О проблеме сопряженного биогеографического анализа разных компонентов биоты

Автор: Нотов Александр Александрович
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Биоразнообразие и охрана природы
Статья в выпуске: 14, 2009 года.
Бесплатный доступ
Обсуждается проблема сопряженного биогеографического анализа разных компонентов биоты. В качестве модельных объектов рассмотрены флоры сосудистых растений и мхов, биоты лишайников и миксомицетов Валдайской и смежных физико-географических провинций Русской равнины. Выявлена специфика изменения основных параметров флор и биот в долготном и широтном направлениях. Общий уровень богатства, специфичность и степень сходства видового состава анализируемых компонентов зависят в разной степени от климатических особенностей, широтной дифференциации растительного покрова, геоморфологии. Разная структура широтных и долготных трендов обусловлена спецификой биологии и экологии таксономических групп.
Биогеографический анализ, биота, флора, сосудистые растения, мхи, лишайники, миксомицеты, уровень видового богатства флор, дифференциальные виды, специфический компонент флор и биот, русская равнина, сопряженный анализ разных компонентов биоразнообразия
Короткий адрес: https://sciup.org/146116243
IDR: 146116243 | УДК: 574
To the problem of integrated biogeographic analysis of various component of biota
Problem of integrated biogeographic analysis of various components of biota is discussed. Florae and biotae of vascular plants, lichens and myxomicetes of Valdai and adjacent physicogeographical provinces of Russian Plain are treated as model objects. Peculiarity of changes in florae and biotae in longitudinal and altitudinal directions is revealed. General level of richness, specificity and degree of similarity of specific composition of analyzed components depend to various degrees on climatic conditions, altitudinal differentiation of plant as well as on geomorphology. Different structure of altitudinal and longitudinal trends is due to peculiarities in biology and ecology of taxonomic groups.
Текст научной статьи О проблеме сопряженного биогеографического анализа разных компонентов биоты
Основы биогеографического анализа разработаны преимущественно на материале по сосудистым растениям и позвоночным животным [12; 18; 19; 22; 39; 47; 48; 62; 65; 66; 83 и др.]. Полученные данные стали базой для флористического и фаунистического районирования биоты Земли и крупных биогеографических единиц [1; 20 – 22; 39; 45; 68; 69; 75]. В настоящее время сравнительная флористика позволяет выявлять закономерности территориального распределения флор [71; 72; 57; 58 и др.]. Формируется концепция мониторинга биоразнообразия на уровне локальных флор сосудистых растений, выясняются возможности ее практического использования [40; 70; 72 – 74], разрабатываются методы картографирования биоразнообразия [36]. Пространственное распределение других компонентов биоты имеет свою специфику, которая обусловлена особенностями биологии и экологии представителей разных царств и таксонов высокого ранга. На примере мхов было показано, что в отличие от сосудистых растений, основные параметры их флор в большей степени зависят от фактора влажности воздуха, чем от температурного режима [77; 78]. Первый фактор является более значимым и для лишайников [50 – 52; 76; 80]. Для некоторых групп организмов зональные и секторальные особенности растительного покрова, климата играют меньшую роль, чем характеристики конкретных экотопов и микрониш. Микрониши могут быть достаточно универсальными и встречаться относительно независимо от широты и долготы на территориях большого размера. Среди таких групп миксомицеты [26 – 28], некоторые микроорганизмы [8; 23; 59 – 61; 63]. Развитие основ биогеографического анализа применительно к водорослям, лишайникам, печеночникам, микроскопическим грибам и другим компонентам биоты сдерживает недостаточная изученность флор и биот многих систематических групп. Традиционное выделение географических элементов сопряжено с определенными трудностями, которые обусловлены особенностями биологии, экологии, спецификой таксономической эволюции и генезиса флор и биот [13; 14; 17; 27; 28; 37; 38; 49; 51; 64; 77].
Проблему сопряженного анализа разных компонентов биоты стали обсуждать только в конце XX в. [43; 77; 78]. Разработка этой проблемы имеет большое общебиологическое значение. Выяснение трендов географического распределения разных компонентов биоты позволит соотнести особенности их биологии, экологии и таксономической эволюции со спецификой флорогенеза и биотогенеза. Все это даст возможность понять глубинные закономерности формирования биомов Земли, механизмов их устойчивости и сохранения целостности [75]. Развитие основ сопряженного биогеографического анализа будет способствовать решению проблемы комплексного биогеографического районирования [71]. Результаты сопряженного анализа полезны при организации многоуровневого мониторинга и разработке стратегии сохранения биоразнообразия [4]. Однако неполнота данных по бриофлорам, альгофлорам, лихенобиотам, микобиотам многих территорий пока не позволяет проводить сопряженный анализ по этим и другим компонентам биоразнообразия, которые традиционно являются предметом изучения флористов и биогеографов. Актуальны комплексные исследования больших по площади территорий, составление каталогов и обзоров по разным компонентам биоты [11; 42], разработка подходов, позволяющих оценивать степень полноты выявления видового состава [2 – 4; 67].
Необходим поиск модельных территорий большого масштаба, на базе которых возможны корректный анализ разных компонентов, выявление основных широтных и долготных трендов изменения параметров и характеристик флор и биот. Более равномерно и полно исследованы флоры мхов [9; 79], менее полно изучены флоры печеночников и биоты лишайников [41; 53]. Особый интерес представляют модельные территории, имеющие естественные физико-географические и ботанико-географические границы. При анализе их флор и биот могут быть выявлены важнейшие градиенты и тенденции изменения параметров разнообразия, которые впоследствии будут детализированы при работе с территориями меньших размеров, моделирующими в общих чертах крупные выделы [66; 67]. Методика «ретроспективной» оценки пространственного распределения разных компонентов биоразнообразия нуждается в дальнейшей разработке. При сопряженном изучении флор и биот целесообразно расширение спектра анализируемых факторов. В большей степени в литературе рассмотрены тенденции изменения показателей флор сосудистых растений в широтном и долготном направлениях [18; 19; 48; 66]. В меньшей степени изучен характер зависимости основных характеристик флор от геоморфологических и ландшафтных особенностей территории [32; 33]. Не получает полного отражения при таксономическом и генетическом анализе флор региональная специфика таксономической дифференциации и расселения многих систематических групп. Есть основания предполагать, что видовой состав флор мхов в большей степени, чем флор сосудистых растений, зависит от литологических особенностей и геоморфологии территории, в их дифференциальных компонентах существенную роль играют эпилитные и эпигейные виды, приуроченные к обнажениям различных субстратов [32]. Определенные трудности при выделении широтных географических элементов у мхов, вероятно, также обусловлены относительно меньшим влиянием на характер распространения многих видов физико-географических факторов, отражающих специфику широтного положения территории. Отмеченное выше справедливо и для лишайников [43; 44; 50 – 52]. В этой связи при разработке основ сопряженного анализа разных компонентов биоразнообразия могут представлять интерес системы физикогеографического районирования.
Традиционно используют разные индексы и показатели сходства видового состава [65; 66]. При сравнении территорий разной площади удобен индекс Отиаи [20; 21; 32; 83].
