О полиморфизме некоторых гетеростильных растений лесостепного Зауралья

Автор: Демьянова Е.И.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2014 года.
Бесплатный доступ
Изучена структура популяций (соотношение морфологических форм) некоторых гетеростиль-ных растений: дистильных Limonium caspium, L. gmelinii, Goniolimon elatum и тристильных Lythrum virgatum и L. salicaria. Исследованы их антэкологические особенности: суточный ритм цветения, экология цветка, семенная продуктивность при свободном опылении и изоляции.
Гетеростилия, структура популяций, антэкология
Короткий адрес: https://sciup.org/147204669
IDR: 147204669 | УДК: 581.162
To the polymorphism of some heterostyled plants of the forest-steppe of the Trans-Urals
We studied the structure of some heterostyled plants populations from the forest-steppe Trans-Urals area (Limonium caspium, L. gmelinii, Goniolimon elatum, Lythrum virgatum, L. salicaria). All morphological types of the plants were represented at same ratio. The blooming began early in morning rapidly and at the same time. That provided cross pollination and high seed production.
Текст научной статьи О полиморфизме некоторых гетеростильных растений лесостепного Зауралья
физиологическая несовместимость появилась раньше разностолбчатости. Она исключала оплодотворение собственной пыльцой, но при этом непроизводительно погибало большое количество пыльцы и семяпочек. Появление гетеростилии ограничивает возможность самоопыления и тем уменьшает напрасное расходование гамет.
Цветки гетеростильных растений, кроме указанных выше различий, могут быть неодинаковы по величине и структуре пыльцевых зерен, их окраске и биохимизму, сосочковой поверхности рылец, количеству цветков в соцветии, размерам семяпочек и другим признакам. Степень выраженности указанных отличий неодинакова у разных видов. Так, у некоторых гетеростильных видов пыльцевой диморфизм развит слабо, у других полиморфизм ограничен высотой столбика, а пыльники имеют одинаковое положение во всех формах цветка.
Следует отметить, что взаимосвязь гетеростилии и самонесовместимости не всегда бывает абсолютна. Например, у тристильного Decodon verticillatus (сем. Lythraceae) при искусственном опылении во всевозможных комбинациях во всех случаях отмечен высокий уровень завязывания семян, что указывает на отсутствие несовместимости или ее рудиментарный характер [Ornduff, 1993].
По мнению Ч. Дарвина (1877), триморфные виды имеют определенное преимущество перед диморфными, поскольку у первых при совместном произрастании даже двух, а не трех форм больше возможностей быть принадлежащими к разным флоральным формам, и они смогут опылить друг друга. Напротив, если две особи диморфного вида относятся к одной и той же форме, они не дадут семян.
Структура популяций у большинства гетеро-стильных видов, как правило, константна: для нее характерна одинаковая представленность флораль-ных форм. Случаи их неравного соотношения связаны чаще всего с адвентивными популяциями (случайным заносом какой-либо одной формы или ее гибелью и другим причинам). Кроме того, в случае интенсивного вегетативного размножения создается искаженное соотношение морф в популяции, сохраняющееся в течение длительного периода (например, у водных гетеростилъных растений). На границах географического распространения вида отмечаются гомостильные особи, у которых наблюдается нормальное завязывание семян. Они самофертиль-ны, в отличие от обычных гетеростилъных растений. Таким образом, популяции гетеростилъных растений по структуре бывают весьма разнородны. Они могут быть представлены только гетеростиль-ными (ди- и тристильными) особями или гомо-стильными, или состоят из гетеро- и гомостильных растений. Семенная продуктивность гетеростильных растений будет зависеть от структуры популяций и ее размеров [Waites, Agren, 2004].
