое дупло Dendrocopos major с узким, заплывшим входом. Ни в дупле, ни по краям летка обмазки никакой не было. Однако в первые два-три дня после начала выстилки лотка приходилось видеть, как самка брала из лужи на лесной дороге глину и мазала ею кору дерева в полуметре от входа в гнездовое дупло. Похоже, что птица смогла найти глину нужной кондиции только после дождя, а найдя, стала лепить её “просто так”, совершенно не используя для укрепления самого дупла. На подобные случаи “бессмысленной” обмазки у поползня орнитологи уже не раз обращали внимание (Lohrl 1957, 1958, 1988; Благосклонов 1991, Султанов, Мухтаров 1995).
В случае на Куршской косе подобная деятельность поползней проявилась, можно сказать, в гипертрофированном виде. Крышка дуплянки, где они загнездились, вращалась на большом гвозде, вбитом в одном углу, а в противоположном крышка была “прихвачена” маленьким гвоздиком. При осмотре крышку чуть поднимали и поворачивали. Естественно, между крышкой и стенками дуплянки оставалась щель. Она и была основательно замазана глинистой землёй, которую поползни брали с находившейся рядом бобровой плотины. Края летка и внутренняя часть дуплянки остались совершенно чистыми. При первом же осмотре гнезда во время насиживания слой глины был нарушен. Поползни на следующий же день обновили обмазку. Когда началась работа с птенцами, крышку дуплянки приходилось открывать очень часто. Обмазка каждый раз разрушалась, но поползни постоянно её обновляли, причём с каждым разом наращивали её толщину. В конце концов они стали не просто замазывать щель, а ежедневно возводили практически полный свод под крышкой гнездовья. Сначала птицы использовали для этого глинистую землю, которую собирали на бобровой плотине и бортах прочищаемых бобрами канав. Потом, когда подросла трава и начался выпас коров, стали применять в качестве шпатлёвки вещество коровьих “лепёшек”, перемешивая его с землёй. Эта деятельность отнимала у птиц много времени, тем не менее они продолжали её вплоть до того дня, когда птенцам исполнилось 16 сут и работа с ними была прекращена.
Удалось заметить, что в штукатурных работах принимал участие и самец, хотя обычно считают, что обустройкой дупла и строительством гнезда у S. еигораеа занимается одна самка. Однако в литературе мнения по этому вопросу расходятся, и он требует специального изучения. Среди других видов рода Sitta описано большое разнообразие в характере участия полов в строительстве (правда, детальных наблюдений мало). Так, у S. krueperi дупло обычно выдалбливает самка, хотя порой в этом участвует и самец (Поливанова 2000). У У. pusilia, наоборот, дупло долбит только самец. У S. white-headi дупло выдалбливают оба члена пары. У S. пеитауег гнездо лепит самец, самка лишь изредка приносит материал для выстилки лотка, а у S. tephroronota строительством занимаются оба члена пары (см.: Cramp, Perrins 7ПЭ Рус. орнитол. журн. 2001 Экспресс-выпуск № 192
1993; Harrap, Quinn 1996). У североамериканского 5. carolinensis, экологически очень похожего на 5. еигораеа, гнездо строит одна самка (Kilham 1968).
Гнездо наблюдавшейся пары представляло собой ворох тонких пластинок коры сосны Pinus sylvestris, заполнявший почти треть объёма гнездовья, в котором буквально утопала насиживающая самка. В основании гнезда находились более грубые куски сосновой коры. Что удивительно, поблизости от гнезда сосны не росли, и за строительным материалом нужно было летать по крайней мере за 700 м. Кора сосны — излюбленный и практически единственный гнездовой материал у наших поползней (Бардин 1983), хотя в других регионах они используют и другие похожие по свойствам материалы. Например, гнёзда S. е. sakhalinensis, найденные на Сахалине (Нечаев 1991, с. 563), состояли “из пластинок коры смородины, ивы и ильма, лоскутов бересты каменной или белой берёзы, кусочков листьев осок и злаков”. Гнёзда S. е. asiatica, обследованные В.В.Правосудовым (1991, 1993) на р. Яна в Магаданской обл., состояли из чешуек коры лиственицы и берёзы, а в основании встречались сухие листья.