Целесообразен дальнейший поиск параметров, характеризующих специфику флорогенеза и особенности региональной дифференциации ведущих таксонов. Во флористических работах уже использовали такие признаки как отношение числа видов в семействах Cyperaceae–Poaceae, Asteraceae–Poaceae, Asteraceae–Fabaceae [66; 73; 74], Pottiaceae–Dicranaceae, число видов в роде Cinclidotus P.Beauv. [78]. Заслуживает специального внимания анализ разнообразия таксонов с широким распространением апомиксиса. Число микровидов в родах Hieracium L., Alchemilla L., Rubus L. для разных базовых флористических выделах сильно отличается [21; 32; 82]. Состав микровидов может существенно дополнить данные о специфике флор, поэтому агамные комплексы на разных территориях также требуют дальнейшего изучения. Необходима критическая ревизия всего материала.
На данном этапе не всегда удается соотнести всю флористическую и биогеографическую информацию с системой геоботанического и физикогеографического районирования, так как многие сводки и обзоры построены по административно-территориальному принципу. В пределах каждой административной области встречаются фрагменты разных провинций, что с одной стороны осложняет систематизацию данных по физикогеографическому принципу, а с другой стороны определяет некорректность сравнительного анализа флор административных областей. Многие области включают крупные фрагменты двух-трех, а некоторые четырех-пяти провинций. Иногда территорию области пересекают разные физикогеографические страны. Например, часть территории Ленинградской обл. входит в состав Фенноскандии, а центральные и южные районы области расположены в пределах Русской равнины.
У сосудистых растений иногда сложно однозначно определить флорогенетический статус вида и отнести его к группе аборигенных или адвентивных. Требует дальнейшего обсуждения проблема выбора системы районирования, которая в большей степени удовлетворяет задаче сопряженного анализа. Характер географического распределения отдельных групп нередко зависит в большей степени от особенностей геоморфологии и литологии территории, чем от структуры растительного покрова. В этой связи больше возможностей представляет физико-географическое, а не геоботаническое районирование. В первом случае оказываются учтенными не только особенности широтной зональности растительности, долготной климатической и ландшафтной дифференциации, но и азональные геологические и связанные с ними геоморфологические особенности территории. Хотя физико-географические провинции имеют разную площадь [55; 56], их размеры меньше, чем размеры фитохорий и ботаникогеографических районов [45; 54]. Провинции более сопоставимы по площади, едины с точки зрения генезиса и ландшафтной структуры.
Для разработки основ сопряженного флористического анализа разных компонентов биоразнообразия необходим поиск модельных территорий большого масштаба. Важным условием является высокая степень флористической и биогеографической изученности разных компонентов биоты, которая сделает возможной корректную оценку относительной роли факторов, определяющих специфические характеристики флор и биот. Удобной модельной территорией для сопряженного анализа флор мхов и сосудистых растений, других компонентов биоты может быть Русская равнина [32; 33]. Она неоднородна в геоморфологическом отношении [7], есть районы, расположенные на стыке природных зон, разнообразны местообитания с компонентами экстразональной растительности. На территории Русской равнины проходили активные миграции разных флороценотических комплексов. Большая протяженность и достаточное разнообразие в физикогеографическом отношении позволяют выделять группы модельных районов, на базе которых можно выяснять роль разных физико-географических факторов, оценивать региональную специфику таксономической дифференциации систематических групп. В пределах Русской равнины достаточно полно выявлен видовой состав флор сосудистых растений и мхов, четко прослеживаются зональные и долготные тренды изменения параметров флор [32]. Продолжается сбор материалов по лишайникам и миксомицетам в разных областях Центральной и Северо-Западной России [5; 10; 15; 24; 25; 34; 35 и др.].
Нами предпринята попытка анализа флор сосудистых растений и листостебельных мхов Валдайской и смежных физико-географических провинций [32; 33]. Начато обобщение материалов по лишайникам и миксомицетам [15; 31; 35]. Выявлен видовой состав флор сосудистых растений и мхов шести физико-географических провинций. Среди них ЗападноПриморская (ПРП), Прибалтийская (ПП), Валдайская (ВП), Смоленско-Московская (СМП), Мещерская (МП) и Верхневолжская (ВВП). В целом модельный регион включает полностью Тверскую, Новгородскую, Ярославскую области, значительную часть Смоленской, Псковской областей, большие фрагменты Ленинградской, Московской, Костромской областей,
Эстонии, Латвии, Литвы, некоторые районы Вологодской, Ивановской и Калужской областей (рис. 1, 2). В общей сложности анализируемая территория характеризуется значительной протяженностью в широтном и долготном направлениях. Все провинции расположены в пределах лесной зоны [7]. Они отличаются по степени континентальности климата структуре растительного покрова, геоморфологическому строению [7]. Бóльшая их часть расположена в подзоне южной тайги. Зональными типами растительности являются южнотаежные и широколиственно-еловые (подтаежные) леса. К ее южной границе примыкает зона широколиственных лесов. В виде небольших фрагментов экстразональной растительности в долинах крупных рек встречаются луговостепные сообщества. Лишь на севере Верхневолжской провинции встречаются средне-таежные леса. Значительную часть Смоленско-Московской провинции занимают широколиственно-еловые (подтаежные) леса, на юге встречаются широколиственные лесные формации. Модельная территория неоднородна в геоморфологическом отношении. В ВП и СМП широко распространены местообитания с большим разнообразием микрониш, на которых формируются эпилитные синузии. ПРП, МП, ПП и ВВП – преимущественно низменные равнинные территории с небольшими по площади отдельными возвышениями и грядами. ВП и СМП – возвышенные территории с холмистым рельефом. В пределах Валдайской возвышенности четко просматривается влияние последнего Валдайского оледенения. Преобладает расчлененный холмисто-грядовый рельеф, наблюдается обилие ледниковых озер самых разнообразных форм и размеров. По всей территории провинции часто встречаются россыпи валунов. В СМП долинные ландшафты с обнажениями карбонатных пород образуют сложные большие по площади комплексы.
Взятые за основу источники информации по бриофлорам и флорам сосудистых растений процитированы в более ранних публикациях, в которых отражены некоторые результаты предварительного анализа [32; 33]. Из флористических списков сосудистых растений исключены адвентивные виды. Дополнительно учтены данные о микровидах и некоторых критических таксонах, видовую самостоятельность которых признают не все исследователи [32]. Выявлен состав специфических компонентов флор мхов и сосудистых растений, биот лишайников и миксомицетов. Специфические компоненты включают дифференциальные (встречающиеся только в пределах одной провинции) и кодифференциальные (отмечены только в двух или трех провинциях) виды. Выяснены уровни видового богатства и объемы специфических компонентов флор сосудистых растений и мхов (рис. 1). Оценена степень сходства видового состава их флор с использованием индекса Отиаи [32]. Составлены предварительные списки лишайников и миксомицетов [см. 15; 35], выявлены уровни видового богатства и специфичности их биот (рис. 2, табл. 3).
Флоры мхов и сосудистых растений
Уровень видового богатства флор
На анализируемой территории в общей сложности отмечено 509 видов мхов и 1677 видов сосудистых растений (с учетом критических таксонов и микровидов – 2801 вид). Уровень видового богатства во флорах мхов варьирует от 222 до 447 видов, во флорах сосудистых растений – от 857 до 1203 (табл. 1). В конкретных физико-географических провинциях представлено от 43,6 до 87,8% общего списка мхов и от 51,1 до 71,7% сосудистых растений (табл. 1, рис. 1).