Гетеростилия - явление сравнительно редкое [Vuilleumier, 1967; Ganders, 1979; и др.]. По современным данным она обнаружена в 36 семействах и примерно в 175 родах [Агаджанян, 2000]. Гетеростилия встречается крайне неравномерно в разных филах покрытосеменных, не имеющих между собой филогенетического родства. Тем не менее, четко прослеживается тенденция большей приуроченности гетеростилии к двудольным, нежели однодольным растениям. По мнению исследователей, это связано с более широко представленным у двудольных биотического опыления, основными агентами которого являются насекомые [Baker, 1963, 1966; Vuilleumier, 1967; и др.]. Еще Ч. Дарвин в 1877 [Дарвин, 1948] подчеркивал, что одной из важных особенностей гетеростилъных растений является их энтомофильное опыление. Наиболее часто гетеростилия отмечена у Magnoliopsida в подклассах Dilleniidae, Rosidae, Lamiidae. В последнем подклассе по числу гетеростилъных родов выделяется сем. Rubiaceae (91 из 175 гетеростилъных родов). Феномен приуроченности гетеростилии к этому семейству, известный еще Ч. Дарвину, не расшифрован до сих пор. По степени встречаемости гетеростилии на родовом уровне двудольные более чем в пять раз превосходят однодольные [Агаджанян, 2000]. Согласно К. Фегри и Л. ван дер Пэйлу [1982], гетеростилия свойственна прежде всего видам умеренного климата и многолетним травам. Она реже встречается в другом климате и у иных жизненных форм. В филогенетическом плане она отсутствует в наиболее примитивных порядках цветковых растений и редко отмечается у однодольных [Агаджанян, 2000]. В локальной флоре Троицкого лесостепного заказника она зафиксирована только у 13 видов
(2.23% всех зарегистрированных здесь видов) и приурочена только к двудольным [Демьянова, 2011]. Нами была изучена структура популяций некоторых гетеростилъных растений, ранее не описанных в литературе, а также их антэкологические особенности.
Организация исследований
Наблюдения проведены в 1980-1981 гг. в природных популяциях Троицкого лесостепного заказника (Челябинская обл.) и его окрестностях. Предметом изучения явились диморфные виды (Limonium caspium (Willd.) Gams, L. gmelinii (Willd.) O. Kuntze, Goniolimon elatum (Fisch, ex Spreng.) Boiss. из сем. Limoniaceae) и триморфные Lythrum virgatum L. и L. salicaria L. (сем. Lythraceae). Наблюдения за Limonium caspium осуществлены на мокром солончаке, а за L. gmelinii - на корково-столбчатом солонце Соленого лога. Lythrum virgatum наблюдали на осоковых болотцах и солончаковых лугах, a L. salicaria - по ивнякам в долине р. Уй (в 20 км от заказника). Здесь же в каменистой степи на скалах исследовали Goniolimon elatum.
Определение структуры популяций производилось во время массового цветения видов путем визуального просмотра и подсчета морфологических типов. Учет растений проводился методом ходовых линий в нескольких ценопопуляциях, при этом фиксировались все растения подряд, независимо от их обилия [Пономарев, Демьянова, 1975; Демьянова, Пономарев, 1979]. Цветение и опыление растений изучалось по методике А.Н. Пономарева [1960]. Наблюдения сопровождались измерением температуры и влажности воздуха с помощью аспирационного психрометра Асмана. Семенная продуктивность определялась по методикам Т.А. Работнова [1960] и И.В. Вайнагия [1973, 1974]. Согласно последнему, семенная продуктивность исследовалась с раздельным определением потенциальной и реальной семенной продуктивности (ПСП и РСП). Под ПСП понимается количество семяпочек на цветок, а РСП - количество полноценных семян на элементарную единицу (цветок). Исследование фертильности пыльцы проведено методом микроскопирования в ацетокармине [Паушева, 1980]. Статистическая обработка материала выполнена согласно Г.Ф. Лакину [1980]. Определение насекомых сделано В.А. Лыковым, доцентом кафедры зоологии беспозвоночных Пермского университета.
-
I. Структура популяций гетеростильных растений
Подсчеты подтвердили известную из литературы закономерность об одинаковом или близком к одинаковому соотношению морфологических типов в природных популяциях гетеростильных видов (табл. 1). Подобное положение демонстрируют оба вида Limonium. Такая же ситуация отмечена нами ранее для L. sareptanum из заповедника Аскания-Нова [Демьянова, Надоль-ская, 1982].
