О состоянии изученности репродуктивной биологии желтопузика (Reptilia, Sauria, Anguidae) в Крыму
 в Крыму")
Автор: Кукушкин О.В., Кармышев Ю.В., Ярыгин А.Н., Шарыгин С.А.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 2 т.22, 2013 года.
Бесплатный доступ
Уровень изученности репродуктивной биологии желтопузика ( Pseudopus apodus ) на Крымском полуострове до настоящего времени остается крайне низким. Впервые для Крыма приводятся размерные характеристики яиц желтопузика, данные о сроках спаривания и откладки яиц. Обсуждается предположение о двухлетнем репродуктивном цикле самок.
Крым, размножение, двухлетний репродуктивный цикл
Короткий адрес: https://sciup.org/148314889
IDR: 148314889 | УДК: 598.112
Текст краткого сообщения О состоянии изученности репродуктивной биологии желтопузика (Reptilia, Sauria, Anguidae) в Крыму
также весьма ориентировочные данные о соотношении полов в популяциях (Кармышев, Кукушкин, 2001). Сведения о размножении желтопузика в других частях его обширного ареала, простирающегося от Адриатики на западе до озера Балхаш на востоке, и от Истрии, Крыма и Калмыкии на севере до Леванта и Афганистана на юге, отрывочны и зачастую противорчивы. Не будет преувеличением сказать, что P. apodus до сих пор остается одним из наименее изученных видов ящериц Северной Евразии, знание многих аспектов биологии которого основывается нередко на не вполне достоверных наблюдениях. Изучение размножения P. apodus сильно осложняется и низкой встречаемостью самок вследствие диспропорции в соотношении полов, либо половых различий в сезонной динамике уровня их активности. Так, по данным 1995-2012 гг., соотношение полов (♂♂: ♀♀) в Керченском Приазовье составило 4:1 (n=80), в Горном Крыму – 2,20:1 (n = 80). Таким образом, на одну самку в норме приходятся 3 самца. Между тем, для разработки эффективных мер по охране этого вида, в настоящее время сокращающего свою численность в Крыму почти повсеместно, и внесенного на этом основании в Красную книгу Украины в категории I – «исчезающий вид» (Кукушкiн, Котенко, 2009), необходимо располагать максимально четкими представлениями о его репродуктивных характеристиках, в конечном итоге, определяющих популяционную динамику. Данная работа не претендует на заполнение существующих многочисленных пробелов, но указывает направление дальнейших исследований.
МАТЕРИАЛ И МЕТОДИКА
Cроки спаривания регистрировали при наблюдениях в природе. Длину тела самок (Ln., мм) измеряли линейкой от кончика морды до анальной щели. Взрослых самок P. apodus отлавливали в пунктах с наиболее высокой плотностью популяций вида в Крыму (Кармишев, 1999; Котенко, Кукушкин, 2010). Одна беременная самка была добыта 21.07.2000 в окрестностях г. Инкерман (возвышенность Мекензиевы горы, территория Севастополя; 44о37`N, 33o39`E). Позже, в 2012 г., на азовском побережье Керченского полуострова были добыты 4 половозрелых самки P. apodus (Ln. 354, 364, 372 и 419 мм). Половину особей из этого числа отловили 29-30.05 в Булганакско-Осовинской степи (3-7 км к северу– северо-западу от с. Бондаренково; 45о29`N, 36o29`E), половину – 17-18.06 в Караларской (Чаганы-Чокраской) степи (12 км к северо-западу от пос. Багерово; 45о28`N, 36o12`E). С целью получения кладок добытые самки содержались в террариумах размерами 0,8×0,4×0,3 м, оборудованных лампами накаливания мощностью 25 Вт, обеспечивающих суточный перепад температуры воздуха от 20-22 ночью до 28-30°С днем. Единственную полученную в 2012 г. кладку инкубировали на керамзите при температуре воздуха и субстрата 25-30°С и влажности 80-90%. Промеры яиц (L. – длина; d. – диаметр) снимали штангенциркулем с точностью ±0,1 мм в первые дни после их откладки. Яйца взвешивались на лабораторных чашечных весах. Погрешность определения массы (W) составила 5 мг.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Откладка яиц как в Горном, так и Равнинном Крыму нами отмечена на протяжении июля. 22.07.2000 самка из Инкермана, добытая днем раньше, отложила 13 овальных белых яиц, находясь в герпетологическом мешке – прежде, чем ее поместили в террариум (рис. 1). Количество отложенных яиц незначительно превысило размер кладки, приводящийся в литературе как максимальный для вида – 10 яиц (Богданов, 1960; Банников и др., 1977; Саид-Алиев, 1979; Кукушкiн, Котенко, 2005; Хидиров, 2006; Arakelyan et al.. 2011) или 12 яиц (Bischoff, 1974; Obst, 1981). Однако, в нашем случае 2 яйца в кладке оказались неоплодотворенными. Размеры нормально равитых яиц приводятся в таблице; жировые яйца отличались несколько меньшими размерами и существенно меньшей массой: L. – 31,0-33,5 мм (X±Sx = 32,3±1,25); d. – 15,0-16,0 мм (X±Sx = 15,5 + 0,05); L./ d. – 2,07-2,09 мм (X±Sx = 2,08 + 0,02); W – 4,2-4,8 г (X±Sx = 4,50±0,03).