У мхов и сосудистых растений выявлена тенденция уменьшения видового богатства флор в направлении с запада на восток. Наибольший уровень видового богатства отмечен в приатлантических районах (табл. 1, рис. 1). Установлены значительные различия в скорости изменения показателей флористического богатства у мхов и сосудистых растений. Анализируя относительный уровень разнообразия флор мхов (видовое богатство флоры провинции в процентах от общего числа видов, отмеченных на территории, включающей все рассматриваемые провинции) получаем следующие ряды характеристик – ПРП – 87,8%, ПП – 85,7, ВП – 47,0, ВВП – 43,6. Для сосудистых растений показатели другие – ПРП – 63,4%, ПП – 71,7, ВП – 51,1, ВВП – 51,7 (табл. 1). Уменьшение уровня видового богатства в направлении с запада на восток более четко проявляется во флорах мхов. Различия количественных характеристик для них более контрастны (рис. 1). Приатлантические флоры мхов более чем в 2 раза богаче по числу видов, по сравнению с флорой ВВП. В соответствующих флорах сосудистых растений аналогичные характеристики отличаются всего на 20%. При этом пропорциональность изменения показателей нарушена, по-видимому, в связи с разной площадью территорий ПП и ВП, а также в результате возможного пограничного положения ПП (на стыке флористических провинций).
Во флорах сосудистых растений в направлении с севера на юг четко выявляется тенденция увеличения уровня видового богатства (ВП – 857 видов, СМП – 951, ВВП – 867, МП – 1023) (табл. 1, рис. 1). При этом приросты числа видов для ВП – СМП и ВВП – МП пропорциональны (в первом случае общий уровень видового богатства увеличивается на 11%, во втором – на 18%). В бриофлорах отмечена сходная тенденция, но различия количественных характеристик менее пропорциональны (ВП – 239, СМП – 291, ВВП – 222, МП – 241, и соответственно на 21% и 8,5%).
Степень специфичности флор
У мхов и сосудистых растений выявлена тенденция уменьшения объема специфического компонента флор с запада на восток (рис. 3, Б, табл. 1, 2). У мхов она проявляется более четко и контрастно. Во флорах сосудистых растений наблюдается значительное увеличение объема специфического компонента в направлении с севера на юг (ВП – 40 видов, СМП – 109, ВВП – 42, МП – 180) (табл. 1, рис. 1). Во флорах мхов по этим показателям отмечена макромозаичность (ВП – 28, СМП – 42, ВВП – 21, МП – 12) (рис. 3, В).
Уменьшение удельного веса дифференциального компонента в направлении с запада на восток выявлено как во флорах мхов, так и во флорах сосудистых растений (рис. 3, Б). Изменение объема дифференциального компонента в направлении с севера на юг у сосудистых растений и мхов имеет разнонаправленный характер (рис. 3, Б). Выявлены различия в соотношении удельного веса дифференциальных и кодифференциальных видов в составе специфического компонента флор мхов и сосудистых растений. Во флорах сосудистых растений доля дифференциальных видов варьирует от 0 до 10,7 %, а кодифференциальных от 0 до 20,9 %. Во флорах мхов эти показатели изменяются в интервале от 0,4 до 9,6% и от 0 до 37,4% соответственно. Особенно четко эти различия прослеживаются в приатлантических флорах (в ПРП дифференциальные виды мхов составляют 22,1% от общего числа специфических видов, а дифференциальные виды сосудистых растений – 40,9% специфических видов, в ПП – 14,2% и 19,7% соответственно). Наиболее контрастны показатели для МП (8,3% и 52,8%). У мхов на анализируемой территории отсутствуют эндемичные виды (табл. 2). Относительно меньший объем дифференциального компонента и отсутствие эндемиков свидетельствуют в пользу отмеченной ранее закономерности – большинство мохообразных имеют более широкие ареалы, чем сосудистые растения [77; 78].
Анализ географических спектров флор мхов и сосудистых растений показал, что большее дифференцирующее значение имеют средне-атлантическо-европейская и степная-лесостепная (аридная) фракции (рис. 3). Во флорах сосудистых растений доля степных и лесостепных видов меняется следующим образом: ВП – 1,2%, СМП – 7,6%, ВВП – 2,9%, МП – 12,8% (рис. 3). Во флорах мхов также наблюдается изменение доли аридных видов, но различия менее контрастны (ВП – 0,4%, СМП – 2,1%, ВВП – 0,5%, МП – 1,7%). Скорость изменения доли степной, лесостепной и аридной фракций у сосудистых растений больше, чем у мхов (рис. 3). Показатели удельного веса аридных видов во флорах мхов более диспропорциональны (рис. 3). Как во флорах мхов, так и во флорах сосудистых растений прослеживается тенденция уменьшения доли среднеевропейских и атлантическо-европейских видов с запада на восток. Во флорах мхов удельный вес этой фракции изменяется более плавно. У сосудистых растений различия более контрастны и диспропорциональны (рис. 3). Изменение долевого участия дифференцирующих фракций во флорах мхов и сосудистых растений происходит по-разному (рис. 3, Г). Во флорах мхов специфика географической структуры в большей степени определяется атлантическо-европейской фракцией, во флорах сосудистых растений – лесостепной-степной. Различия в структуре географических спектров флор сосудистых растений и мхов могут быть еще более четкими при анализе отдельных физико-географических районов, расположенных в пределах разных провинций [30].
Установленные тенденции изменения географической структуры флор проявляются более контрастно при качественном анализе специфических компонентов (рис. 3). Во флорах мхов доля специфических видов с атлантическими связями меняется более пропорционально, чем у сосудистых растений (рис. 3). Во флоре сосудистых растений ВВП практически не остается специфических видов с атлантическими связями, но появляются восточноевропейско-сибирские виды. Среди дифференциальных видов ВВП есть некоторые евросибирские растения (Cacalia hastatа L., Calamagrostis obtusata Trin., Schizahne callosa (Turcz. ex Griseb.) Ohwi, Anemonoides altaica (C.A. Mey.) Holub). Градиент изменения доли аридных видов во флорах мхов (ВП – 0 дифференциальных и 0 кодифференциальных видов, СМП – 1 и 4, ВВП – 0 и 0, МП – 1 и 3 соответственно) менее контрастен, чем градиент увеличения доли степных и лесостепных видов у сосудистых растений (ВП – 0 и 2, СМП – 4 и 44, ВВП – 0 и 7, МП – 65 и 41) (табл. 2).
Показатели видового богатства и уровня специфичности состава флор сосудистых растений существенно изменяются при учете разнообразия микровидов и критических таксонов (табл. 1, 2, рис. 3). Уменьшается относительный уровень репрезентативности флор в связи с увеличением общего уровня флористического богатства территории и более локальным распространением специфических микровидов. Существенно возрастает доля дифференциальных видов и объем специфического компонента в приатлантических флорах. Относительные значения этих показателей даже превосходят соответствующие количественные характеристики флор мхов (рис. 3, Б). В некоторых случаях, например, на отрезке ВП – СМП, градиенты, характеризующие удельный вес специфических компонентов, приобретают противоположную направленность. Тенденция увеличения объема специфических видов в приатлантических районах выявлена почти для всех агамных комплексов. Особенно четко она прослеживается в родах Hieracium , Taraxacum Wigg., Ranunculus L. [32]. Характер географического распределения разнообразия агамных микровидов в роде Alchemilla иной. В этом роде проявляется тенденция увеличения числа дифференциальных видов и объема группы в направлении с запада на восток [32]. В Прибалтике, как и в Скандинавии, дифференциальных видов рода Alchemilla немного, а большинство широко распространенных в Европе агамных видов, по-видимому, являются натурализовавшимися заносными растениями, которые в настоящее время их можно отнести к группе археофитов. Географическая оригинальность флор приатлантических районов существенно возрастает благодаря включению агамных микровидов (рис. 3, Г). Изменяются пропорции долевого участия средне-атлантическо-европейской и лесостепной-степной фракций. В приатлантических районах увеличивается доля первой фракции, а в более континентальных районах уменьшается доля второй. В целом учет мировидов увеличивает контрастность долготных трендов на отрезке, примыкающем к атлантическим районам.