|
У Goniolimon elatum нами отмечен морфизм пыльцы, на что ранее указ! тературе [Vuilleumier, 1967], но и бс диморфизм - разница в строении стол ках двух морфологических типов. С Соотношение мор( |
не только ди- тить, что у близкого в систематическом отношении явалось в ли- G. tataricum (L.) Boiss. в заповеднике Аскания-Нова лее глубокий также зафиксировано наличие длинно- и коротко-биков в цвет- столбчатых форм [Демьянова, Надольская, 1982]. ле дует отме- Таблица 1 юлогических форм у гетеростильных растений |
||||
|
Название растений |
Год исследования |
Морфологические формы, % |
Число исследованных растений |
||
|
длинно-столбчатая |
средне-столбчатая |
коротко-столбчатая |
|||
|
Limonium caspium |
1980 1981 |
48.0 48.98 |
0 0 |
52.0 51.02 |
1578 680 |
|
Limonium gmelinii |
1980 1981 |
51.0 43.67 |
0 0 |
49.0 56.33 |
6397 973 |
|
Goniolimon elatum |
1980 |
51.0 |
0 |
49.0 |
252 |
|
Lythrum virgatum |
1980 1981 |
35.60 37.20 |
32.30 9,36 |
32.10 53.44 |
1768 973 |
|
Lythrum salicaria |
1981 |
43.20 |
13.02 |
43.79 |
169 |
У тристильного Lythrum salicaria популяция в долине р. Уй в месте наблюдений весьма немногочисленна. Кроме того, ежегодно весной при подъеме уровня воды заливаются и местами уничтожаются отдельные группы особей. При малой выборке материала пока трудно судить об истинном соотношении морфологических форм у этого растения в лесостепном Зауралье. Из литературных источников известно, что нарушение структуры популяций у тристильных видов имеет место при случайном выпадении отдельных форм [Barrett, 1977; Eckert, Barrett, 1992; Barrett, Harder Lawrence, Cole, 2004].
Малой выборкой материала мы объясняем и резкое сокращение среднестолбчатой формы у Lythrum virgatum в 1981 г. Нужны дополнительные исследования с большей выборкой для интерпретации структуры популяций этих видов.
-
II. Антэкологические особенности гетеростильных видов
Дистильные виды
Limonium caspium (кермек каспийский) цветет с начала июня до середины июля. Limonium gmelinii (к. Гмелина) зацветает позже - с начала июля и заканчивает цветение в августе. Оба растения в обилии встречаются на засоленных почвах, давая красочный сиреневый (к. каспийский) и лиловый (к. Гмелина) аспекты.
Оба вида имеют четко выраженный утренний тип цветения (рис. 1-2), приуроченный к 7-8 ч. Распускание цветков у них происходит необычайно дружно: оно начинается на всех соцветиях одновременно и продолжается в течение 30 мин. -1ч. Пик распускания цветков приходится на 8 ч. В.Ф. Шаму-рин [1958] по аналогии с темпами распускания цветков у некоторых злаков [Пономарев, 1960] такое раскрывание цветков назвал «взрывчатым». Следует отметить, что подобный ход раскрывания цветков отмечен нами ранее и у других видов
Limonium Mill, в Центральном Казахстане - L. otolepis, L. chrysocomum, L. suffruticosum [Демьянова, 1976], а также у L. gmelinii. У перечисленных растений все цветки можно считать распустившимися одновременно в пределах получаса (много времени занимает непосредственный учет цветков, что трудно отметить в рис. 1-2).