Характерно, что в гнёздах поползней никогда не бывает такого обилия паразитов (блох, личинок мух), как в гнёздах многих других дуплогнёздни-ков — в т.ч. и у гнездившегося на соседнем дереве скворца.
Описываемое гнездо обнаружено 10 мая 1982 с насиженной кладкой из 8 яиц. Все 8 птенцов вылупились в течение первой половины дня 12 мая. Приведу описание новорождённых птенцов, согласующееся с имеющимися в литературе. Кожа телесно-розового цвета. Клюв серовато-желтоватый, яйцевой зуб белый. Ротовая полость и язык жёлтые, с возрастом их окраска становится насыщеннее. Клювные валики белые с желтоватым оттенком, с возрастом становятся чисто-белыми. Когти светло-серые. Тёмнопепельный пух расположен на 4 пуховых птерилиях: надглазничной (длина пушинок до 11 мм, 6-10 пучков), затылочной (13 мм, 4 пучка с каждой стороны), плечевой (14 мм, 6 пучков) и спинной (до 15 мм, 6 пучков с каждом из двух рядов).
Развиваются птенцы поползня довольно медленно. Лишь в возрасте 7 сут* над поверхностью кожи начали появляться пеньки перьев (в числе первых — трубки маховых). Приоткрывать глаза птенцы начали в возрасте 10 сут. Кисточки маховых появились в возрасте 12 сут. Поверхность крыла стала сплошной (исчезли щели между большими верхними кроющими и разворачивающимися опахалами маховых) в возрасте 18 сут. В это время птенцы уже способны при опасности выпархивать из гнезда, поэтому в 16 сут работа с ними была закончена. Все восемь птенцов оставили гнездо 6 июня в течение первой половины дня, т.е. провели в гнезде 25 полных суток и вылетели на 26-е. В сводках указано, что птенцы европейских форм S. еигораеа сидят в гнезде от 18 до 28, обычно 23-24 сут (Воинствен-ский 1954; Cramp, Perrins 1993; Harrap, Quinn 1996). В.А.Нечаев (1991) пишет, что птенцы S. е. sakhalinensis находятся в гнезде около 25 сут. У S. е. asiatica в Магаданской обл., по наблюдениям за 31 гнездом, период пребы- вания птенцов в гнезде был 18-22, в среднем 20 сут (Правосудов 1991). Не исключено, правда, что эти гнёзда птенцы покинули несколько раньше обычного срока, поскольку подвергались регулярному обследованию.
В целом у поползня, как и многих других воробьиных, можно говорить лишь об определённом периоде постэмбрионального развития (стадия слётка), когда происходит оставление гнезда. Характерный признак вступления молодых особей в этот возрастной период — появление в их репертуаре т.н. птенцового призывного крика. Конкретные же сроки вылета индивидуальны и варьируют в зависимости от внешних обстоятельств и комфортности условий в гнезде. При этом у разных видов стадия слётка может приходиться на разный абсолютный возраст и иметь разную продолжительность (см. также: Бардин 1985; Марковец 1991).
Самка в наблюдавшемся гнезде оказалась очень доверчивой. Когда она сидела на яйцах или птенцах, то позволяла брать себя рукой, а посаженная обратно, оставалась в гнезде. Её практически невозможно было выгнать из дуплянки, и яйца и птенцов приходилось брать из-под неё, чему птица всячески препятствавала и даже яростно клевала руку; но в то же время охотно брала из пальцев предлагаемый ей корм. Это обстоятельство крайне затрудняло работу с птенцами, поэтому наложение лигатур пришось начать лишь через 8 сут после вылупления, когда самка стала проводить в гнезде меньше времени. Удивительная доверчивость некоторых особей (не прирученных, а совершенно диких) уже была описана у целого ряда птиц (Мальчевский 1959; Прокофьева 2002). Мне приходилось встречать таких индивидуумов ещё у пухляка Pants montanus и коноплянки Cannabina саппаЫпа.