Уровень сходства видового состава
Показатели, характеризующие степень сходства видового состава флор (индекс Отиаи), у мхов и сосудистых растений разные [32]. Состав флор сосудистых растений без учета разнообразия микровидов сходен. Более близки флоры СМП, ВВП и ВП. Максимальные различия выявлены между МП и ПРП (индекс Отиаи 0,67).
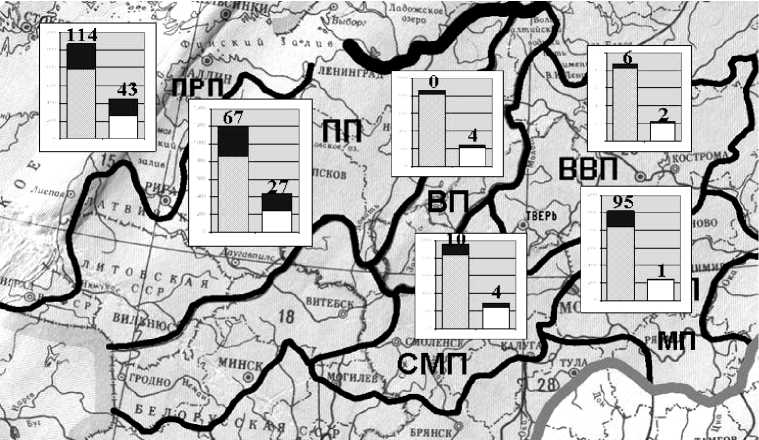
Рис. 1. Видовое богатство, специфические и дифференциальные компоненты флор сосудистых растений (левые столбцы)
и мхов (правые столбцы) Валдайской и смежных провинций: заливка – специфический компонент; цифры – дифференциальные виды; пунктир – административные единицы; черная жирная линия – физико-географическе страны; черная линия средней толщины – провинции; светлая жирная – природные зоны
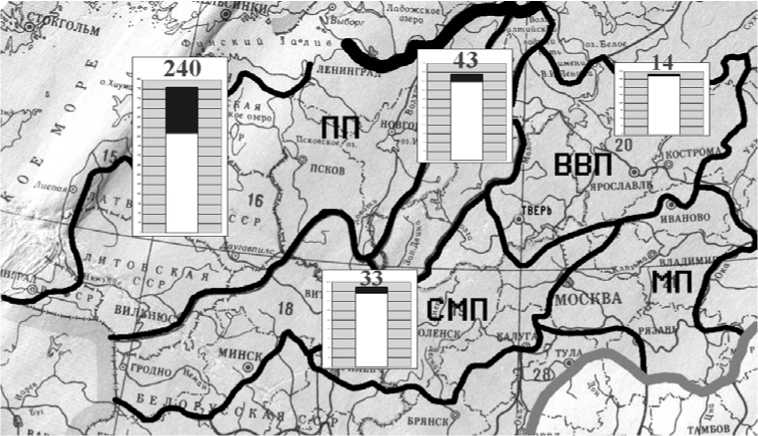
Рис. 2. Видовое богатство и число дифференциальных видов лихенобиот Валдайской и смежных провинций: заливка – дифференциальный компонент; прочие обозначения как на рис. 1
Таблица 1
Уровень видового богатства и объем специфического компонента флор сосудистых растений и мхов Валдайской и смежных провинций
|
Флора |
Сосудистые растения |
Мхи |
||||
|
общее число видов |
диф. виды |
спец. компонент |
общее число видов |
диф. виды |
спец. компонент |
|
|
ПРП |
1064 (1478) 63,4 (52,8) |
114 (241) |
279 (646) |
447 87,8 |
43 |
195 |
|
ПП |
1203 (1641) 71,7 (58,6) |
67 (161) |
340 (718) |
436 85,7 |
27 |
190 |
|
ВП |
857 (1037) 51,1 (37,0) |
0 (2) |
40 (130) |
239 47,0 |
4 |
28 |
|
СМП |
951 (1080) 57,0 (38,6) |
10 (18) |
109 (136) |
291 57,2 |
4 |
42 |
|
ВВП |
867 (972) 51,7 (34,7) |
6 (11) |
42 (66) |
222 43,6 |
2 |
21 |
|
МП |
1023 (1107) 61,0 (39,5) |
95 (108) |
180 (206) |
241 47,3 |
1 |
12 |
Примечание. диф. – дифференциальные, спец. – специфический; ПРП – ЗападноПриморская; ПП – Прибалтийская; ВП – Валдайская; СМП – Смоленско-Московская; ВВП – Верхневолжская; МП – Мещерская; цифровые данные с учетом состава агамных комплексов и критических таксонов даны в скобках. Под чертой указана доля от общего числа видов, отмеченных на всей анализируемой территории (в %).
Таблица 2
Характеристика специфического компонента флор сосудистых растений и мхов Валдайской и смежных провинций
|
Флора |
Общее число видов |
Е |
СЕ |
Э |
С, А |
|
Сосудистые растения |
|||||
|
ПРП |
279(646) |
122(380) |
11(330) |
4(70) |
9(11) |
|
ПП |
340(718) |
130(341) |
92(285) |
5(38) |
57(58) |
|
ВП |
40(130) |
4(20) |
3(15) |
0(4) |
2(2) |
|
СМП |
109(136) |
18(32) |
7(12) |
2(7) |
48(48) |
|
ВВП |
42(66) |
7(17) |
2(8) |
1(4) |
7(7) |
|
МП |
180(206) |
32(44) |
4(10) |
6(15) |
106(106) |
|
Мхи |
|||||
|
ПРП |
195 |
58 |
49 |
0 |
13 |
|
ПП |
190 |
49 |
45 |
0 |
12 |
|
ВП |
28 |
5 |
7 |
0 |
0 |
|
СМП |
42 |
8 |
3 |
0 |
6 |
|
ВВП |
21 |
3 |
2 |
0 |
0 |
|
МП |
12 |
1 |
0 |
0 |
4 |
Примечание. Е – европейские и преимущественно европейские; СЕ – атлантическо-европейские и преимущественно среднеевропейские; Э – эндемики; С, А – степные и лесостепные или аридные.
Флоры мхов четко распадаются на две плеяды [32]. Одна из них объединяет приатлантические флоры (ПРП и ПП), другая – континентальные. Связи между плеядами слабые. По сравнению с флорами сосудистых растений связи внутри плеяд слабее. Максимальное сходство отмечено между ПРП и ПП (0,88). Минимальное сходство обнаруживают флоры ПРП и ВВП. При учете разнообразия микровидов флоры сосудистых растений также группируются в две корреляционные плеяды. Максимальное сходство обнаруживают СМП и ВВП (0,86), а минимальное – МП и ПРП (0,60).