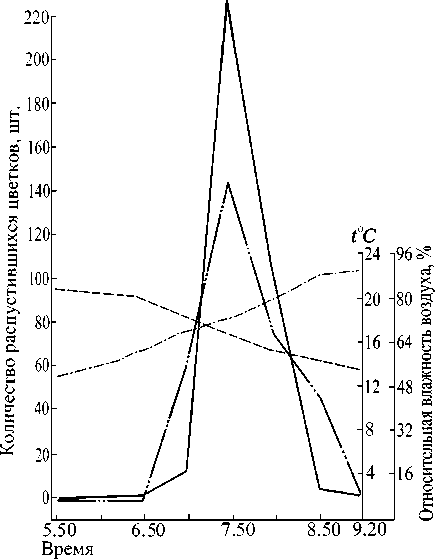
Рис. 1. Суточный ход распускания цветков у дистильного кермека каспийского 02.06.1978 г.:
-
- относительная влажность воздуха;
-
- температура;
-
- количество распустившихся короткостолбчатых цветков;
-
- количество распустившихся длинностолбчатых цветков
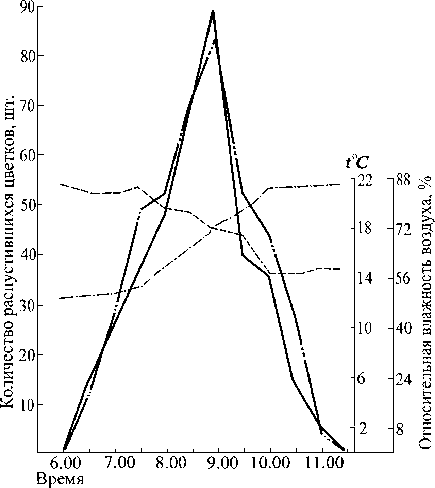
Рис. 2. Суточный ход распускания цветков у цистильного кермека Гмелина 07.07.1980 г.: ____- относительная влажность воздуха; ----- температура;
- - количество распустившихся короткостолбчатых цветков;
—— - количество распустившихся длинностолбчатых цветков
Нами замечено, что на распускание цветков большое влияние оказывает солнечный свет. Для изучения действия освещенности мы предприняли следующий опыт. Накануне вечером на модельные растения одевались колпаки из плотной светонепроницаемой бумаги. На следующее утро обнаружено, что растения, помещенные под колпаками не зацветали, хотя и находились в стадии бутона, в то время как другие растения в популяции оказывались в полном цвету. Отсутствие света при одинаковой температуре под светонепроницаемыми колпаками и вне его замедляло распускание цветков на 30-40 мин.
Судя по рис. 1, короткостолбчатая форма у кер-мека каспийского зацветает несколько раньше, и пе риод ее цветения в утренние часы более растянут. Однако максимум раскрывшихся цветков у обеих форм совпадает. У длинностолбчатой формы пик цветения выражен резче, и в целом эта форма характеризуется более кратковременным и дружным суточным периодом цветения.
У обеих форм ежедневно на генеративном побеге распускаются по 50-60 цветков. От начала раскрывания венчика до его полного распускания проходит около 30 мин. Тождественный ход распускания цветков у обеих морфологических форм подчеркивает их полную зависимость друг от друга при перекрестном опылении. Запаздывание хотя бы одной морфологической формы оказало бы неблагоприятное влияние на семенную продуктивность.
У всех видов кермека цветки недолговечны и увядают спустя 4-10 ч. после распускания. Продолжительность их жизни зависит от температурных условий. В жаркие дни увядание цветков происходит быстрее.
Цветки изученных видов кермека гомогамны. Все пыльники вскрываются интрорзно и одновременно. У только что раскрывшихся цветков пыльники и рыльца настолько близко располагаются друг от друга (практически контактируют), что, казалось бы, может привести к автогамии. Примерно через час со времени распускания цветков тычинки расходятся в стороны, располагаясь по окружности венчика. Ко второй половине дня венчики подсыхают, но не опадают, принимая голубовато-серую окраску у обоих видов. Увядая, они оказываются на уровне чашечки, оставаясь в таком состоянии до созревания плодов.
Цветки кермека каспийского, к. Гмелина и гониолимона высокого содержат высоко фертильную пыльцу (табл. 2). У двух последних мы не обнаружили разницы в фертильности пыльцы у обеих морфологических форм. Напротив, у к. каспийского длинностолбчатая форма характеризовалась более высокой фертильностью пыльцевых зерен в сравнении с короткостолбчатой, хотя эта разница и не столь значительна.
Таблица 2
Фертильность пыльцы у гетеростилъных растений в 1980 г.