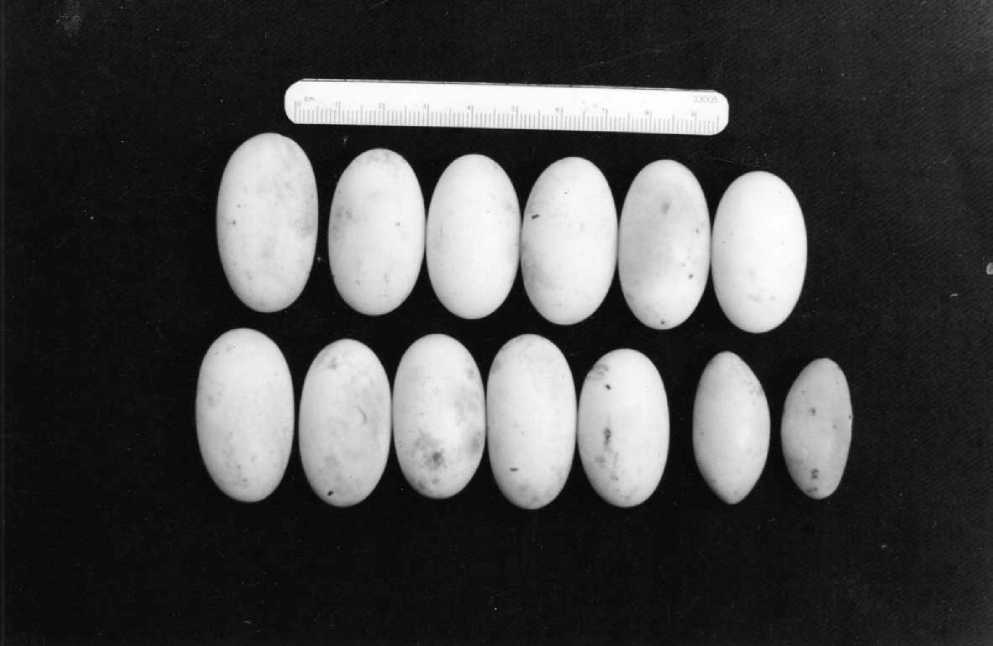
Рис. 1. Кладка самки P. apodus из окрестностей г. Инкерман (Севастополь). Жировые яйца расположены в нижнем ряду справа
8.07.2012 одна из самок, отловленных в Караларской степи 17.06, отложила (порционно: с интервалом в несколько часов) 8 яиц, 6 из которых были нормальными, а 2 жировыми – так же, как и в предыдущем случае (табл.). Размеры свежеотложенных яиц желтопузиков из исследованных нами кладок в общем соответствуют литературным данным (Сыроечковский, 1958; Богданов, 1960; Bischoff, 1974; Банников и др., 1977; Obst, 1981; Evans, 1995), но для Крыма приводятся впервые. Н.Н. Щербак (1966), по результатам вскрытия недатированной особи с Южного берега Крыма (Ялта) из коллекции А.А. Браунера, указывает меньшие размеры яиц P. apodus: 25,2-34,3×15,0-17,2 мм (n=10). Очевидно, яйца не были вполне сформированы на момент поимки особи либо уменьшили свои размеры вследствие длительного пребывания в фиксирующем растворе. G. Evans (1995) при сходных размерах (около 40×20 мм) жизнеспособных яиц, полученных при лабораторном разведении от самки, содержащееся в неволе около 15 лет, указывает их меньшую массу – 8-12 г (n=10).