Сопоставление характера связей флор мхов и сосудистых растений геоморфологически сходных провинций (ВП, СМП и ВВП) обнаруживает определенные различия [32]. У мхов более сходны между собой флоры ВП и СМП. Значение индекса (0,81) приближается к максимальному для флор мхов уровню. При этом флоры провинций, имеющих максимальную по протяженности общую границу (ВП и ВВП) характеризуются меньшим сходством (0,76), чем СМП и ВП. У сосудистых растений сходство между флорами ВП и ВВП достигает максимального значения (0,92), а флоры СМП и ВП менее сходны.
Специфические особенности флор мхов и сосудистых растений
По основным количественным характеристикам (флористическое богатство, относительный уровень видового богатства флор, доля дифференциальных видов, удельный вес специфического компонента, пропорции долевого участия дифференцирующих фракций географических элементов) в пределах анализируемой территории долготные тренды во флорах мхов проявляются более четко и контрастно, чем во флорах сосудистых растений. Изменения параметров более пропорциональны. Широтные тренды во флорах мхов по сравнению с флорами сосудистых растений выражены менее четко. Градиенты менее контрастны. Некоторые из них имеют противоположную по сравнению с флорами сосудистых растений направленность (например, доля дифференциальных видов во флорах) или не выявляются в связи с макромозаичностью (объем специфического компонента). При сопоставлении структуры географических спектров флор мхов и сосудистых растений отмечено разное соотношение удельного веса фракций, имеющих дифференцирующее значение. Специфика географической структуры флор мхов в большей степени определяется атлантическо-европейской фракцией, а специфика флор сосудистых растений – степной и лесостепной фракциями (рис. 3, Г). Выявленные различия обусловлены разным характером влияния широтных и долготных физико-географических градиентов на пространственное распределение разнообразия мхов и сосудистых растений. Географическое распространение мхов в отличие от сосудистых растений в большей степени зависит от характеристик водного режима территории и в меньшей степени от температурного фактора. Учет разнообразия микровидов сосудистых растений усиливает долготные флористические тренды, а в некоторых случаях «затушевывает» проявления широтных тенденций (например, уменьшение объема специфического компонента на отрезке ВП – СМП) (рис. 3, В).
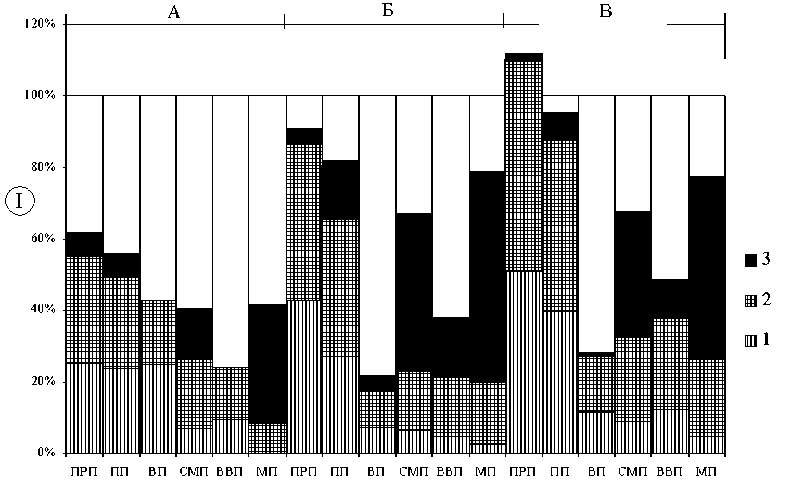
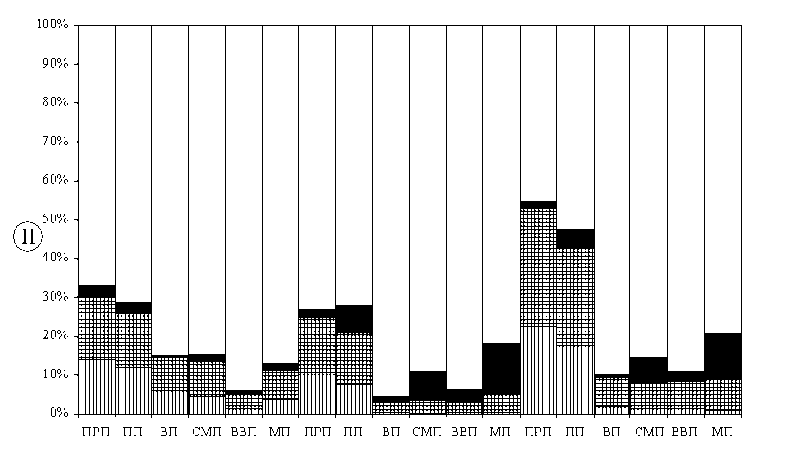
Рис. 3. Доля некоторых фракций географических элементов во флорах (II) и в составе специфического компонента флор (I):
А – мхи; Б – сосудистые растения; В – сосудистые растения с учетом микровидов; 1 – преимущественно среднеевропейские и атлантическо-европейские виды; 2 – европейские виды; 3 – степные и лесостепные или аридные виды; прочие обозначения такие же, как в табл. 1.
Проявление макромозаичности в пределах направленных рядов изменения количественных харакетристик флор мхов и сосудистых растений
Вестник ТвГУ. Серия «Биология и экология». Вып. 14, 2009 обусловлены разными факторами. Во флорах сосудистых растений «маскировка» направленных изменений некоторых параметров
(относительный уровень видового богатства флор, доля дифференциальных видов, относительный объем специфического компонента) на отрезках ПРП – ПП, ВП – ВВП (рис. 1, табл. 1) может быть связана с разными размерами провинций, а в случае ПП – возможным прохождением по территории провинции границ фитохорий.
Невысокий уровень видового богатства флоры ВП и низкая ее специфичность могут быть также следствием более ограниченного миграционного потока и степных и лесостепных видов. Противоположная направленность тренда у сосудистых растений при учете разнообразия микровидов на отрезке ВП – СМП обусловлена спецификой таксономической дифференциации и расселения крупных агамных комплексов. В пределах ВП распространен целый ряд скандинавских и атлантическо-европейских микровидов из родов Hieracium, Taraxacum, Ranunculus , которые уже не встречаются в СМП. Случаи макромозаичности характеристик флор мхов (противоположная направленность градиентов изменения доли специфического компонента на отрезке ВП – СМП (рис. 3, В)) связаны с геоморфологической спецификой территории. В СМП широко распространены ландшафты с разнообразными микронишами, занимаемыми кальцефильными эпилитами. Эти же факторы определяют и различия уровней сходства видового состава флор мхов и сосудистых растений ВП, СМП и ВВП. Территории всех изученных провинций имеют достаточно большие площади (в среднем около 98,3 кв. км) и характеризуются неоднородностью ландшафтной и геоморфологической структур. При анализе параметров конкретных флор мхов, которые приурочены к более однородным в геоморфологическом и ландшафтном отношениях территориям, макромозаичность может проявляться еще полнее и четче [30].
Таким образом, в пределах Валдайской и смежных физикогеографических провинций, зональные тенденции изменения общего уровня видового богатства и степени специфичности флор более четко проявляются у сосудистых растений, чем у мхов. Специфика бриофлор в большей степени определяется долготным положением территории. Степень оригинальности флор сосудистых растений в долготном направлении увеличивается при учете состава агамных комплексов. На основные характеристики флор мхов оказывают определенное влияние геоморфологические особенности территории.
