О популяционной дифференциации гуменника ( Anser fabalis) при миграционных перераспределениях на юге России
 при миграционных перераспределениях на юге России")
Автор: Линьков А.Б., Воробьев В.Н., Шалаев А.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Охотоведение, динамика популяций
Статья в выпуске: 6 т.47, 2012 года.
Бесплатный доступ
На основе анализа генетической изменчивости индивидуальной митохондриальной ДНК (мтДНК) обсуждается экоморфизм и популяционная принадлежность гуменников, мигрирующих на юге России. Оценивается популяционная дифференциация вида в целом, рассматриваются проблемы его охраны и научно-методическая обоснованность регламентации гусиной охоты.
Миграции, гуменник, популяционные дифференциации, миграционный мониторинг, митохондриальная днк, миграционные перераспределения, популяционные арены
Короткий адрес: https://sciup.org/142133355
IDR: 142133355 | УДК: 639.1.052:575.17:577.21.06:599.735.34(470.45)
About population variability of bean goose ( Anser fabalis) at the migration over distribution in the south of Russia
In the paper, an ecomorphism, a geographic variability and population origin of bean gooses during their migration in the South of Russia are discussed. A few genetic features of the bean goose populations were considered. On the basis of analysis of nucleotide sequences of mitochondrial DNA (mtDNA) haplotypes the authors estimated the genetic differentiation in forms (subspecies) of bean goose. The problems of conservation of the bean goose species in whole, and a methodological validity of regulation of goose hunting are under consideration.
Текст научной статьи О популяционной дифференциации гуменника ( Anser fabalis) при миграционных перераспределениях на юге России
Инвентаризация популяций охотничьих животных с использованием анализа мтДНК и развитие теории «популяционных арен» имеют не только филогенетическое значение, но и важнейшее практическое применение в охотоведении (1-3). Длительное время пернатая дичь, к сожалению, считалась неисчерпаемым ресурсом и в научном охотоведении ей не уделялось должного внимания. Именно поэтому реализация программы инвентаризации началась на популяциях лосей, на что, в основном, и выделялись значительные средства. Результатом такой концентрации ресурсов стало появление знаковой для охотоведения книги, посвященной лосю (3). То же обстоятельство, к сожалению, существенно сдерживало исследования популяционной дифференциации массовых видов пернатой дичи. Тем не менее, общее число появившихся публикаций, в которых с использованием мтДНК-тестирования анализировали принадлежность птиц разных систематических групп к тем или иным разноранговым популяциям, или посвященных их филогенетике, в зарубежной и отечественной литературе с конца 1990-х годов значительно возросло (4-8).
Гуменник — политипичный вид, разноранговая популяционная структурированность которого представляет собой весьма красноречивый результат экоморфной эволюции. Современный экоморфизм этого вида (экоморфный облик), как нам представляется, сложился прежде всего в результате адаптаций к внутривидовой и даже межвидовой конкуренции в миграционный период (многовидовые миграционные скопления гусей) и на зимовках, возникших в плейстоцене. Чередующиеся периоды оледенений в Евразии (в какой-то мере разнофазные в силу больших различий условий в ее западных и восточных частях) то открывали, то закрывали порталы в огромные репродуктивные пространства на севере материка, одновременно включая или, наоборот, ограничивая механизмы миграционного адаптогенеза, направленного на снижение внутривидовой и межвидовой конкуренции, что привело к современному глубоко структурированному в популяционном аспекте характеру пролета. В период миграций подобная структурированность внешне проявляется, как правило, в выраженной волнообразности пролета наряду с территориальной дифференциацией этих структур на присадах и так называемых миграционных станциях. Именно популяционный миграционный мониторинг гуменника, впрочем, как и двух других массово промышляемых видов гусей — белолобого и серого, позволит оптимизировать регламентацию гусиной охоты, которую сейчас вряд ли можно назвать научно обоснованной.
Нашей целью было изучение популяционной дифференциации гуменника в миграционный период на основе сравнения митохондриальной ДНК.
Методика . Для сбора проб был выбран район юго-восточной границы предположительно второй евразийской популяционной арены гуменника, выделенной нами (1), что территориально соответствует Волгоградской области. Одновременно эта территория представляет собой часть большой миграционной станции гусей в так называемом Срединном регионе (8-10), откуда в периоды сезонных миграций, пополнив запасы жира, гуси, преимущественно белолобый и гуменник, перелетают к следующим станциям, расположенным на расстоянии 800-1000 км, где на них ведется интенсивная охота.