По сравнению с синицами, у поползня птенцы более активны и крикливы с первого дня жизни. Уже с возраста 7-8 сут писк голодных птенцов можно услышать за десяток шагов от гнездового дерева. Если птенцы не просят корм и не берут предлагаемую им пищу, самка реагирует на это обогреванием выводка. Временное снижение активности птенцов случалось и в результате наложения лигатур — самка в таких случая неизменно садилась греть птенцов. Если в это время прилетал с кормом самец, то он отдавал пищу самке, а она или съедала его, или пыталась отдать птенцам.
С 20 по 28 мая 1982 у птенцов наблюдавшегося выводка изъято 36 порций пищи, содержащих 131 пищевой объект общей массой 9.064 г (сухая масса 2.908 г). От 8-сут птенцов взята 1 порция, от 10-сут — 5, от 11-сут — 10, от 12-сут — 8 и от 16-сут птенцов — 12 порций пищи.
Рассмотрим состав пищи по крупным таксонам.
Odonata. 1 стрекоза четырёхпятнистая Libellula quadrimaculata. Скормлена с оторванной головой, но с крыльями. Сырая масса (w) 93 мг. Стрекозы составили 0.76% от общего числа объектов (% и), 3.2% от общей сырой массы (%ww), 3.6% от общей сухой массы (%Jw).
Hemiptera. 1 мелкий неопределённый клоп (0.76%я, 0.16%ww, 0.19Jw).
Coleoptera. 10 жуков: 4 щелкуна Elateridae, 1 мелкий жук из Chryso-melidae, 5 жуков из Dytiscidae — 4 бороздчатых полоскуна Asilus sulcatus и 1 желобчатый A. canaliculatus, 2 маленькие личинки Dryophthoridae. Крупные жуки были частью лишены головогруди, надкрылий и ног, т.е. птенцам 704 Рус. орнитол. журн. 2001 Экспресс-выпуск № 192
скармливались практически одни брюшки, в случае полоскунов набитые яйцами. Все Coleoptera: 9.16%я, 16.3%ww, 16A%dw.
Lepidoptera. 4 гусеницы: 2 Graphiphora augur, 1 Blepharita satura (Noctu-idae) и 1 мелкая гусеница Tortricidae. 1 куколка Noctuidae. 3 бабочки: совка, стеклянница Bembecia hylaeiformis и очень мелкая “моль”. Гусеницы, куколка и мелкая бабочка скормлены целиком, от крупных бабочек птенцам были даны только брюшки. Все Lepidoptera: 6.11%и, 13.55%ww, \\.%6%dw. Из них на стадии гусеницы: 3.05%и, 9.95%ww, 8.49%^w.
Diptera. Представлены только имаго. Крупные Tipula sp. (Tipulidae) — бб экз. (50.38%), 50.38% от сырой массы всего корма и 54.95% от сухой. Chironomus plumosus (Chironomidae) — 29 экз. (22.14%), 5.22%ww, 5.34%dw. Обнаружен также 1 экз. Syrphidae. Двукрылые составили основную часть в выборке птенцового корма: 79.39%я, 60.44%ww, 61.90%dw.
Aranei. 5 крупных крестовиков (Araneus angulatus и A. diadematus) — 3.82%и, 6.35%ww, SWodw.