|
Название растений |
Морфологическая форма |
Фертильность пыльцы |
Выборка материала |
|
Limonium caspium |
длинностолбчатая |
85.29 ± 4.27 |
30 |
|
короткостолбчатая |
77.84 ±4.58 |
30 |
|
|
Limonium gmelinii |
длинностолбчатая |
90.25 ± 7.20 |
25 |
|
короткостолбчатая |
89.71 ± 10.50 |
25 |
|
|
Goniolimon datum |
длинностолбчатая |
95.07 ± 9.80 |
30 |
|
короткостолбчатая |
96.15 ±8.90 |
30 |
|
|
Lythrum virgatum |
длинностолбчатая |
93.90 ±7.80 |
25 |
|
среднестолбчатая |
94.70 ± 8.50 |
25 |
|
|
короткостолбчатая |
91.50 ±6.91 |
25 |
Сразу же после распускания цветков кермека их энергично посещают насекомые (до 13-14 ч.). В числе насекомых-опылителей, посещающих обе формы цветков, зарегистрированы шмели, дикие пчелы, небольшие серые мушки. К сожалению, последние не были определены. Что касается пчелиных, то к. Гмелина посещали лучше, чем к. каспийский. Первый представляет собой более крупное растение с яркими, хорошо заметными соцветиями. К. каспийский, вероятно, менее заметен для насекомых. На цветках к. Гмелина зарегистрированы Andrena thoracica F., A. flavipes Pz., Nomia femoralis Pallas, N. diversipes Latr., Bombus lucorum L., B. muscorum F., B. equestris F., B. terrestris L., B. distinguendus F. Mor., а на к. каспийском - Nomia diversipes Latr., N. femoralis Pallas, Bombus sichelii Rad., B. lucorum L., B. terrestris L.
Высокая фертильность пыльцы и интенсивное посещение насекомыми цветков к. Гмелина способствуют хорошему завязыванию плодов при свободном опылении (табл. 3). Напротив, более слабое посещение насекомыми к. каспийского, возможно, приводит к худшему в сравнении с к. Гмелина завязыванию плодов у обеих морфологических форм. У гониолимона высокого отмечено почти одинаковое завязывание плодов у обеих форм.
Таблица 3
Процент плодоцветения при свободном опылении и изоляции диморфных растений в 1980 г.
|
Название растений |
Морфологическая форма |
Завязывание плодов |
|
свободное опыление |
||
|
Limonium caspium |
длинностолбчатая короткостолбчатая |
73.20 ±5.30 75.30 ±4.70 |
|
Limonium gmelinii |
длинностолбчатая короткостолбчатая |
97.40 ± 7.22 96.10 ±5.04 |
|
Goniolimon elatum |
длинностолбчатая короткостолбчатая |
96.81 ±7.30 95.30 ±6.70 |
|
Limonium caspium |
изоляция длинностолбчатая короткостолбчатая |
13.60 ± 1.30 23.10 ± 1.90 |
|
Limonium gmelinii |
длинностолбчатая короткостолбчатая |
12.90 ± 1.40 15.70 ± 1.25 |
При изоляции соцветий у обоих видов кермека завязались плоды, хотя и в гораздо меньшем числе в сравнении с вариантом свободного опыления (табл. 3). Вероятно, при неудавшемся перекрестном опылении реакция самонесовместимости делается слабой, и потому становится возможным самоопыление (гейтоно- и автогамия). При завядании венчик свертывается, напоминая бутон, что приводит к соприкосновению пыльников с остатками пыльцы и рыльца. Возможность самоопыления известна и у других видов сем. Limoniaceae (например, Armeria maritime) [Lefebvre, 1970, 1976].
Тристильные виды
Lythrum virgatum - дербенник прутовидный.
Изучение суточного ритма раскрывания цветков показало, что их распускание у всех трех морфологических типов начинается одновременно -около 6 ч. (рис. 3). Более того, максимум раскрывания у них приходится также на одно время -8 ч. В течение суток распускание цветков продолжается до 12 ч., после чего прекращается также одновременно у всех морфологических форм. Такая удивительно точная синхронизация в суточном ритме цветения всех флоральных типов, по аналогии с дистальными формами изученных видов кермека и гониолимона, подразумевает их полную зависимость друг от друга и облегчает перекрестное опыление.