Праймеры подбирали таким образом, чтобы можно было ампли-фицировать 5'-конец контрольного региона CRI митохондриального генома Anser fabalis (нуклеотидные позиции 154-429, депонировано в GenBank как Anser fabalis rossicus AF 159951). Использованные праймеры позволяют амплифицировать фрагмент размером 274 п.н. с митохондриальной ДНК (мтДНК) всех подвидов гуменника. Амплифицированный фрагмент содержит 13 вариабельных сайтов, что составляет почти половину значимых полиморфизмов полного контрольного региона CRI митохондриального генома размером 1164 п.н. (29 вариабельных сайтов). Методы выявления гаплотипов и первичной компьютерной обработки подробно изложены в работах Н.В. Кол с соавт. (11) и М. Ней и С. Кумар (12).
При филогенетической кластеризации в число анализируемых дополнительно включали последовательности гаплотипов BRA1, BRA2, BRA6 ( Anser brachyrhynchus ), MID1, MID2, MID4, MID5, MID6 ( Anser fabalis middendorffii ), FAB1, FAB3, FAB6 ( Anser fabalis fabalis ), ROS2, ROS3, ROS4 ( Anser fabalis rossicus) , SER2, SER4 ( Anser fabalis serrirostris ) и общий для тундровых подвидов гаплотип SER1 (номера в базе данных GenBank — EU186813-EU186828, AY552063, AY552064, AY552068).
Резуёътаты . Мы проанализировали популяционную дифференциацию 41 особи, что составляет лишь небольшую часть банка проб, собранных нами в последнее десятилетие.
Учитывая наличие копии митохондриального генома в ядерной ДНК у птиц рода Anser , праймеры выбирали в области с нуклеотидными различиями между последовательностями митохондриальной и ядерной ДНК ( Anser fabalis rossicus — AF 159965, Anser anser anser — AF 159970, Anser brachyrhynchus — AF 159966), что позволило избежать амплификации хромосомной ДНК и искажения результатов исследования.
Установлено, что степень различий между нуклеотидными последовательностями колебалась от 0 % (один гаплотип) до 3,8 % (образцы AN29 гаплотипа MID6 и AN31 гаплотипа ROS4). На основании анализа исследуемых образцов были выявлены четыре группы, которые таксономически совпадают с подвидам гуменника (рис. 1).
I группа соответствует подвиду западного тундрового гуменника Anser fabalis rossicus и включает подгруппы:
Та — последовательности AN2, AN4, AN5, AN6, AN7, AN13, AN14, AN15, AN17, AN18, AN19, AN21, AN27, AN30, AN32, AN33, AN34, AN35, AN38, AN39, AN40, AN41, AN42 соответствуют последовательности EU186826 (Anser fabalis гаплотип ROS2, Vaygach VY3 Russia); последовательность AN23 также принадлежит к этой группе, хотя отличается от гаплотипа ROS2 одной нуклеотидной заменой;
I6 — последовательности AN9, AN26, AN28, AN37 соответствуют последовательности EU186827 ( Anser fabalis гаплотип ROS3, Chornaya H19 Russia);
1в — последовательность AN31 соответствует последовательности EU186828 ( Anser fabalis гаплотип ROS4, Krasnoyarsk MO1 Russia, предположительно подвид rossicus ).
AN13
AN39
AN42
AN15
AN41
AN21
AN6
AN23
AN18
AN7
AN14
AN19
AN38
ROS2
AN2
AN5
AN30
AN35
AN27
AN36
AN40
AN17
AN32 AN34 AN33
AN4
87 ROS4
AN31
AN28
AN37
AN26
ROS3
AN9
SER4
AN20
AN8
AN24
AN12
AN25
AN16
AN1
AN10
SER1
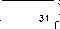
SER2
SER3
FAB3
30 FAB1
AN22 (Московская область)
45 FAB6
MID3
AN29 (Новосибирская область) 65
MID6
MID2
MID5
MID1
MID4
BRA2
95 BRA1
81 BRA6
Рис. 1. Дендрограмма, полученная на основании филогенетического анализа 5'-концевого фрагмента из контрольного региона CRI митохондриальной ДНК у изученных гуменников.