Таблица
Размеры и масса яиц в кладках P. apodus из Крыма
|
Популяция |
n |
Статистические параметры |
Размеры и пропорции яиц |
W, g |
||
|
L., мм |
d., мм |
L./d. |
||||
|
горная (Севастополь) |
11 |
Lim. X±S x CV, % |
34,5-39,3 36,5±0,45 4,06 |
19,8-2,1 20,7±0,24 3,77 |
1,68-1,93 1,76±0,02 2,26 |
18,4-21,5 19,4±0,31 2,78 |
|
равнинная (Приазовье) |
6 |
Lim. X±S x CV, % |
31,2-34,0 33,1±0,40 2,94 |
17,0-9,8 18,6±0,42 5,48 |
1,69-1,84 1,78±0,02 3,17 |
– |
|
все |
17 |
Lim. X±S x CV, % |
31,2-39,3 35,3±0,51 5,93 |
17,0-22,1 20,0±0,32 6,63 |
1,69-1,93 1,77±0,02 3,50 |
– |
Несмотря на высоко достоверные отличия значений метрических параметров (длины и диаметра) яиц (P<0,001) в кладках из Горного и Равнинного Крыма, отношение длины к диаметру в обеих кладках оказалось близким. К сожалению ограниченные данные, которыми мы располагаем, недостаточны для сравнения плодовитости самок и размеров яиц в горной и равнинной популяциях P. apodus .
Никаких проявлений заботы о потомстве (охраны кладки), описанной у P. apodus многими авторами (Bischoff, 1974; Кудрявцев и др., 1991; Green, Rodriguez, Powell, 2006; Milto, 2010; Arakelyan et al., 2011), у самки из Караларской степи не отмечалось, и до 20.07 все яйца погибли – по-видимому, вследствие грибкового поражения. При вскрытии трех яиц, погибших последними, найдены эмбрионы на ранних стадиях развития, по степени сформированности сравнимые с эмбрионами роющей безногой ящерицы Calyptommatus sinebrachiatus Rodrigues, 1991 (сем. Gymnophthalmidae, инфраотряд Scincomorpha) седьмой стадии нормального развития (см.: Roscito, Rodrigues, 2012).
7.09.2012 одна из содержащихся в лаборатории самок (Ln. 364 мм) из Керченского Приазовья пала. При вскрытии животного были обнаружены белые овоциты (по 6 в каждом яичнике) диаметром 2-3,5 мм (в среднем 2,81
мм). От оставшихся двух ящериц, нормально питавшихся в неволе, кладки вплоть до ноября 2012 г. получить не удалось, и они были определены на искусственную зимовку. Таким образом, лишь одна из 4 добытых в репродуктивный период на Керченском полуострове самок P. apodus отложила яйца лишь одна (25%).

Рис. 2. Спаривание P. apodus (Мекензиевское лесничество, Севастополь). Самец во время копуляции челюстями удерживает самку за голову
Описанная картина представляет значительный интерес. Можно было ожидать, что в конце мая – начале июня в яйцеводах половозрелых самок должны находиться развивающиеся яйца на стадии интенсивного вителлогенеза. Судя по динамике гонад, спаривание у P. apodus в Крыму, как и в иных частях ареала, бывает, как правило, в середине апреля – середине мая (Сыроечковский, 1958; Богданов, 1962; Щербак, 1966; Саид-Алиев, 1979; Пицхелаури, 1985; Сартаева, Акрамов, 1985, Туниев, Туниев, 1987). Это подтверждается и нашими наблюдениями. Так, спаривающиеся желтопузики (3 пары в течение одного дня) были отмечены 3.05.1998 близ Инкермана (рис. 2). Пары ящериц, предположительно проявлявших сексуальную активность, встречали также 16.05.1961 близ Сюренских гротов (окрестности с. Куйбышево Бахчисарайского района) (Щербак, 1966), 16.04.1996 на юговосточной окраине г. Балаклава в балке Кефало-Врисси и 1.05.1996 на Фиолентском оползневом побережье (южный берег Гераклейского полуострова, черта Севастополя). На Керченском полуострове спаривание нами не наблюдалось, однако нет никаких оснований предполагать, что его сроки существенно отличаются от таковых в южной части Горного Крыма, так как более холодная зима на равнинном побережье Азова в норме компенсируется более ранним, чем на черноморском берегу, наступлением весны. Об отсутствии популяционных отличий в масштабах Крымского полуострова говорят и данные по динамике гонад (Щербак, 1966).
