О возможности использования эффекта кластеризации в системах связанных отображений для описания динамики метапопуляций

Автор: Кулаков М.П.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Региональный прогноз. Математическое моделирование
Статья в выпуске: 2 т.17, 2014 года.
Бесплатный доступ
Работа посвящена изучению явления кластеризации, проявляющегося в системах связанных отображений, рассматриваемых в качестве моделей динамики метапопуляций. Изучается возможность агрегации элементов каждого кластера суммарным или средним значением всех переменных в кластере. Показано, что в случае полностью синхронной или близкой к ней динамики в кластере, такие агрегированные величины могут быть описаны или аппроксимированы системой намного меньшего числа переменных (по числу кластеров). Предлагается использовать предложенную аппроксимацию для описания динамики реальных метапопуляций.
Метапопуляция, мультистабильность, системы связанных отображений, синхронизация, кластеризация, аппроксимация
Короткий адрес: https://sciup.org/14328877
IDR: 14328877 | УДК: 517.9:574.34
Using of clusterization effect in coupled map lattices for the description of real metapopulations dynamics
The article deals with the phenomenon of clusterization in coupled map lattices considered as simple models of metapopulation dynamics. The author investigates the opportunity of cluster elements aggregation by a summary or average value of all variables in the cluster. It is shown that in case of completely synchronous (or close to it) dynamics, such aggregate values can be described or approximated with the system of a considerably less in number variables (according to the number of clusters). It is proposed to use this approximation for the description of real metapopulations dynamics.
Текст научной статьи О возможности использования эффекта кластеризации в системах связанных отображений для описания динамики метапопуляций
Хозяйственная деятельность человека оказывает существенное влияние на среду обитания диких животных и, как следствие, воздействует на динамику их численности. Направления хозяйственной деятельности достаточно разнообразны, к ним относятся рубки леса, освоение угодий сельскохозяйственными предприятиями, развитие дорожной сети, разработка полезных ископаемых и т.д.
Рубки леса главного пользования - один из основных антропогенных факторов, в значительной мере трансформирующий среду обитания лесных животных, следовательно, существенно меняющий состав, численность и соотношение видов лесной фауны [6]. Рубки леса оказывают весьма неоднозначное влияние на условия обитания диких животных. С одной стороны, условия жизни для коренных лесных обитателей ухудшаются. Например, резкое сокращение количества плодоносящих деревьев уменьшает качество кормовой базы, а открытость территории делает животных уязвимыми для браконьеров. С другой стороны, в леса, пройденные рубками, проникают ранее несвойственные им виды животных, в ряде случаев при этом увеличивается фаунистическое разнообразие [6]. Вырубка темнохвойных и смешанных лесов приводит к появлению обильного подроста на стадии естественного восстановления древесной и кустарниковой растительности, разрастаются подлесок и травы. При этом сохранившиеся деревья и ягодные кустарники лучше плодоносят [5]. Происходящие на вырубках сукцессионные процессы способствуют росту численности древоядных копытных (косуля, изюбрь и лось), так как большую часть года их пищевой рацион составляют побеги древесных растений [1]. Обилие мышевидных грызунов на лесосеках привлекает не только колонка и лисицу, но и соболя [5]. Однако продолжительность существования таких благоприятных угодий зависит от площади и ширины лесосеки и условий возобновления. При переходе лесонасаждений в стадию жердняков запас древесно-веточных кормов сокращется, следовательно, уменьшается численность животных.
Другой важный аспект хозяйственной деятельности заключается в освоении и эксплуатации угодий сельскохозяйственными предприятиями. Сельскохозяйственные земли, представляющие собой сочетание пашен с сенокосами, зарослями кустарников и колко-лиственного леса, являются хорошими кормовыми угодьями и служат средой обитания для многих диких животных (например для косули, лисицы, колонка). Однако, несмотря на наличие сельскохозяйственных культур, которые благоприятствуют обитанию животных, осенняя вспашка полей, браконьерский пресс и периодически повторяющиеся весенние и осенние палы, уменьшающие площади естественных лесов, ухудшают защитные и гнездовые условия в сельскохозяйственном ландшафте [8, 17].