Итак, основную долю (по сухой массе) в пище птенцов поползня занимали имаго двукрылых, прежде всего Tipulidae (62%), на втором месте были жуки (16.4%), на третьем — гусеницы (8%), на четвёртом — пауки (6%). С точки зрения, так сказать, гастрономической, 62% от массы всей пищи составили нежные имаго, 23% — грубые имаго (включая куколку) и 15% — пауки и гусеницы. Из-за небольшой величины выборки в пробах не обнаружены гастролиты и минеральный корм, в частности, раковины моллюсков, которых находили в корме птенцов поползня другие исследователи.
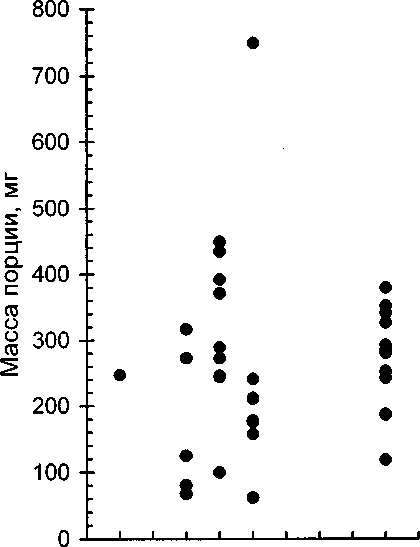
7 8 9 10 11 12 13 14 15 16 17
Возраст птенцов, сут
Рис. 1. Сырая масса порций корма, получаемых птенцами разного возраста.
• Средняя
- Максимальная
400 т --------------- : • Средняя
350 -- “ Максимальная _
300 "
го 250
Ё- - ф ■_
о:
го■
О 150
50 -;•
0 Л_____।_______।_______।_______।_______।_______।_______।_______।_______।।
7 8 9 1011 12 1314151617 Возраст птенцов, сут
Рис. 2. Сырая масса экземпляров корма птенцов разного возраста.
800 т
700 --
600 --
500 --
400 --
- Медиана
05 О О 05
100 --
0 -I—।—।—।—1—1—1—।—।—।—।—।—।
0 2 4 6 8 10 12
Число объектов в порции
Рис. 3. Сырая масса порции корма в зависимости от числа объектов в порции.
400 -г
350 -- - -----------------
-
• Средняя
300 -- - Масимальная га 250 -:
В- •"
200 --о:
05 о 150
-
50 ~ # "
0 -С-1—।—।—।—।—।—।—।—।—।—।—।
0 2 4 6 8 10 12
Число объектов в порции
Рис. 4. Сырая масса объекта корма в зависимости от числа объектов в порции.
В изученном возрастном интервале 8-16 сут (птенцы уже большие, скорость прироста массы тела уменьшилась, идёт формирование оперения) масса порций, число объектов в порции и масса кормовых объектов больше зависели от конкретной кормовой ситуации, чем от возраста (рис. 1,2). Поэтому количественные характеристики корма рассмотрим на объединённой выборке. За один раз поползни приносили птенцам от 1 до 12 пищевых объектов. В 14 порциях содержался 1 объект, в 1 порции — 2, в 2 порциях — 3, в 4 порциях — 4, в 7 порциях — 5, в 1 порции — 6, в 2 порциях — 8, в 1 порции — 10, в 2 порциях — 12 объектов. В среднем одна порция содержала 3.7 объекта (мода — 1, медиана — 3 объекта). Длина приносимых объектов достигала 40 мм (стрекоза). Из нескольких объектов в клюве взрослой птицы формировался комок, обычно имевший размеры 8x15 мм. Самый крупный комок (из комаров-долгоножек) имел размеры 10x27 мм. Сырая масса порции, приносимой взрослой птицей за один прилёт к гнезду, варьировала от 62 до 749 мг (2-3-й квартили: 87-327 мг). Средняя сырая масса порции составила 259, медиана — 247 мг. Средняя сухая масса порции — 83 мг. Таким образом, средняя масса порции корма, приносимая поползнями к гнезду за один прилёт, составляет порядка 1% их массы тела. Сырая масса одного объекта варьировала от 9 до 353 мг и в среднем была 69 мг. Средняя сухая масса объекта составила 22 мг.