В некоторых частях ареала, судя по резкому увеличению семенников у самцов, спаривание бывает еще раньше: в конце марта – апреле (Яковлева, 1964), в то время как последние спаривания проходят в начале – середине июня (Богданов, 1960; Мусхелишвили, 1970). Однако, несмотря на продолжительный период спаривания, длящийся, по крайней мере, 1,5-2 месяца, кладки (либо самки с готовыми к откладке яйцами) всеми перечисленными авторами регистрировались в июне и июле (Сыроечковский, 1958; Богданов, 1960; Яковлева, 1964; Мусхелишвили, 1970; Саид-Алиев, 1979; Пицхелаури, 1985; Obst, 1981), и никогда – в мае, даже в частях ареала, расположенных значительно южнее Крыма. Так, при вскрытии в середине мая двух самок из долины реки Мургаб (южный Туркменистан) в их яичниках были обнаружены желтые фолликулы диаметром 5-8 мм (Богданов, 1958).
Несмотря на резкое потепление в конце апреля – начале мая 2012 г. (суточный максимум температуры воздуха доходил до до 27-30оС), представляется практически невероятным, что яйца были отложены большинством самок в мае – как говорилось выше, весенние кладки у P. apodus с территории бывшего Советского Союза до сих пор никем не регистрировались, и не известны также у западного (балканомалоазиатского) подвида P. a. thracius (Obst, 1978). Кроме того, при допущении возможности откладки яиц в конце весны вызывает удивление значительный временной интервал между основной частью кладок в мае (гипотетических) и единственной наблюдавшейся июльской. Противоречит такому предположению и довольно высокая активность самок в период отлова – если допустить, что охрана кладки у желтопузика является нормой, а не редким исключением.
Основная часть кладок не могла прийтись и на начало июня, так как в этом случае самки, добытые в конце мая, несомненно, отложили бы яйца в неволе. Резорбироваться за столь короткий отрезок времени яйца не могли даже в самых неблагоприятных условиях. Например, известен случай, когда при вскрытии 7.07.2008 содержавшейся без кормления самки, добытой месяцем раньше на полуострове Маяк севернее Керчи, было обнаружено единственное сформированное яйцо размерами 37,9×15,9 мм. Предполагать, что сроки откладки яиц в двух близлежащих популяциях Керченского Приазовья, расположенных на одной широте на удалении 20 км друг от друга, могут существенно отличаться, также не приходится, несмотря даже на определенные биотопические различия (в Караларской степи ящериц отлавливали на песчаной дюне и скалистых склонах приморских амфитеатров, в то время как в Булганакско-Осовинской степи желтопузик населяет почти совершенно лишенную скальных выходов и каменистых россыпей местность и использует в качестве убежищ норы грызунов). Описание аналогичной ситуации содержится в обстоятельной работе Е.Е. Сыроечковского (1958): в Чечне из числа 22 вскрытых в период с мая до середины июля 1950 г. самок готовые к откладке яйца были обнаружены лишь у двух (9,1%), причем у обеих – в начале июля (5.07 и 11.07). У прочих исследованных ящериц состояние фолликулов оставалось стабильным (диаметр – 3-5 мм) на протяжении всего периода наблюдений, поэтому этот автор высказал мнение, что июльские кладки скорее всего являются следствием задержки размножения у отдельных особей, однако доказательств этому не представил. По нашему мнению, такая картина может быть объяснена при допущении, что самки желтопузика приносят потомство не ежегодно и для них характерен двухлетний репродуктивный цикл. Впрочем, в доступной нам литературе имеются указания на ежегодное размножение P. apodus (Богданов, 1965; Meiri, Brown and Sibly, 2012), однако, необходимо учитывать, что размножение желтопузика, как и большинства других представителей семейства Anguidae, на сегодняшний день изучено чрезвычайно слабо, а характер представленных данных ясно указывает на то, что они не могут претендовать на абсолютную точность и, следовательно, не являются неоспоримыми.