Цель данной работы - выполнить количественный анализ динамики численности охотничьих животных в зависимости от таких видов хозяйственной деятельности человека, как рубки леса главного пользования и освоение угодий сельскохозяйственными предприятиями (на примере Еврейской автономной области). Результатом воздействия совокупности разнородной хозяйственной деятельности на лес (основную среду обитания диких животных) и сопутствующих ей пожаров является изменение состава и площади лесных насаждений. В связи с этим в данной работе особое внимание уделено анализу динамики площади основных лесообразующих пород в контексте ее влияния на изменение численности диких животных.
Объектами исследования являются следующие виды животных: медведь бурый уссурийский (Ursus arctos lasiotus Gray, 1867), рысь (Fells lynx Linnaeus, 1758), лиса длинномордая уссурийская (Vulpes vulpes dolichocrania Ognev, 1926), соболь баргузинский (Martes zibellina princes Birula, 1922), колонок дальневосточный (Mustela sibiricamanchurica^Qrnss., 1911), лось уссурийский (Alces alces cameloides Milne-Edwards, 1867), изюбр (Cervus elaphus xanthopigus Milne-Edwards, 1867), кабан уссурийский (Sus scrofa ussuricus Heude, 1888), косуля маньчжурская (Capreolus pygargus bedfordi Thomas, 1908), заяц-беляк приамурский (Lepus timidus mordeni Goodwin, 1933), заяц маньчжурский (Lepus mandschuricus mandschuricus Radde, 1861).
Данная работа является продолжением начатых ранее исследований по изучению динамики численности охотничье-промысловых видов животных. В предыдущих работах [14-16,23] были проанализированы основные тенденции популяционной динамики, а также исследовано влияние кормовых запасов на динамику численности популяции белки, высоты снежного покрова, жидких осадков и температуры воздуха на динамику численности копытных в ЕАО. В настоящей статье акцент смещен на изучение взаимосвязи между популяционной динамикой и хозяйственной деятельностью человека, трансформирующей среду обитания диких животных.
Материалы и методы исследования
В работе использовались материалы по животному миру, земельным и лесным ресурсам:
-
- годовые отчеты по зимнему маршрутному учету диких копытных на территории Еврейской автономной области (ЕАО) с 1981 по 2014 гг. [2, 3, 9, 13];
-
- площадь, покрытая лесом по преобладающим породам (1997-2013 гг); [3, 10-13, 19];
-
- рубки леса главного пользования (площадь вырубки леса с 1995 по 2009 гг. и объем вырубленной ликвидной древесины с 1993 по 2013 гг); [2, 3, 10-12, 19, 20];
-
- посевные площади сельскохозяйственных культур (1995-2013 гг.) [18, 19, 21].
Для количественного анализа влияния внешних факторов на динамику численности животных применены методы корреляционного и регрессионного анализа и математического моделирования, при котором использовалась модифицированная модель Рикера с учетом внешних факторов [22].
Обозначим через хп численность популяции в и-ом году, HW- площадь твердолиственных пород, SW- площадь мягколиственных пород, С - площадь посевных сельскохозяйственных культур, WF - объем вырубленной ликвидной древесины. Параметры h, s, с, w характеризуют интенсивности влияния на популяционные процессы изменения площадей твердолиственных, мягколиственных и посевных сельскохозяйственных культур и объема вырубленной ликвидной древесины, соответственно. Также рассматривается влияние общей площади лиственных пород D (D=HW+SW), интенсивность ее воздействия выражается параметром d. Тогда модифицированная модель Рикера с учетом внешних факторов имеет вид:
Ли+1—и0Лие ’ или хй+1 = аох„е
где
(или
d-Di.+w-WFi1+c-Ci. х ап = а^е п « «) является коэффициентом попол нения популяции за счет процессов естественного вое-производства и миграции в и-ом году. Коэффициент bQ отражает влияние на популяцию плотностно-зависимых факторов, а величина Ьп = с ь* характеризует выживаемость популяции в и-ом году. Значения коэффициентов а и v меняются в зависимости от флуктуаций внешних факторов. В работе также были вычислены средние значения коэффициентов пополнения ари выживаемости v .