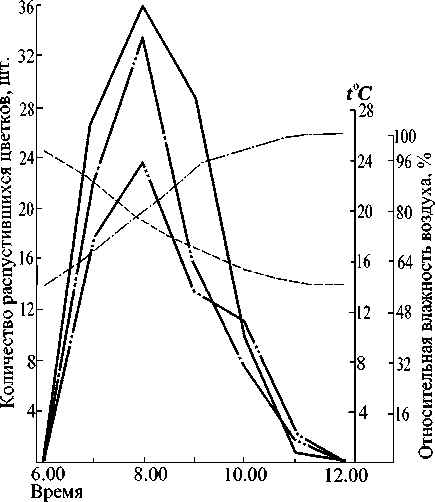
Рис. 3. Суточный ход распускания цветков у тристильного дербенника прутьевидного 12.07.1980 г.:
______ - относительная влажность воздуха;
-------температура;
- - ----- - - количество распустившихся короткостолбчатых цветков;
_ _____ _ ' - количество распустившихся среднестолбчатых цветков;
- количество распустившихся длинностолбчатых цветков
Наблюдения за экологией цветения отдельных цветков позволили констатировать у них гомогамию. В бутонах все пыльники одного цвета - слегка розоватые. По мере раскрывания цветков их окраска изменяется. Короткие и средние тычинки у всех форм имеют ярко-желтые пыльники с такого же цвета пыльцой. У длинных тычинок пыльники окрашены в серовато-зеленоватый цвет. Такого же цвета и содержащаяся в них пыльца. Все пыльники тычинок разного уровня вскрываются интрорзно и одновременно при распускании цветков. Пыльца в них сохраняется 5-6 ч. Рыльца становятся воспри имчивыми к пыльце спустя 20-30 мин. после раскрывания цветков. У длинностолбчатой формы рыльце выдвигается быстрее, чем у других форм.
Растения разных морфологических форм различаются по сосочковой поверхности рылец. Более длинные сосочки имеются у короткостолбчатой формы, а слабее всего выражена сосочковая поверхность у длинностолбчатой формы. Среднестолбчатая форма занимает промежуточное положение. Размеры пестиков и тычиночных нитей детерминированы у всех трех флоральных типов (табл. 4).
Таблица 4
Размеры пестиков и тычиночных нитей (в мм) в цветках разных морфологических форм у Lythrum virgatum (n = 30)
|
Морфологическая форма |
Длина пестика (завязь ± столбик) |
Длина тычинок |
||
|
длинные |
средние |
короткие |
||
|
длинностолбчатая |
9.05 ±0.01 |
5.17 ±0.03 |
3.00 ±0.08 |
|
|
среднестолбчатая |
6.71 ±0.03 |
9.00 ±0.01 |
- |
3.00 ±0.02 |
|
короткостолбчатая |
4.37 ±0.03 |
9.00 ±0.01 |
5.10 ±0.02 |
- |
Продолжительность жизни цветков всех морфологических типов одинакова - 34-36 ч. Обращает на себя внимание особая «экономность» в цветении особей: на одном генеративном побеге каждый день раскрывается не более 3-4 цветков. Эго приводит к лучшей обеспеченности перекрестного опыления, увеличивая продолжительность цветения вида и почти одинаковой длительности цветения разных форм. Эти приспособления в генеративной сфере лишний раз подчеркивают тесную зависимость между отдельными флоральными формами. Успешное образование семян полностью определяется одновременностью в прохождении всех этапов цветения особей и цветков разных морфологических типов.
Определение фертильности пыльцы дербенника прутовидного показало ее высокое качество для вида в целом (табл. 5) и для отдельных форм. У последних не обнаружено и значительной разницы между флоральными формами, хотя у коротко-столбчатой формы этот показатель несколько выше.
Таблица 5
Фертильность пыльцы разных морфологических типов тычинок у тристильного Lythrum virgatum в 1980 г. (п = 30)
|
Морфологическая форма цветка |
Фертильность пыльцы, % |
||
|
длинные тычинки |
средние тычинки |
короткие тычинки |
|
|
длинностолбчатый |
89.78 ±3.15 |
91.41 ±2.34 |
|
|
среднестолбчатый |
89.77 ± 1.88 |
- |
89.58 ±3.23 |
|
короткостолбчатый |
93.13 ±2.17 |
93.03 ± 1.15 |
- |
Опылители на цветках д. прутовидного появляются в утренние часы сразу же после раскрывания цветков. Эго дикие пчелы Andrena marginata F. и Megachile analis Nyl., шмели Bombus sichelii Rad., B. terrestris L., B. lucorum L., B. muscorum F. Насекомые посещают цветки этого растения в течение всего светлого времени суток.
Изучение некоторых элементов семенной продуктивности показало, что наибольшее завязывание семян при свободном опылении обнаружено у длинностолбчатых форм, а наименьшее - у короткостолбчатых, хотя разница между всеми флоральными формами невелика. Число цветков на генеративных побегах практически одинаково (табл. 6).