II группа несет последовательности гаплотипа SER1, общего для тундровых подвидов гуменника serrirostris и rossicus . Она представлена последовательностями AN1, AN8, AN10, AN12, AN16, AN20, AN24, AN25, которые на 100 % соответствуют EU186822 ( Anser fabalis гаплотип SER1, Yamal N5 Russia).
-
III группу составляют птицы, относящиеся к подвиду западного лесного (таежного) гуменника Anser fabalis fabalis и имеющие последовательность AN22, соответствующую (100 %) последовательности EU186819 (гаплотип FAB1, Pudasjarvi-S28-Finland).
-
IV группа соответствует виду сибирского таежного гуменника Anser middendorffii . У выявленной у этой группы последовательности AN29 наблюдается полное (100 %) совпадение с EU186818 ( Anser middendorffii гап-124
лотип MID6, Tomsk TO6 Russia).
Таким образом, по результатам исследования 41 образца тканей гуменника один (AN29) принадлежит к виду Anser middendorffii (сибирский таежный гуменник), один (AN22) — к подвиду Anser fabalis fabalis (западный лесной, или таежный, гуменник). Наибольшее число образцов (всего 29) относятся к подвиду Anser fabalis rossicus (западный тундровый гуменник), 8 — имеют гаплотип SER1, общий для тундровых подвидов гуменника serrirostris и rossicus .
Исходя из морфологических данных и полученных результатов мо лекулярно-генетического анализа, гуменника можно подразделить на три крупные экоморофные группы, или экологические формы — макроэкотипы. Очевидно, вслед за А.Я. Тугариновым (13), M. Ruokonen с соавт. (14) и некоторыми другими исследователями необходимо признать видовую самостоятельность короткоклювого гуменника (Anser brachyrhynchus). Весьма обособлен и восточный таежный гуменник, по Л.С. Степаняну (15) — Anser fabalis middendorffii (или, что вполне вероятно, Anser middendorffii). Обе эти экоморфы, по имеющимся сведениям, демонстрируют низкую внутривидовую вариабельность генома. В свою очередь, третья экоморфа, наоборот, включает три вполне экоморфичных подвида — западного таежного (лесного) гуменника Anser fabalis fabalis, обитающего в таежной зоне Европы и Западной Сибири вплоть до Енисея, тундрового гуменника Anser fabalis rossicus с ареалом на северо-востоке Европы и на севере Западной
Сибири и репродуктивным ареалом, заходящим на Таймыр, и восточного тундрового гуменника Anser fabalis serrirostris, обитающего от восточного
Таймыра до севера Восточной Сибири.
Ранее мы схематично очертили арены основных популяций гумен ника (рис. 2), выделяемые в настоящее время (1).
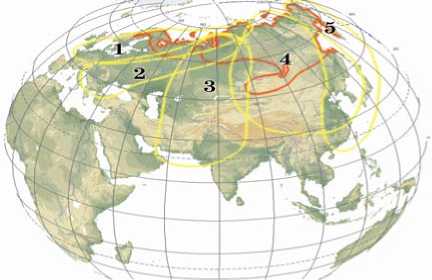
Рис. 2. Популяционные арены (1-5) гуменника в Евразии по А.Б. Линькову (1).
Исходя из полученных нами данных, вторая популяционная арена, границы которой требуют уточнения, по-видимо-му, включает преимущественно группы гуменника rossicus и популяции с гаплотипом SER1, общим для rossicus и serrirostris. Специально включенные в анализ европейские и сибирские пробы (AN22 и AN29) позволили еще раз продемонстрировать отсутствие fabalis fabalis и fabalis middendorffii (таежных гу менников) в миграционных потоках в юго-западной области этой арены. Вместе с тем подтверждается и общая схема миграционного перераспределения разноранговых популяций гуменника в разработанной нами модели миграционного перераспределения гусей в Срединном регионе, куда мы включили и Таймыр (16).
Итак, впервые на основе анализа мтДНК гуменников, мигрирую щих на юге европейской части России, выявлена их принадлежность к двум популяционным группировкам — западно-тундровой (rossicus) и гибридной (rossicus-serrirostris). Использование результатов количественно-популяционного миграционного мониторинга при регламентации гусиной охоты в южных регионах России наряду с оценкой успешности размноже- ния той или иной популяции и ее состояния на зимовках позволит сохранить популяционное разнообразие этого важного объекта, а значит, и саму гусиную охоту.