ср
Модифицированную модель Рикера (1) можно представить в виде линейной функции 1и(х /х^ от х HWn, SWn, WFrf С В этом случае оценка параметров модели сводится к вычислению коэффициентов множественного уравнения регрессии и может быть осуществлена с помощью любого доступного статистического пакета (например, Microsoft Excel). Кроме этого, оценка параметров моделей осуществлялась численно методом Левен-берга-Марквардта в программе MathCAD 14. Использование моделей (1)-(2) позволяет выделить те факторы, которые оказывают наибольшее влияние на годовые изменения численности популяций. Для полученных математических моделей, включающих комбинации факторов, вычислялись коэффициенты детерминации (R2) и ошибки аппроксимации (А). Коэффициент детерминации является мерой качества аппроксимации, те. чем больше R2, тем сильнее взаимосвязь между статистическими данными и модельным приближением к ним. Средняя ошибка аппроксимации Л показывает на сколько процентов в среднем расчетные значения отклоняются от фактических. Также качество моделей определялось согласно значениям /-критерия Фишера.
Результаты исследования
В настоящей работе сначала анализируются колебания численности диких животных вследствие изменения площадей лесонасаждений. Затем исследуется воздействие рубок леса главного пользования и освоения угодий сельскохозяйственными предприятиями на динамику численности рассматриваемых видов. Потом рассматривается комплексное влияние исследуемых показателей на флуктуации численности животных.
Анализ взаимосвязи между изменениями площади лесонасаждений и колебаниями численности охотничьих животных
Лес является основной средой обитания диких животных. Лесная растительность разносторонне влияет на состав и структуру диких животных - как кормовая база, как место для укрытия и как исходный пункт для размножения и расселения животных [24-25]. В связи с этим естественно предположить, что изменение площади лесных насаждений приводит к изменению ареала обитания животных, тем самым воздействуя на динамику их численности.
На рис. 1 представлена динамика площади основных групп лесообразующих пород: хвойных, твердолиственных и мягколиственных на территории ЕАО за период 1965-2013 гг.
Как видно из рис. 1, в период 1965-2013 гг. наиболее неравномерно изменялась площадь хвойных лесов,
650 з й 600 -
О 550 - д g 500 -
Й 450 - н 400 -
350 - о
300 - х
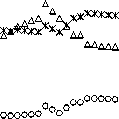
1960 1970 1980 1990 2000 2010 год д Хвойные ° Твердолиственные * Мягколиственные
Рис. 1. Динамика площадей хвойных, твердолиственных и мягколиственных пород деревьев представленных следующими лесообразующими породами: сосна, ель, пихта, лиственница, кедр корейский. По данным учета лесного фонда на начало 2014 г. [3], площадь насаждений из хвойных пород осталась в целом на уровне 1965 г. Вместе с тем, начиная с 2004 г, в ЕАО наблюдается сокращение площади хвойных насаждений. Площади насаждений из твердолиственных (дуб высокоствольный, ясень, клен, вяз и другие ильмовые, береза каменная) и мягколиственных (береза, осина, ольха серая, липа, тополь, ивы древовидные) пород за 1965-2013 гг. увеличились в 1,3 и 2 раза соответственно. По-видимому, в ЕАО продолжается многолетняя тенденция замещения хвойных пород лиственными (замены площади хвойных насаждений на площадь лиственных), в основном в результате рубок хвойных деревьев и лесных пожаров в хвойных и хвойно-широколиственных лесах [3, 7].
Для выявления статистической зависимости между оценками численности животных и площадью лесообразующих пород (по группам пород) был проведен корреляционный анализ, результаты которого приведены в табл. 1.
Согласно результатам корреляционного анализа (табл. 1) выявлено, что увеличение площади хвойного хозяйства в период 1997-2003 гг. благоприятно сказалось на популяционной динамике копытных, рыси и соболя спустя почти десятилетие. Вместе с тем, между численностью рассматриваемых видов и площадью хвойных насаждений в год, предшествующий размножению животных, был получен отрицательный коэффициент корреляции, то есть, несмотря на уменьшение хвойного хозяйства, поголовье животных увеличивается. Вероятно, для них так сложились условия среды обитания в пределах автономии, что сокращение площади хвойных насаждений с 2004 г. по настоящее время не препятствует росту их численности.
По результатам корреляционного анализа (табл. 1) установлено, что увеличение площади твердолиственных пород способствует росту численности рассматриваемых животных. Твердолиственные группы пород в ЕАО почти на 80% представлены дубом монгольским [3]. Дуб монгольский не только основная лесообразующая, но и главная кормовая порода в дубняках. В годы урожая желудей в дубняках в течение всей зимы обитают многие виды животных. Желуди имеют высокую кормовую ценность и охотно поедаются медведем, кабаном, изюбрем, косулей и другими животными [8]. Кормовые и защитные качества дубняков значительно повышают обильно произрастающие в подлеске лещина разнолистная и леспедеца двухцветная. Хорошо развита в дубняках травянистая растительность, представленная злаками и разнотравьем [8]. В дубняках и других лиственных лесах обитает рысь, так как там присутствуют большие запасы зайцев, косуль, грызунов, являющихся основным кормом для рыси [5].