Биоты лишайников
Детальный сравнительный анализ биот лишайников пока не возможен в связи с разной степенью изученности многих регионов России [35; 53]. На основе анализа данных литературы (252 источника, которые процитированы в предварительном сообщении [35]) и гербарных материалов составлены предварительные списки лишайников ПП, ВП, ВВП, СМП, ПРП. Сделан обзор по областям Центральной России, расположенным на территории этих провинций [35]. Списки для провинций существенно дополнены с учетом новых публикаций [6; 24; 25; 29; 34; 46; 81 и др.]. В результате проведенных в последнее время исследований в Ленинградской, Ярославской, Тверской и Московской областях степень изученности этих регионов стала в настоящее время относительно сопоставимой.
На территории, включающей все отмеченные провинции к настоящему времени зарегистрировано не менее 952 видов лишайников и лихенофильных грибов. Оценить уровень видового богатства биот каждой провинции точно пока невозможно в связи с тем, что данные по лишайникам стран Балтии не всегда удавалось соотнести с физико-географическим районированием. В этой связи сделан предварительный анализ биот только четырех провинций (ПП, ВП, ВВП, СМП). В ПП зарегистрирован 761 вид, в СМП – 425, в ВП – 490, в ВВП – 387 (табл. 3). Разный уровень видового богатства обусловлен проявлением тенденции уменьшения разнообразия лихенобиот вдоль градиента увеличения степени континентальности климата (рис. 2). Характер изменения показателей сходен с мохообразными. Дальнейшие лихенологические исследования позволят дать более точную характеристику этого тренда, с учетом территории Западно-Приморской провинции. В пределах одного широтного интервала большим уровнем видового богатства характеризуются провинции, на территории которых есть крупные возвышенности и более широко распространены ландшафты с обнажениями коренных материнских пород и россыпями валунов (ВП и СМП). Структура таксономических спектров биот отдельных провинций и территории в целом сходна и отражает характерные особенности флор бореальной зоны [35].
Во всех провинциях встречаются 260 видов. Уровень специфичности видового состава на данном этапе изучения лихенобиот можно охарактеризовать следующим образом. Только в ПП зарегистрировано 240 видов, в ВП – 43, в СМП – 33, в ВВП – 14 (табл. 3, рис. 2). Для получения более точных данных об уровне специфичности лихенобиот необходимо специальное изучение многих групп накипных лишайников. Микролишайники и лихенофильные грибы еще недостаточно изучены во многих областях Центральной России. В хорошо изученных биотах микролишайники примерно в два раза преобладают по численности над макролишайниками [53]. Общие списки модельных территорий может существенно дополнить более детальный анализ материала по представителям родов Arthopyrenia A. Massal., Buellia De Not., Calicium Pers., Caloplaca Th. Fr., Chaenothecopsis Vain., Lepraria Ach., Micarea Fr., Pertusaria DC., Rinodina (Ach.) S. Gray, Usnea Dill. ex Adans., Verrucaria Schrad. Списки лишайников отдельных областей могут быть дополнены после изучения разнообразия родов Arthonia Ach., Bacidia De Not., Bryoria Brodo et D. Hawksw., Collema F. H. Wigg., Lecania A. Massal., Lecanora Ach., Opegrapha Ach. Пока только в одной или немногих областях зарегистрированы представители семейств Arthopyreniaceae W. R. Watson, Catillariaceae Hafellner, Mycoblastaceae Hafellner, Pannariaceae Tuck., Pilocarpaceae Zahlbr., Pyrenulaceae Rabench., Thelenellaceae H. Mayrhofer, Thrombiaceae Poelt ex. J. C. David et D. Hawksw., Trichothelliaceae (Müll. Arg.) Bitter et F. Schill.
Среди дифференциальных и специфических видов есть лишайники, представляющие разные географические элементы. Зональная приуроченность в распространении лишайников также как и у мхов выявляется не четко. Большинство отмеченных арктоальпийских видов по территории Валдайской возвышенности и приморским районам заходят достаточно далеко на юг. Лишь немногие арктоальпийские виды отмечены только в самых северных районах территории ( Arctoparmelia centrifuga (L.) Hale, Flavocetraria nivalis (L.) Kärnefeld et Thell). Среди аридных видов только на южной границе модельной территории зарегистрирована Caloplaca lactea (A. Massal.) Zahlbr.
Среди специфических видов ПП четко выявляются лишайники с океаническим и субокеаническим характером распространения – Dimerella lutea (Dicks.) Trevis., Hypotrachyna revoluta (Flörke) Hale, Lecidella laureri (Hepp) Körb., Leptogium palmatum (Huds.) Mont., Maronea constans (Nyl.) Hepp, Peltigera membranacea (Ach.) Nyl., Pertusaria flavida (DC.) J.R. Laundon, Punctelia subrudecta (Nyl.) Krog, Pyrenula nitidella (Flörke ex Schaer.) Müll. Arg. [35]. Долготные градиенты проявляются также в разной активности и частоте встречаемости некоторых субокеанических видов на анализируемой территории. Среди них Acrocordia cavata (Ach.) R. C. Harris, Anaptychia rucinata (With.) J.R. Laundon, Cetrelia olivetorum (Nyl.) W.L. Culb. et C.F. Culb., Collema nigrescens (Huds.) DC., Heterodermia speciosa (Wulfen) Trevis., Lobaria pulmonaria (L.) Hoffm., Menegazzia terebrata (Hoffm.) A. Massal., Pertusaria hemisphaerica (Florke) Erichsen, Phaeographis dendricta (Ach.) Mull. Arg., Pyrenula nitida (Weigel) Ach., Sphinctrina anglica Nyl., Thelotrema lepadinum (Ach.) Ach. Они обычны в приокеанических районах, но редки в субконтинентальных и континентальных [35].
Таблица 3
Уровень видового богатства и число дифференциальных видов биот лишайников и миксомицетов Валдайской и смежных провинций
|
Биота |
Лишайники |
Миксомицеты |
||
|
всего видов |
дифференциальные виды |
всего видов |
дифференциальные виды |
|
|
ПП |
761 86,6 |
240 |
154 75,5 |
32 |
|
ВП |
490 55,8 |
43 |
63 30,9 |
2 |
|
СМП |
425 48,4 |
33 |
150 73,5 |
28 |
|
ВВП |
387 44,1 |
14 |
109 53,5 |
6 |
Примечание . Под чертой указана доля от общего числа видов, отмеченных на всей анализируемой территории (в %).
Значительное влияние на характер распространения видов и уровень видового богатства флор оказывают геоморфологические особенности. Распространение ряда лишайников четко связано с территорией Валдайской возвышенности. Относительный уровень видового богатства и специфичности лихенобиоты этой территории выше, чем аналогичные показатели для флор мхов и сосудистых растений. На территории Валдайской возвышенности у лишайников более полно, чем у мхов представлены арктоальпийские и монтанные виды. Четко проявляются связи с Фенноскандией благодаря широкому распространению валунов и глубоких оврагов с ручьями и каменистым дном. В Валдайской провинции отмечены разные арктоальпийские (Bellemerea cupreoatra (Nyl.) Clauzade, Cetraria odontella (Ach.) Ach., Lecidea plana (J. Lahm.) Nyl., Porpidia speirea (Ach.) Kremp., Protoparmelia badia (Hoffm.) Hafellner, Stereocaulon alpinum Laurer) и монтанные виды (Acarospora sinopica (Wahlenb. in Ach.) Korb., Aspicilia gibbosa (Ach.) Korb., A. laevata (Ach.) Arnold, Dermatocarpon luridum (With.) J.R. Laundon, D. rivulorum (Arnold) Dalla Torre et Sarnth., Lecanora cenisia Ach., Lecidea lithophila (Ach.) Ach., Melanelia sorediata (Ach.) Goward et Ahti, Parmelia fraudans (Nyl.) Nyl., Rhizocarpon disporum (Nageli ex Hepp.) Müll. Arg., R. hochstetteri (Korb.) Vain., Stereocaulon saxatile H. Magn., Thelidium minutulum Korb., T. zwackhii (Hepp) A. Massal., Umbilicaria hyperborea (Ach.) Hoffm., U. polyphylla (L.) Baumg, U. torrefacta (Lightf.) Schrad.) [35].