Л И Т Е Р А Т У Р А
-
1. Линьков А.Б. Охотничьи водоплавающие птицы России. М., 2002.
-
2. Линьков А.Б., Железнов-Чукотский Н.К. Научные направления в развитии государственного управления охотничьими ресурсами России (концепция). Мат. XXIX Межд. конгр. биологов-охотоведов. М., 2009, ч. 1: 141.
-
3. Рожков Ю.И., Проняев А.В., Давыдов А.В., Холодова М.В., Сипко Т.П. Лось: популяционная биология и микроэволюция. М., 2009.
-
4. Double M.C., P eakall R., Beck N.R., Cockburn A. Dispersal, philopatry, and infidelity: Dissecting local genetic structure in superb fairy-wrens ( Valurus cyaneus ). Evolution (USA), 2005, 59(3): 625-635.
-
5. Liukkonen T. Finnich native grey partridge ( Perdixperdix ) population differs clearly in mitochondrial DNA from the farm stock used for releases. Ann. Zool. Fenn., 2006, 43(3): 271-279.
-
6. Ruokonen M. Genetic composition of the captive Lesser White-fronted Goose population. Fennoscandian Lesser White-fronted Goose conservation project Annual report 1999. WWF Finland Report No 12, NOF Rapportserie Report No 1. 2000: 54-56.
-
7. Popovkina A.B., Poyarkov N.D. History of phylogenetic and taxonomic studies of Anseri-formes . Casarca . Bulletin of goose, swan and duck study group of Northern Eurasia (English Supplement 1), 2005: 19-51.
-
8. Давыдов А.В., Холодова М.В., Мещерский И.Г., Груздев А.Р., Сипко Т.П., Кол Н.В., Царев С.А., Железнов-Чукотский Н.К., Мирутен-ко В.С., Губарь Ю.П., Линьков А.Б., Рожков Ю.И. Дифференциация диких и домашних форм северного оленя ( Rangifer tarandus L.) по результатам анализа мтДНК. С.-х. биол., 2007, 6: 48-53.
-
9. Гаврин В.Ф., Кривенко В.Г., Иванов Г.К., Азаров В.И., Молочаев А.В., Линьков А.Б., Антипов А.М., Дебело П.В., Стопалов В.С. Особенности весеннего пролета водоплавающих птиц в Срединном регионе СССР. В сб. науч. тр. ЦНИЛ Главохоты РСФСР: Экология и охрана охотничьих птиц. М., 1980: 5-45.
-
10. Кривенко В.Г., Иванов Г.К., Азаров В.И., Молочаев А.В., Линьков А.Б., Антипов А.М., Дебело П.В., Стопалов В.С., Лысенко И.Г. Особенности осеннего пролета, размещение и численность водоплавающих птиц в Срединном регионе СССР. В сб. науч. тр. ЦНИЛ Главохоты РСФСР: Экология и охрана охотничьих птиц. М., 1980: 65-96.
-
11. Кол Н.В., Королев А.Л., Захаров И.А. Полиморфизм митохондриальной ДНК в тувинской популяции северного оленя ( Rangifer tarandus L.). Генетика, 2006, 42(1): 110-112.
-
12. Ней М., Кумар С. Молекулярная эволюция и филогенетика. Киев, 2004.
-
13. Тугаринов А.Я. Птицы СССР (утки, гуси, лебеди и крохали). Л., 1932.
-
14. Ruokonen M., Litvin K., Aarvak T. Taxonomy of the bean goose-pink-footed goose complex. Mol. Phylogenet. Evol., 2008, 48: 554-562.
-
15. Степанян Л.С. Конспект орнитологической фауны СССР. М., 1990.
-
16. Кривенко В.Г., Линьков А.Б. Оптимизация охоты на водоплавающих птиц и модели «движения численности» как основы нормирования добычи. Вопросы интенсификации охотничьего хозяйства. Тр. ЦНИЛ Главохоты РСФСР (М.), 1988: 160-169.
ФГБОУ ВПО Российский государственный аграрный заочный университет,