Высокие статистически значимые положительные коэффициенты корреляции также были получены между численностью животных и площадью мягколиственных пород (табл. 1). По преобладанию пород в древостоях площади мягколиственных насаждений распределяются следующим образом (%): береза -63, осина -18, липа -16, ольха серая - 2, ивы древовидные -0,6, тополь - 0,4 [3]. Данные виды являются ценными кормовыми растениями для древоядных копытных [4]. Бе-
Таблица 1
Результаты парной корреляции между численностью диких животных в и-год и площадью лесообразующих пород в (и-^)-год, где к - номер лага
|
Основные породы / Виды копытных |
Изюбрь |
Косуля |
Кабан |
Медведь бурый |
Рысь |
Соболь |
|
Хвойные породы (И) |
-0,5767 (0,05) |
-0,6696 (0,01) |
-0,543 (0,05) |
* |
* |
-0,7957 (0,01) |
|
Хвойные породы (к=8 для косули, к=9 для кабана, рыси, соболя) |
* |
0,7997 (0,01) |
0,8519 (0,01) |
0,618 (0,05) |
0,8637 (0,01) |
0,8827 (0,01) |
|
Твердолиственные породы (к=4 для изюбря, рыси, соболя, к=\ для остальных) |
0,5895" (0,05) |
0,961 (0,01) |
0,8524 (0,01) |
0,7327 (0,01) |
0,7908 (0,01) |
0,9111 (0,01) |
|
Мягколиственные породы (к=4 для изюбря, рыси, соболя, к=\ для остальных) |
0,5825" (0,05) |
0,9563 (0,01) |
0,8119 (0,01) |
0,7183 (0,01) |
0,8466 (0,01) |
0,9112 (0,01) |
|
Лиственные породы (к=4 для изюбря, рыси, соболя, к=\ для остальных) |
0,5913" (0,05) |
0,9648 (0,01) |
0,8368 (0,01) |
0,73 (0,01) |
0,8276 (0,01) |
0,9197 (0,01) |
Статистически незначимый коэффициент корреляции реза белая (реже осина) является доминирующей породой в мелколиственных лесах. Мелколиственные леса -это вторичные насаждения, возникшие на месте выгоревших и вырубленных хвойных и хвойно-широколиственных лесов [8, 13]. В подлеске мелколиственных лесов растут ивы, рододендрон, шиповник, спирея, а в травяном покрове - осоки, злаки, разнотравье. Хорошие кормовые условия складываются в мелколиственных лесах для копытных из-за обилия травяной растительности и запасов древесно-веточного корма. Из плодово-ягодных растений в этих лесах произрастает брусника, черемуха, рябина, шиповник, смородина, жимолость. Однако в этих лесах для животных существуют плохие защитные условия, выраженные незначительной захламленностью [8]. Обилие грызунов и ягодники привлекают в лиственные леса также соболя, несмотря на то, что он в большей степени представитель таежный фауны.
Анализ популяционной динамики в зависимости от динамики площади лесных насаждений был дополнен модельным анализом. Результаты оценки параметров модифицированного варианта модели Рикера (1) с учетом площади мягколиственных и твердолиственных пород представлены в табл. 2.
Сравнение коэффициентов, характеризующих интенсивности влияния изменения площади лиственных пород на численность животных, показало, что воздействие динамики площадей твердолиственных пород на развитие большинства рассматриваемых популяций сильнее, чем мягколиственных пород. Высокие значе ния коэффициентов детерминации и низкие ошибки аппроксимации, полученные при сравнении динамики модельных и фактических численностей косули (табл. 2), позволяют судить о существенной взаимосвязи динамики площадей этих лесов и изменения численности вида. Завышенный коэффициент пополнения популяции (3,23,6), по-видимому, отражает не столько высокий уровень естественного прироста популяции, сколько повышенную миграционную активность животных. Для остальных видов животных модель (1) с учетом площади лиственных насаждений несколько хуже описывает динамику их численности. Кроме того, это связано с тем, что на рост поголовья данных популяций, кроме рассмотренных показателей, также влияют не включенные в модель (1) прочие факторы. Кроме этого, при описании динамики численности рыси посредством модели (1) с учетом изменения площадей твердолиственных пород получен низкий коэффициент детерминации, свидетельствующий о весьма слабой взаимосвязи между статистическими данными и модельным приближением к ним.