Неежегодное размножение достаточно распространено в некоторых семействах Sauria – особенно среди видов, у которых предполагается охрана кладки самкой. Например, оно отмечено у крупных сцинков Novoeumeces schneideri (Daudin, 1802) и Eurylepis taeniolatus Blyth, 1854 (Атаев, 1985). Описан двухгодичный репродуктивный цикл и для ряда видов подсемейства Anguinae. Так, значительная часть самок веретеницы ломкой ( Anguis fragilis Linnaeus, 1758) ежегодно не участвует в размножении, причем это происходит не только в напряженных экологических условиях европейской субальпики и близ северной границы ареала, но и в районах, с климатом, близким к оптимальному для этого вида (Capula et al., 1992; Ferreiro, Galán, 2004). Периодичность размножения A. fragilis имеет нерегулярный характер и зависит не только от широты местности, но и от продолжительности периода активности, поэтому доля неразмножающихся самок существенно изменяется от года к году и варьирует в различных западноевропейских популяциях от 10-20 до 50-70%. В связи с этим отметим, что вторая половина зимы 2011-2012 гг. в восточном Крыму выдалась суровой (средняя температура февраля ниже среднемноголетней более, чем на 5°С), а весна запоздала (до середины марта минимальные температуры воздуха были стабильно отрицательными). На азовском побережье Крыма желтопузик выходит из зимовки обычно в начале – середине апреля, иногда в конце марта (Кукушкин, 2006). Таким образом, условия зимы и ранней весны, безусловно, могут оказывать влияние и на его репродуктивную активность.
Установлено также, что ежегодно размножается лишь около половины самок стройной панцирной веретеницы, Ophisaurus attenuatus Cope, 1880 (Fitch, 1989) – представителя сестринской по отношению к кладе Anguis – Pseudopus группы (Macey et al., 1999). H.S. Fitch (1989) предположил, что охрана кладки, известная у этого крупного вида североамериканских желтопузиков, является весьма энергозатратной, поскольку в период инкубации самки редко покидают подземные убежища и практически не принимают пищу. Поэтому для накопления резервов организма, достаточных для реализации репродуктивной функции, требуется длительный период покоя – не менее года. Низкая встречаемость в природе беременных самок P. apodus также может быть связана с их пониженной активностью ввиду нахождения в убежищах и охраны кладок, и в действительности соотношение особей, принимающих и не принимающих участие в размножении, может быть близким к равному. Заметим здесь же, что причиной отмечавшейся большинством авторов необычайно низкой встречаемости сеголеток P. apodus может быть не только скрытный образ жизни и/или высокая смертность, но также и изначально невысокая их численность, обусловленная неежегодным участием в размножении всех самок репродуктивного возраста.

Рис. 3. Эмбрионы из разоренной лисицей поздней кладки P. apodus, (окрестности пос. Сотера, Алушта)
Достоверно не известна и продолжительность инкубации яиц P. apodus в природе. По литературным данным, сеголетки появляются с конца июля – начала августа по сентябрь включительно (Банников и др., 1977; Кармишев, 1999; Туниев, Туниев, 2007; Arakelyan et al., 2011). На Южном берегу Крыма в долине р. Алака (окрестности пос. Сотера, территория Алушты) состоящая как минимум из 7 яиц с полностью сформированными крупными эмбрионами кладка желтопузика, разоренная, судя по оставленным следам, лисицей, была обнаружена 28.09.2005 (рис. 3). В Никитском ботаническом саду недавно вышедших из яиц сеголеток со следами пупочных канатиков отмечали 23.10.2003 (5 особей в полости под каменной плитой проезжей дороги; здесь же находилась взрослая самка), 2.11 и 5.11.2003 (по одной особи), 21.10.2005 (2 особи неподалеку друг от друга). Таким образом, в условиях Южного берега Крыма успешный выход сеголеток может происходить и в конце сентября – октябре.