Особенно четко специфика биоты, обусловленная геоморфологическими особенностями, выявляется на более ограниченных по площади территориях. Например, в пределах Тверской обл. в пределах Валдайской возвышенности наблюдается массовое распространение многих эпилитных видов. Среди них Acarospora fuscata (Schrad.) Th. Fr., Aspicilia cinerea (L.) Körb., Lecanora cenisia Ach., Protoparmeliopsis muralis (Schreb.) M. Choisy, Rhizocarpon eupetraeum (Nyl.) Arnold, Xanthoparmelia conspersa (Ach.) Hale. Регулярно встречается в пределах Валдайской возвышенности Melanelia sorediata [35]. В настоящее время вид зарегистрирован во всех физикогеографических районах тверского участка Валдайской возвышенности, отмечен на крайнем юго-западе области. В Ленинградской обл. большая часть местонахождений вида сосредоточена на территории относящейся к Фенноскандии. Единичные местонахождения за её пределами приурочены к Вепсовской и Тихвинской возвышенностям (одна точка в Ленинградской и три точки в Новгородской обл.) [35].
Биоты миксомицетов
Биоты миксомицетов анализируемых физико-географических провинций характеризуются разной степенью изученности [15; 28]. Меньше всего материалов по ВП [16]. По-видимому, еще не до конца выявлен видовой состав ВВП. В этой связи, данные об уровнях видового богатства и степени специфичности биот (табл. 3) носят предварительный характер. Выявляемые различия в большей степени связаны с неравномерной изученностью территории. Наиболее полно исследованные провинции (СМП и ПП) мало отличаются по уровню видового богатства (табл. 3).
На современном этапе изучения миксомицетов данные о дифференциальных и кодифференциальных видах свидетельствуют о более высоком уровне своеобразия биоты СМП (табл. 4). В ВП обнаружено только два дифференциальных вида (Physarum serpula, Symphytocarpus impexus). Каждый из них отмечен только в единственном местонахождении – болотный массив в Еремковском лесничестве Удомельского р-на [16]. Принадлежность этих видов к определенному географическому элементу не выявлена [28]. Возможно Symphytocarpus impexus и другие виды этого рода могут быть найдены при более детальном исследовании крупных болотных массивов в ВВП и других провинциях.
Таблица 4 Некоторые дифференциальные и кодифференциальные виды миксомицетов
Валдайской и смежных физико-географических провинций
[по: 15, с изменениями]
|
Вид |
ГЭ |
Са+ |
ВП |
ВВП |
СМП |
|
Physarum serpula Morgan |
? |
+ |
+ |
||
|
Symphytocarpus impexus Ing et Nann.-Brem. |
? |
+ |
|||
|
Didymium iridis (Ditmar) Fr. |
нб |
+ |
+ |
||
|
Licea biforis Morgan |
бн |
+ |
|||
|
Lycogala conicum Pers. |
н |
+ |
|||
|
Physarum citrinum Schum. |
? |
+ |
+ |
||
|
P. famintzinii Rost. |
бн |
+ |
+ |
||
|
Symphytocarpus flaccidus (Lister) Nann.-Brem. |
бн |
+ |
|||
|
Badhamia nitens Berk. |
? |
+ |
+ |
||
|
B. panicea (Fr.) Rost. in Fuckel |
бн |
+ |
+ |
||
|
Brefeldia maxima (Fr.) Rost. in Fuckel |
бн |
+ |
|||
|
Diderma alpinum Meylan |
? |
+ |
+ |
||
|
D. globosum Pers. |
бн |
+ |
+ |
||
|
D. hemisphaericum (Bull.) Hornem. |
бн |
+ |
+ |
||
|
D. niveum (Rost.) Macbr. |
бн |
+ |
+ |
||
|
D. ochraceum Hoffm. |
? |
+ |
+ |
||
|
D. radiatum (L.) Morgan |
бн |
+ |
+ |
||
|
D. testaceum (Schr.) Pers. |
бн |
+ |
+ |
||
|
Didymium minus (Lister) Morgan |
бн |
+ |
+ |
||
|
D. serpula Fr. |
бн |
+ |
+ |
||
|
Physarum bivalve Pers. |
бн |
+ |
+ |
||
|
P. compressum Alb. et Schw. |
бн |
+ |
+ |
||
|
P. confertum T. Macbr. |
бн |
+ |
+ |
||
|
P. decipiens Curt. |
бн |
+ |
+ |
||
|
P. dictyospermum Lister et G. Lister |
? |
+ |
+ |
||
|
P. galbeum Wingate |
? |
+ |
+ |
||
|
P. gyrosum Rost. |
бн |
+ |
+ |
||
|
P. murinum Lister |
? |
+ |
+ |
||
|
P. nudum Macbr. in Peck et Gilb. |
бн |
+ |
+ |
||
|
P. oblatum Macbr. |
бн |
+ |
+ |
||
|
P. pezizoideum (Jungh.) Pav. et Lag. |
н |
+ |
+ |
||
|
P. pusillum (Berk.) G. Lister |
бн |
+ |
+ |
Окончание табл. 4
|
Вид |
ГЭ |
Са+ |
ВП |
ВВП |
СМП |
|
P. rubiginosum Fr. |
бн |
+ |
+ |
||
|
P. stellatum (Massee) G. W. Martin |
? |
+ |
+ |
||
|
P. tenerum Rex |
бн |
+ |
+ |
||
|
P. vernum Somm. in Frees. |
бн |
+ |
+ |
||
|
Colloderma oculatum (Lipp.) G.Lister |
бн |
+ |
+ |
||
|
Cribraria purpurea Schr. |
бн |
+ |
+ |
||
|
C. splendens (Schr.) Pers. |
бн |
+ |
+ |
||
|
Lepidoderma tigrinum (Schr.) Rost. |
бн |
+ |
+ |
||
|
Physarum leucopus Link |
бн |
+ |
+ |
+ |
|
|
Licea tenera Jahn |
бн |
+ |
+ |
||
|
L. variabilis Schr. |
бн |
+ |
+ |
||
|
Lindbladia tubulina Fr. |
бн |
+ |
+ |
||
|
Lycogala exiguum Morgan |
бн |
+ |
+ |
||
|
L. flavofuscum (Ehr.) Rost. in Fuckel |
бн |
+ |
+ |
||
|
Physarum conglomeratum (Fr.) Rost. |
бн |
+ |
+ |
+ |
|
|
P. didermoides (Pers.) Rost. |
бн |
+ |
+ |
+ |
|
|
P. flavicomum Berk. |
бн |
+ |
+ |
+ |
|
|
P. globuliferum (Bull.) Pers. |
бн |
+ |
+ |
+ |
|
|
P. polycephalum Schw |
? |
+ |
+ |
+ |
|
|
P. psitacinum Ditmar in Sturm |
бн |
+ |
+ |
+ |
|
|
Reticularia splendens Morgan |
бн |
+ |
+ |
||
|
Stemonitis flavogenita E. Jahn |
бн |
+ |
+ |
||
|
S. pallida Wing. in Macbride |
бн |
+ |
+ |
||
|
S. smithii Macbr. |
бн |
+ |
+ |
||
|
Stemonitopsis typhina (F. H. Wigg.) Nann.- Bremek. |
бн |
+ |
+ |
||
|
Symphytocarpus amaurochaetoides Nann.- Brem. |
? |
+ |
+ |
||
|
S. confluens (Cooke et Ellis) Ing et Nann |
бн |
+ |
+ |
Примечание. ГЭ – географический элемент; Са+ – облигатные или факультативные кальцефилы; бн – бореально-неморальные, н – неморальные.