Влияние рубки леса на динамику численности охотничьих животных
Заготовка древесины является важнейшим видом лесопользования. Основная часть древесины изымается в процессе рубок главного пользования. Рубки леса главного пользования проводятся в спелых и перестойных древостоях для заготовки древесины и направлены на восстановление лесов хозяйственно-ценными древесными породами и эффективную эксплуатацию [2]. Ди-
Таблица 2
Оценки параметров модели (1) с учетом изменения площадей твердолиственных и мягколиственных пород
|
Вид ЖИВОТНОГО |
Коэффициенты модели (1) |
«ср |
Vcp |
R2 |
Л, % |
|||
|
«0 |
h |
5 |
bo |
|||||
|
Косуля |
1,9-10"4 |
0,023 |
* |
-0,00015 |
3,4 |
0,34 |
0,9 |
8,5 |
|
1,08-10"5 |
* |
0,02165 |
-0,00014 |
3,4 |
0,35 |
0,87 |
11,9 |
|
|
1,12-Ю"5 |
0,008402 |
0,01566 |
-0,00015 |
3,6 |
0,34 |
0,89 |
9,6 |
|
|
7,ЗЛО"5 |
0,01068** |
-0,00014 |
3,2 |
0,37 |
0,9 |
9,3 |
||
|
Изюбрь |
0,02951 |
0,00988 |
* |
-0,0002 |
1,7 |
0,58 |
0,48 |
15,6 |
|
0,01944 |
* |
0,007822 |
-0,00022 |
1,8 |
0,55 |
0,39 |
16,3 |
|
|
0,50241 |
0,02996 |
-0,01941 |
-0,00015 |
1,5 |
0,66 |
0,61 |
13,4 |
|
|
0,02124 |
0,00446** |
-0,00021 |
1,8 |
0,56 |
0,43 |
16,1 |
||
|
Кабан |
0,00779 |
0,01367 |
* |
-0,0003 |
2,3 |
0,45 |
0,69 |
13,5 |
|
0,00295 |
* |
0,01123 |
-0,00026 |
2,1 |
0,5 |
0,63 |
14 |
|
|
0,01022 |
0,01601 |
-0,000303 |
-0,0003 |
2,3 |
0,44 |
0,69 |
13,3 |
|
|
0,00406 |
0,00629** |
-0,00028 |
2,2 |
0,47 |
0,66 |
13,8 |
||
|
Соболь |
2,496-10"7 |
0,039 |
* |
-0,0002 |
2,7 |
0,52 |
0,85 |
23,8 |
|
2,ЮЗ-10"8 |
* |
0,03192 |
-0,00019 |
2,5 |
0,54 |
0,85 |
22,9 |
|
|
5,68-10"7 |
0,0424 |
-0,00386 |
-0,0002 |
2,6 |
0,52 |
0,85 |
23,8 |
|
|
1,5829-10"8 |
0,01902** |
-0,00021 |
2,8 |
0,51 |
0,84 |
22,4 |
||
|
Рысь |
2,43-10"4 |
0,02121 |
* |
-0,00986 |
1,6 |
0,64 |
0,28*** |
23,3 |
|
2,03-10"6 |
* |
0,023729 |
-0,01459 |
2 |
0,53 |
0,47 |
23,9 |
|
|
0,000103 |
0,017159 |
0,004486 |
-0,0113 |
1,7 |
0,63 |
0,53 |
23,6 |
|
|
1,4-10"5 |
0,01185** |
-0,01327 |
1,9 |
0,59 |
0,55 |
24,1 |
||
|
Медведь бурый |
0,33767 |
0,005433 |
* |
-0,00183 |
3,2 |
0,32 |
0,54 |
4,7 |
|
0,23152 |
* |
0,00427 |
-0,00159 |
2,8 |
0,36 |
0,52 |
4,6 |
|
|
0,33584 |
0,00538 |
0,000045 |
-0,00182 |
3,2 |
0,32 |
0,54 |
4,6 |
|
|
0,26222 |
0,002444** |
-0,00171 |
3 |
0,34 |
0,54 |
4,5 |
||
* Данный фактор не оценивался в модели.