Характерно, что масса порции почти не зависела от числа содержавшихся в ней объектов (рис. 3), т.е. поползни набирали некую оптимальную массу пищи, с которой и летели к гнезду. Чаще всего (40% случаев) поползни приносили лишь одно насекомое или паука за один раз. Наиболее крупные объекты встречались в порциях из одного или двух объектов. В последнем случае, судя по нескольким визуальным наблюдениям, первым добывался мелкий объект. В целом величина и масса объектов находились в обратной зависимости от их числа в порции (рис. 4). Нужно заметить, однако, что порции корма были неравнозначны. Одни можно назвать основными, доставляемыми в гнездо в определённом ритме, а другие — дополнительными, своего рода “добавкой”, дать которую родителей вынуждает реакция выпрашивания корма особо голодных птенцов, неутихнувшая после основного кормления. Эту добавочную пищу поползень за считанные секунды находит рядом с гнездом. Естественно, что такие дополнительные порции обычно меньше, и это несколько смазывает общую картину.
53 порции корма подросших птенцов (8-19 сут) на р. Луге в Ленинградской обл. в среднем содержали по 2.4 объекта (Гавлюк 1972). В 82 порциях, собранных В.А.Нечаевым (1991) на р. Айнской на Сахалине от птенцов 8-13-сут возраста, было в среднем по 1.7 объекта. В нашем случае порции корма птенцов в возрасте 8-16 сут в среднем состояли из 3.7 объекта. Во всех гнёздах, однако, поползни чаще всего приносили птенцам по одному экземпляру добычи, предпочитая охотиться на относительно крупных насекомых. В то же время в определённых условиях они способны собирать даже таких мелких насекомых, как тли (Нечаев 1991), которых на Северо-Западе России в пище поползней не находили (Прокофьева 2001).
Вслед за И.В.Прокофьевой (2001), нельзя не поражаться большому разнообразию птенцовой пищи поползней и способностью этих птиц быстро переключаться на массовые в данном месте и в данное время корма. Благодаря этому основным компонентом в выборках птенцового корма у разных выводков могут оказаться совершенно разные, экологически непохожие друг на друга беспозвоночные. Так, в Ленинградской обл. в одном гнезде в пище птенцов преобладали подёнки Syphlonurus linnaeanus и гусеницы совок (Гавлюк 1972), в другом родители чаще всего приносили птенцам имаго Trichoptera и Plecoptera (Прокофьева 2001). На Сахалине в пище одного выводка преобладали имаго Lepidoptera и Coleoptera, другого — гусеницы зимней пяденицы Operophtera brumata (Нечаев 1991). В нашем случае в Калининградской обл. в целом преобладали комары-долгоножки Tipula. Однако на этот результат, несомненно, сильное влияние оказал и чисто случайный фактор при выборе времени сбора образцов корма. Поползни ежедневно меняли свои предпочтения, поэтому в том же гнезде — но в другое время — основным компонентом в выборке могли бы оказаться и звонцы, и стрекозы, и жуки, и гусеницы.
Переходя к вопросу о способах добывания пищи поползнями, прежде всего нужно отметить чрезвычайно высокую пластичность их кормового поведения и способность быстро осваивать новые корма. На это обращали внимание все исследователи, проводившие наблюдения за этими птицами (Осмоловская, Формозов 1950; Образцов, Королькова 1954; Кошкина 1956; Lohrl 1957, 1958; Гавлюк 1972; Прокофьева 2001). В период выкармливания птенцов способы охоты поползня совершенно не соответствуют представлению о нём как высокоспециализированной птице-древолазе, предпочитающей ксилофагов (Иноземцев 1965). Он ловит беспозвоночных не только на стволах и толстых ветвях, но схватывает их с тонких веточек и листьев, кормится на кустарниках и травянистых растениях, много времени проводит на земле.