В ВВП отмечено 6 дифференциальных видов. Многие из них встречаются в пределах этой провинции редко. Для четырех видов ( Didymium iridis , Licea biforis , Physarum citrinum , P. famintzinii ) обнаруженные местонахождения приурочены к северной границе их ареалов в Европейской России.
Значительное своеобразие биоты СМП обусловлено широким распространением долинных ландшафтов с обнажениями карбонатных пород. Многие виды, отмеченные только в СМП, являются облигатными или факультативными кальцефилами (табл. 4). Особенно широко кальцефильные миксомицеты представлены на территории Ржевско-Старицкого Поволжья, где встречаются сложные комплексы долинных ландшафтов с выходами карбонатных пород. Здесь отмечено обилие микрониш с высокой влажностью по берегам рек и ручьев, в оврагах, которые являются местообитаниями миксомицетов. Полученные данные свидетельствуют о том, что зональные и секторальные особенности территории, специфика растительного покрова имеют для миксомицетов меньшее значение, чем характеристики конкретных микрониш.
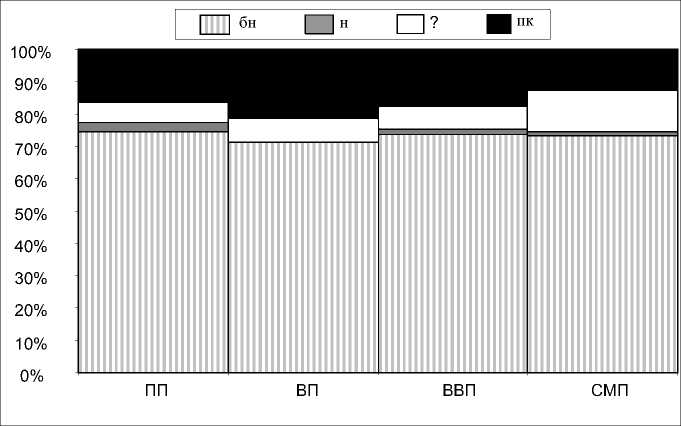
Рис. 4. Спектр географических элементов биот миксомицетов Валдайской и смежных физико-географических провинций [по: 15, с изменениями]:
бн – бореально-неморальные, н – неморальные, пк – полизональные
Среди кодифференциальных видов, отмеченных в ВП и СМП, представлена группа «реликтовых» горно-таежных криотолерантных миксомицетов аркто-бореально-альпиского элемента субаркто-горной дизъюнкции. Среди них Colloderma oculatum , Lepidoderma tigrinum , Barbeyella minutissima , представители рода Cribraria Pers. Эти виды нередко ассоциированы с печеночниками.
Некоторые кодифференциальные виды, зарегистрированные в ВВП и СМП, Licea tenera и Physarum psitacinum находятся на северной границе их ареалов в Европейской России. Остальные виды представляют бореальнонеморальную группу. По-видимому, часть из них может быть найдена в ВП при более детальном исследовании территории.
В специальном анализе нуждаются болотные массивы Тверской обл. В области уже зарегистрировано четыре вида рода Symphytocarpus Ing et Nann.-Bremek., редко встречающиеся в России, преимущественно на болотах таежной зоны.
В целом спектры географических элементов биот хорошо изученных провинций являются типичными для таежной зоны и характеризуются низкой специфичностью (рис. 4). Выявляется незначительное увеличение доли бореально-неморальных видов в ряду ВП – ВВП – СМП, в некоторой степени возрастает роль полизональных видов. При этом долевое участие первой группы в ПП меньше, чем в ВП и ВВП. По-видимому, изменение степени континентальности климата оказывает незначительное влияние на географическую структуру биоты миксомицетов в пределах лесной зоны. Об этом свидетельствует также состав спектра выделяемых групп географических элементов [28]. Миксомицеты являются компонентом биоты, географическое распределение разнообразия которого в большей степени зависит от характеристик конкретных микрониш. Долготные и широтные тренды слабо выражены.
Заключение
Таким образом, для дальнейшей разработки проблемы сопряженного биогеографического анализа разных компонентов биоты необходима реализация программ по комплексному изучению модельных территорий большого масштаба. В качестве такой территории может быть Русская равнина, в пределах которой представлены разные ботанико-географические зоны, геоморфологические районы и провинции. При сопряженном анализе разных компонентов целесообразно использовать физико-географическое районирование. Актуален поиск характеристик, показателей и индексов, позволяющих сравнивать разные биоты. Целесообразно выяснение зависимости выявляемых трендов от особенностей широтной дифференциации климата и растительного покрова, специфики геоморфологии, характера приуроченности разных таксономических групп к фитоценозам и микронишам. При предварительном анализе материала большое значение приобретает выявление географической специфики дифференциальных и кодифференциальных видов.
В пределах Валдайской и смежных физико-географических провинций зональные тенденции изменения общего уровня видового богатства и степени специфичности флор более четко проявляются у сосудистых растений, чем у мхов. Специфика бриофлор в большей степени определяется долготным положением территории. Степень оригинальности флор сосудистых растений в долготном направлении увеличивается при учете состава агамных комплексов, относящихся к родам Hieracium , Taraxacum .
Зональные тенденции в изменении параметров разнообразия лихенобиот Валдайской и смежных физико-географических провинций так же, как и у мхов, проявляются менее четко, чем долготные тренды, обусловленные увеличением степени континентальности климата. Уровни разнообразия и специфичности флор мхов и биот лишайников в определенной степени зависят от геоморфологических особенностей территории. Флоры и биоты провинций, в которых представлены крупные возвышенности, разнообразные ландшафты с обнажениями коренных материнских пород и россыпями валунов, характеризуются более высоким уровнем разнообразия и специфичности.
Географическое распределение разнообразия миксомицетов, по-видимому, в меньшей степени, чем у сосудистых растений, мхов и лишайников, обусловлено широтными и долготными особенностями территории. Уровень разнообразия миксомицетов в большей степени зависит от широты распространения специфических микрониш. Более богатые биоты формируются в физико-географических районах и провинциях, в которых обычны различные местообитания с обнажениями карбонатных пород.
Автор выражает глубокую благодарность О.В. Ребристой, Г.П. Урбанавичюсу, М.С. Игнатову, А.Н. Лебедеву за плодотворное обсуждение материалов.