** Учитывалась общая площадь лиственных пород.
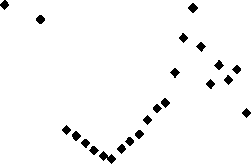
*** Согласно ^-критерию Фишера R2 является статистически незначимым намика объема вырубленной ликвидной древесины при рубках леса главного пользования за период 19802013 гг. продемонстрирована на рис. 2.
Проведение рубок леса в Еврейской автономной области (ЕАО) в 70-80 гг. XX столетия на больших площадях и в большом объеме привело к сокращению лесопокрытых площадей и смене пород, а также значительному сокращению наиболее ценных кедровых лесов. В мае 1994 г. кедр корейский занесен в Красную книгу ЕАО и его рубка запрещена. Однако на площадях, пройденных рубкой, в настоящее время до сих пор произрастают вторичные леса с преобладанием лиственных пород [2]. Как видно из рис. 2, в 90-е гг. в ЕАО наблюдался спад активности лесозаготовки, а к концу 2000-х гг. наметилась тенденция роста объемов заготовок древесины. В последние 6 лет объем изъятия леса имеет тенденцию к снижению, и в настоящее время объемы лесозаготовок незначительны.
Зарастающие рубки леса являются местами обитания многих животных. На вырубках охотничьего типа обитают те же виды животных, что и в елово-пихтовых или лиственничных лесах, на месте которых они возникли. К ним относятся лось, изюбрь, косуля, соболь,
. 400 1
«о
* 300 -
1975 1980 1985 1990 1995 2000 2005 2010 2015
♦ Заготовка древесины
Рис. 2. Динамика объема вырубленной ликвидной древесины при рубках леса главного пользования колонок [8]. Вырубкам маньчжурского типа в основном свойственна фауна кедрово-широколиственных лесов (лось, изюбрь, косуля, кабан, соболь, колонок и лисица) [8].
Согласно результатам корреляционного анализа (табл. 3), между оценками численности животных и рубками леса главного пользования выявлено, что измене-
Таблица 3
|
Показатели антропогенных факторов |
Вид ЖИВОТНОГО |
Коэффициент кор |
реляции гь где к - номер лага (уровень значимости а) |
||||||||
|
И |
Гг |
Гз |
Г5 |
Г6 |
г- |
rs |
г» |
По |
|||
|
Площадь рубки леса главного пользования |
Косуля |
0,74 (0,01) |
0,8025 (0,01) |
0,9021 (0,01) |
0,8349 (0,01) |
0,8309 (0,01) |
0,7469 (0,01) |
0,7012 (0,01) |
0,6618 (0,05) |
0,6337 (0,05) |
0,5063 (*) |
|
Кабан |
0,2972 |
0,5298 (0,05) |
0,6183 (0,05) |
0,6614 (0,01) |
0,7093 (0,01) |
0,7389 (0,01) |
0,8006 (0,01) |
0,8544 (0,01) |
0,9161 (0,01) |
0,8193 (0,01) |
|
|
Изюбрь |
-0,39 (*) |
-0,256 (*) |
-0,063 (*) |
0,0696 (*) |
0,2296 (*) |
0,5413 (0,05) |
0,6948 (0,01) |
0,7001 (0,05) |
0,4729 (*) |
0,4387 (*) |
|
|
Медведь бурый |
0,56 (0,05) |
0,721 (0,01) |
0,766 (0,01) |
0,58 (0,05) |
0,583 (0,05) |
0,486 п |
0,457 И |
0,434 И |
0,403 п |
0,407 И |
|
|
Рысь |
-0,4 (*) |
-0,068 (*) |
0,22 (*) |
0,51 (*) |
0,661 (0,05) |
0,8531 (0,01) |
0,8541 (0,01) |
0,8137 (0,01) |
0,7776 (0,05) |
0,7469 (0,05) |
|
|