В период выкармливания птенцов поползень часто ловит насекомых в воздухе, взлетая с ветви или ствола наподобие мухоловки. Воздушную охоту поползней я видел и в Псковской, и в Калининградской, и в Белгородской областях. Приходилось также неоднократно наблюдать, как поползни во время лёта крупных насекомых совершали и более продолжительные охотничьи полёты. Ловкость, с какой эта, казалось бы, неприспособленная к воздушной охоте птица догоняет и схватывает в полёте крупную стрекозу или бабочку, всегда изумляет. Крупных жуков (например, навозников, жуков-носорогов, майских хрущей) в воздухе схватить обычно не удаётся: сбитый жук складывает крылья и падает на землю. Поползень приземляется вслед за ним и берёт (если находит) уже на земле, а затем летит на ближайшее дерево для разделки.
Иногда, поймав крупное насекомое, поползень засовывает его в щель коры и продолжает охоту. Вскоре он возвращается и берёт припрятанную добычу, соединяя её в одну порцию с только что пойманной. Когда птенцы сыты и не берут корм, поползень тоже может припрятать принесённый корм, чтобы скормить его птенцам чуть позже. Учитывая сильно выраженный у поползня инстинкт запасания корма, такое поведение не удивительно. Однако, если не считать этих отрывочных наблюдений, запасательное поведение поползня в гнездовой сезон совершенно не изучено.
Звонцов и комаров-долгоножек находившиеся под наблюдением поползни собирали на стволах деревьев; долгоножек они ловили и в воздухе. Водных жуков полоскунов им, вероятно, удалось найти в то время, когда они выползали из воды для откладки яиц (в их брюшках находились яйца). Одну бабочку совку поползень достал из ловчей сети паука-крестовика.
* Однако Э.В.Гавлюк (1972, с. 20), описывая воздушную охоту поползня, пишет следующее: “Кроме обычного поиска корма на стволах и толстых ветках, поползень иногда подстерегает добычу, сидя на каком-либо сучке. Заметив летящее насекомое, стремительно летит за ним, поймав, отлетает на некоторое расстояние и, сидя на ветке, зажимает добычу лапами и обрабатывает её”.
Кстати сказать, такой своеобразный способ добывания летающих насекомых, ещё не отмеченный в литературе, я часто наблюдал у больших синиц Parus major, за которыми вёл длительные наблюдения. В лесах Куршской косы, где благодаря регулярным массовым вылетам хирономид поддерживается поразительно высокая плотность пауков, в частности крестовиков, проверка ловчих сетей наряду с добыванием их хозяев — достаточно эффективный способ добычи корма для особей, научившихся этому приёму.
Корм для птенцов поползни собирают с такой же жадностью, с какой они осенью занимаются запасанием корма. Они менее разборчивы в отношении качества пищи для птенцов, чем, например, синицы, и хватают всё, что кажется съедобным. Поэтому они часто кормят птенцов более грубой пищей, чем едят сами. Находясь всегда в спешке, они если и разделывают добычу перед тем, как скормить её птенцам, то лишь для того, чтобы она могла пролезть им в глотку. При этом они быстро находят новые источники корма и легко переключаются на новые виды пищи, в том числе антропогенного происхождения (Lohrl 1957, 1958, 1988; Прокофьева 1998, 2001, 2002). Весьма скудные данные, имеющиеся в литературе, свидетельствуют о том, что поползни во время выкармливания птенцов используют настолько широкий набор кормов, что для этого сезона невозможно говорить о какой-либо трофической специализации, свойственной этому виду. В то же время мы сталкиваемся с ярко выраженной индивидуальной специалиацией, когда каждая гнездящаяся пара использует весьма ограниченный набор кормов, часто совершенно непохожий на рацион других пар.