Таблица 6
Некоторые показатели семенной продуктивности Lythrum virgatum при свободном опылении в 1980 г. (п = 60)
|
Показатели |
Морфологические формы, % |
||
|
длинностолбчатая |
среднестолбчатая |
короткостолбчатая |
|
|
ПСП в расчете на цветок РСП в расчете на цветок Число цветков на побеге |
121 ±7.35 112 ± 13.50 73.50 ±5.09 |
116 ±8.20 110± 15.50 66 ± 4.60 |
112 ±6.80 107.80 ± 10.5 70.50 ±3.50 |
Примечание. ПСП - потенциальная семенная продуктивность; РСП - реальная семенная продуктивность.
При изоляции соцветий марлевыми мешочками наблюдали чрезвычайно малое количество плодов и семян (в немногих плодах присутствовало по 1015 семян). В литературе д. прутовидный рассматривается как перекрестноопыляемый само несовместимый вид [Fryxell, 1957]. По-видимому, в условиях изоляции реакция самонесовместимости сдвигается в сторону частичной самосовместимо-сти, и при попадании пыльцы с соседних цветков этого растения (у длинностолбчатой формы) или даже собственных цветков (у короткостолбчатой и среднестолбчатой формы) становится возможным завязывание семян.
Таким образом, в условиях лесостепного Зауралья не обнаружено особых нарушений в структуре популяций дистильных кермека каспийского, к. Гмелина, гониолимона высокого и тристильно го дербенника прутовидного. Впервые в литературе у гониолимона высокого обнаружен не только пыльцевой диморфизм, что было известно ранее, но и разница в длине столбиков. Что касается тристиль -ного дербенника иволистного, необходимы дополнительные наблюдения по структуре популяций. В суточной ритмике цветения энтомофильных кермека каспийского и к. Гмелина обнаружено стремительное, «взрывчатое» распускание цветков, напоминающее раскрывание цветков у некоторых ветроопыляемых злаков. Феномен этого явления мы не можем расшифровать. Активное посещение насекомыми-опылителями исследованных гетеро-стильных растений приводит к успешной семенной продуктивности при свободном опылении.
Список литературы О полиморфизме некоторых гетеростильных растений лесостепного Зауралья
- Агаджанян А.М. Распространённость и распределение гетеростилии в системе покрытосеменных//Успехи современной биологии. 2000. Т. 120, № 4. С. 348-360.
- Вайнагий И.В. Методика статистической обработки материала по семенной продуктивности растений на примере Potentilla aurea L.//Растительные ресурсы. 1973. Т. 9, № 2. С. 287-296.
- Вайнагий И.В. О методике изучения семенной продуктивности растений//Ботанический журнал. 1974. Т. 59, № 6. С. 826-831.
- Дарвин Ч. Различные формы цветов у растений одного и того же вида//Собр. соч. М.; Л., 1948. Т. 7. С. 35-251.
- Демьянова Е.И. К антэкологии растений пустынных степей Центрального Казахстана//Экология опыления: межвуз. сб. науч. тр. Пермь, 1976. Вып. 2. С. 63-78.
- Демьянова Е.И. Спектр половых типов и форм в локальных флорах Урала (Предуралья и Зауралья)//Ботанический журнал. 2011. Т. 96, № 10. С. 1297-1315.
- Демьянова Е.И., Надольская Е.И. К изучению полового полиморфизма растений целинной степизаповедника Аскания-Нова//Экология опыления растений: межвуз. сб. науч. тр. Пермь, 1982. С. 70-79.
- Демьянова Е.И., Пономарев А.Н. Половая структура природных популяций гинодиэцичных и двудомных растений лесостепи Зауралья//Ботанический журнал. 1979. Т. 64, № 7. С. 1017-1024.
- Лакин В.Ф. Биометрия. М.: Высш. школа, 1980. 293 с.
- Паушева З.П. Практикум по цитологии растений. М., 1980. 304 с.
- Пономарев А.Н. Изучение цветения и опыления растений//Полевая геоботаника. М.; Л.: Наука, 1960. Т. 2. С. 9-19.