Лисица |
-0,005 |
0,0365 И |
0,364 п |
0,5473 (0,05) |
0,5208 (0,05) |
0,294 п |
0,2095 И |
0,2421 И |
0,5055 п |
0,5987 И |
|
|
Соболь |
0,7094 (0,01) |
0,7917 (0,01) |
0,6314 (0,05) |
0,7458 (0,01) |
0,8445 (0,01) |
0,9642 (0,01) |
0,9406 (0,01) |
0,9236 (0,01) |
0,9202 (0,01) |
0,8635 (0,01) |
|
|
Зайцы |
0,6621 (0,01) |
0,6843 (0,01) |
0,6273 (0,05) |
0,4425 (*) |
0,4299 (*) |
0,2202 (*) |
0,1812 (*) |
0,0771 (*) |
-0,116 (*) |
-0,356 (*) |
|
|
Объем вырубленной ликвидной древесины при рубках леса главного пользования |
Косуля |
0,7145 (0,01) |
0,8074 (0,01) |
0,8911 (0,01) |
0,8786 (0,01) |
0,8353 (0,01) |
0,7583 (0,01) |
0,6517 (0,01) |
0,5892 (0,05) |
0,4761 п |
0,2911 И |
|
Кабан |
0,2972 (*) |
0,5298 (0,05) |
0,6183 (0,01) |
0,6614 (0,01) |
0,7093 (0,01) |
0,7389 (0,01) |
0,8006 (0,01) |
0,8544 (0,01) |
0,8709 (0,01) |
0,7512 (0,01) |
|
|
Изюбрь |
-0,376 |
-0,258 И |
-0,12 |
0,0854 (*) |
0,2443 И |
0,3678 п |
0,4528 И |
0,6052 (0,05) |
0,5316 п |
0,3494 И |
|
|
Медведь бурый |
0,073 |
0,616 (0,01) |
0,7312 (0,01) |
0,6665 (0,01) |
0,5363 (0,05) |
0,4529 п |
0,4621 И |
0,438 И |
0,341 п |
0,2569 И |
|
|
Рысь |
-0,34 |
-0,21 И |
-0,087 |
0,1061 (*) |
0,3492 И |
0,794 (0,01) |
0,7723 (0,01) |
0,744 (0,01) |
0,7865 (0,01) |
0,7535 (0,01) |
|
|
Лисица |
0,2812 п |
0,2287 И |
0,3387 п |
0,4993 (0,05) |
0,5448 (0,05) |
0,4173 п |
0,2665 И |
0,2824 И |
0,382 п |
0,4973 И |
|
|
Соболь |
0,4147 п |
0,6214 (0,01) |
0,6585 (0,01) |
0,7427 (0,01) |
0,8568 (0,01) |
0,95 (0,01) |
0,9278 (0,01) |
0,879 (0,01) |
0,8192 (0,01) |
0,722 (0,05) |
|
|
Зайцы |
0,7316 (0,01) |
0,6382 (0,01) |
0,6386 (0,01) |
0,461 (*) |
0,4227 И |
0,3245 |
0,163 И |
0,0988 И |
-0,082 |
-0,173 |
|
(*) - незначимый коэффициент корреляции
Оценки параметров модели (1) с учетом изменения объема вырубленной ликвидной древесины
|
Вид ЖИВОТНОГО |
Коэффициенты модели (1) |
Яср |
Vcp |
R2 |
А, % |
||
|
«0 |
W |
Ьо |
|||||
|
Косуля |
1,72 |
0,00206 |
-0,0001 |
2,33 |
0,47 |
0,78 |
12,4 |
|
Изюбрь |
1,71 |
0,00129 |
-0,00024 |
1,95 |
0,51 |
0,36 |
15,6 |
|
Кабан |
2,7 |
0,00394 |
-0,00044 |
3,5 |
0,29 |
0,72 |
8,7 |
|
Рысь |
1,41 |
0,00214 |
-0,0139 |
1,9 |
0,55 |
0,65 |
25,2 |
|
Медведь |
2,77 |
0,00080 |
-0,00179 |
3,1 |
0,32 |
0,57 |
5,4 |
|
Лисица |
1,65 |
0,00117 |
-0,00071 |
1,93 |
0,55 |
0,3 |
22,2 |
|
Соболь |
1,27 |
0,00456 |
-0,00021 |
2,6 |
0,53 |
0,85 |
23,1 |
|
Зайцы |
2,37 |
0,00331 |
-0,0003 |
4,1 |
0,28 |
0,28 |
25,4 |
Результаты парной корреляции между численностью диких животных в и-год и показателями рубки леса в (и-^)-год (к - лаг)
ния численности рассматриваемых популяций хорошо коррелируют с изменениями площадей, на которых осуществляются рубки леса главного пользования, и объемом вырубленной ликвидной древесины при данных рубках в предыдущие годы (табл. 3).