Если исходить из того, что последние кладки у P. apodus приходятся на конец июля, приходим к выводу, что продолжительность инкубации в природе достигает приблизительно 60 суток и более (до 80-90 суток), то есть ее продолжительность больше, чем это известно по литературным данным – 30-60 суток, чаще 45-50 суток (Petzold, 1971; Obst, 1981; Кудрявцев и др., 1991; Evans, 1995; Arakelyan et al., 2011). Сходные данные получены и для трех видов североамериканского рода Ophisaurus Daudin, 1803 – 39-53, иногда до 60-75 суток (Fitch, 1989; Witz and Wilson, 1993). Можно допустить также, что откладка яиц у P. apodus в исключительных случаях продолжается в августе. Такая ситуация кажется вполне вероятной. Например, подавляющее большинство кладок у O. attenuatus приходится на июнь – первую половину июля, но описан и единичный случай откладки яиц в начале октября (Fitch, 1989). Откладка яиц в первой половине августа регистрировалась нами у многих обитающих в Крыму видов ящериц: Mediodactylus kotschyi (Steindachner, 1870), Darevskia lindholmi (Szczerbak, 1962), Eremias arguta (Pallas, 1773), Podarcis tauricus (Pallas, 1814) – как правило, в годы с экстремально суровой зимой и/или поздней весной (Кукушкин, 2007; Kukushkin, 2007).
Продолжительность беременности у самок желтопузика в природе не известна. Лишь опираясь на известные для Крыма сроки спаривания (середина апреля – середина мая) и откладки яиц (середина июня – конец июля), можно рассчитать, что длится она приблизительно 60-80 дней. Немногочисленные наблюдения за размножением желтопузика в неволе подтверждают эти предварительные выводы: развитие яиц в теле самки длится около 10-11 недель (Rogner, 1992; Evans, 1995). Таким образом, если период спаривания в Крыму, как и в других частях ареала (например, на Черноморском побережье Кавказа и в Восточной Грузии), продолжается в конце мая – начале июня (например, при позднем вступлении в процесс размножения особей, недавно достигших половой зрелости), последние кладки вполне могут приходиться на начало августа. Отсутствие данных о кладках в конце лета по всему ареалу P. apodus может объясняться сезонной динамкой активности этого вида, особенно ясно выраженной у самок. В засушливой второй половине лета встречаемость желтопузика падает практически до нуля, оставаясь низкой вплоть до ухода на зимовку на протяжении октября – первой декады ноября (наиболее поздняя дата встречи взрослой особи в Крыму, в урочище Микро-Яло к юго-востоку от г. Балаклава – 10.11.1996). Эта особенность послужила поводом для предположения о наличии у желтопузика летней спячки (Банников, 1954), причем в отдельные годы складывается впечатление, что летняя спячка может переходить в зимнюю (Богданов, 1960, 1965; Яковлева, 1964; Мусхелишвили, 1970; наши данные).
Обилие нерешенных вопросов указывет на необходимость уделить изучению репродуктивной биологии желтопузика самое пристальное внимание.
Список литературы О состоянии изученности репродуктивной биологии желтопузика (Reptilia, Sauria, Anguidae) в Крыму
- Атаев Ч.А. К размножению горных пресмыкающихся Туркменистана // Вопросы герпетологии. Л.: Наука, 1985. С. 11.
- Банников А.Г. Материалы по биологии земноводных и пресмыкающихся южного Дагестана // Учен. зап. Моск. гор. пед. ин-та им. Потемкина. 1954. Т. 28. C. 75-88.
- Банников А.Г., Даревский И.С., Ищенко В.Г., Рустамов А.К., Щербак Н.Н. Определитель земноводных и пресмыкающихся фауны СССР. М.: Просвещение, 1977. 415 с.
- Богданов О.П. Ящерицы долины реки Мургаб // Тр. ин-та зоологии и паразитологии. Т. 5. Зоол. сб. Ташкент: Изд-во АН Уз. ССР, 1958. С. 67-94.
- Богданов О.П. Земноводные и пресмыкающиеся. Ташкент: Изд-во АН УзССР, 1960. 259 с. (Фауна Узбекской ССР; Т. 1).