- Пономарев А.Н., Демьянова Е.И. К изучению гинодиэции у растений//Ботанический журнал. 1975. Т. 60, № 1. С. 3-15.
- Работнов Т.А. Методы изучения семенного размножения травянистых растений в сообществах//Полевая геоботаника. М.; Л.: Изд-во АН СССР, 1960. Т. 2. С. 20-40.
- Фегри К., Л. ван дер Пэйл. Основы экологии опыления. М.: Мир. 1982. 381 с.
- Шамурин В.Ф. Суточная ритмика и экология цветения некоторых степных растений//Ботанический журнал. 1958. Т. 43, № 4. С. 548-557.
- Agren J. Population size, pollinator limitation, and seed set in the self-incompatibile herb Lythrum salicaria//Ecology. 1996. Vol. 77, № 6. P. 1779-1790.
- Arasu N.P. Self-incompatibility in Angiosperm: a review//Genetica. 1968. Vol. 39, № 1. P. 1-24.
- Baker H.G. Race formation and reproductive method in flowering plants//Symp. Soc. Exp. Biol. 1953a. Vol. 7. P. 114-143.
- Baker H.G. Dimorphism and monomorphism in the Plumbaginaceae. 3. Correlation of geographical distribution patterns with dimorphism and monomorphism in Limonium//Ann. Bot. (N.S.). 1953b. Vol. 17, № 68. P. 614-615.
- Baker H.G. Evolutionary mechanism in pollination biology//Science. 1963. Vol. 139, № 3558. P. 877-883.
- Baker H.G. The evolution of floral heteromorphism and gynodioecism in Silene maritima//Heredity. 1966. Vol. 21, № 4. P. 689-692.
- Barrett Spencer C.H. Tristyly in Eichhornia crassipes (Mart.) Solms (Water Hyacinth)//Biotropica. 1977. Vol. 9, № 4. P. 230-238.
- Barrett Spencer C.H., Harder Lawrence D., Cole W.W. Correlated evolution of floral morphology and mating-type frequencies in a sexually polymorphic plant//Evolution (USA). 2004. Vol. 58, № 5. P. 964-975.
- Crowe L.K. The evolution of outbreeding//Heredity. 1964. Vol. 19, part. 3. P. 435-457.
- Darwin Ch. The different forms on plants on the same species. London: John Murray, 1877. 352 p.
- Eckert Ch., Barrett S. Stochastic loss of style morphs from populations of tristylous Lythrum salicaria and Decodon verticillatus (Lythraceae)//Evolution (USA). 1992. Vol. 44, № 4. P. 1014-1029.
- Fryxell P.A. Mode of reproduction of higher plants//Bot. Rev. 1957. Vol. 23, № 3. P. 135-233.
- Ganders F.R. The biology of heterostyly//New Zealand J. of Bot. 1979. Vol. 17. P. 607-635.
- Lefebvre C. Self-fertility in maritime and zinc mine populations of Armeria maritima (Mill.) Willd.//Evolutiom (USA). 1970. Vol. 24, № 3. P. 571-577.
- Lefebvre C. Breding system and population structure of Armeria maritima (Mill.) Willd. On a zinc-lead mine//New Phyt. 1976. Vol. 77, № 1. P. 187-192.
- Lewis D. Incompatibility in flowering plants//Biol. Rev. 1949. Vol. 24, № 4. P. 472-496.
- Lewis D. Genetic versatility of incompatibility in plants//New Zealand J. Bot. 1979. Vol. 17, № 4. P. 637-644.
- Nettancourt D. de. Incompatibility in Angiosperms. N.Y.: Springer, 1977. 230 p.
- Ornduff R. Intramorph and intermorph compatibility in tristylous population of Decodon verticillatus (Lythraceae)//Bull. Torrey Bot. Club. 1993. Vol. 120, № 1. P. 19-22.
- Vuilleumier B.S. The origin and evolutionary development of heterostyly in the Angiosperms//Evolution. 1967. Vol. 21, № 6. P. 210-225.
- Waites Anna R., Agren J. Pollinator visitation, stigmatic pollen loads and among-population variation in seed set in Lythrum salicaria//J. Ecol. 2004. Vol. 92, № 3. P. 512-526.
- Yeo P.F. Some aspects of heterostyly//New Phyt. 1975. Vol. 75, № 1. P. 147-153.