Из табл. 3 видно, что увеличение площадей и объема рубок леса способствует увеличению численности косули, соболя и зайцев на следующий год, кабана - на второй год, лисицы - на четвертый, рыси - на пятый, изюбря - на шестой год после рубок. Кроме этого, тесная положительная корреляционная связь между рубками леса и численностью животных прослеживается довольно длительный период. Для популяции косули эта связь наблюдается в течение первых 9 лет после рубок леса, для кабана, соболя - со 2 по 10 год, для изюбря - с 6 по 8 год, для медведя - со 2 по 5 год, для рыси - с 5 по 10 год, для лисицы - с 4 по 5 год, зайцев - с 1 по 3 год. По-видимому, колебания численности видов связаны с сукцессиями леса, вызванными рубкой леса главного пользования. Рубки леса (особенно выборочные) ведут к осветлению и буйному разрастанию подлеска, подроста и трав, сохранившиеся деревья и ягодные кустарники лучше плодоносят, куртины деревьев чередуются с полями. Все это улучшает условия для обитания косули, зайцев, изюбра и лося [8]. По мере развития разнообразной светолюбивой травянистой растительности лесосеки заселяют мышевидные грызуны, привлекающие не только лисицу, но и соболя. Такие виды, как бурый медведь и рысь, требуют наличия значительных лесных массивов, но даже для них смешанные леса, состоящие из разнообразных пород и возрастов и их сочетаний, являются благоприятным типом угодий [24]. Более того, на лесосеках может поселиться рысь, поскольку здесь возрастают запасы ее основной пищи - грызуны, косули и зайцы, а медведя привлекают разрастающиеся на вырубках ягодники [5]. Сроки зарастания лесосек довольно изменчивы, иногда лесная поросль может не появляться несколько лет. Поэтому в процессе возрастно-породной сукцессии лесной растительности периоды наибольшей благоприятности для животных варьируются [24]. Вместе с тем, наиболее продуктивными для животных являются молодняки, то есть насаждения в возрасте до 20 лет - для хвойных и твердолиственных пород и до 10 лет - для мягколиственных [23]. Молодняки сосны, осины, березы, дуба являются важнейшими зимними кормовыми стациями копытных и зайцев [5]. Но по мере увеличения высоты деревьев доступ ность и биомасса кормов сокращаются, а соответственно сокращается и поголовье копытных.
Корреляционный анализ динамики численности копытных в зависимости от интенсивности лесозаготовок был дополнен модельным анализом, при котором использовалась модифицированная модель Рикера (1) с учетом объема вырубленной ликвидной древесины при рубках леса главного пользования (табл. 4).
Отметим, что лаг (запаздывание) в модели определялся в зависимости от наибольшего значения коэффициента корреляции между рубками леса и численностью животных (табл. 3). Полученные оценки параметров модели (1) приведены в табл. 4. Сравнительно высокие значения коэффициентов детерминации (табл. 4), полученные между оценками модельных и учетных численностей косули, кабана и соболя, позволяют судить о немаловажном влиянии динамики лесозаготовок на флуктуации их численности. Вместе с тем, отклонения расчетных оценок от учетных данных, скорее всего, связаны с неучтенными в модели показателями.
Воздействие изменения посевных площадей на динамику численности охотничьих животных
Активное освоение земель под сельскохозяйственные культуры улучшает условия обитания многих диких животных. Лучшими культурными растениями для диких копытных являются такие растения, которые содержат максимум белка (например, рапс, соя, овес, озимая рожь, кормовые бобы, кормовая капуста, свекла, гречиха, горох, люцерна, эспарцет, вика). Привлекают также копытных (особенно кабанов) картофельные и кукурузные поля [4]. Хищных зверей (например, лисицу и колонка) на сельскохозяйственных полях привлекают обильные запасы мышевидных грызунов, являющихся для них основным кормом [17].
По результатам корреляционного анализа установлено, что расширение посевных площадей кукурузы на зерно способствует росту численности популяции кабана, сои - кабана и косули, кормовых культур, кукурузы на корм и многолетних трав - изюбря, однолетних трав - кабана, косули и лисицы, кормовых бахчевых -косули, кабана и лисицы, зерновых культур и картофеля - колонка (табл. 5).
Наиболее тесная корреляционная связь была получена между численностью косули и посевной площадью сои, между численностью кабана и посевами сои, а также между численностью изюбря и посевами многолетних трав. Согласно литературным источникам, на-
Результаты парной корреляции между посевными площадями сельскохозяйственных культур в (и-1)-году и численностью диких копытных в и-году
|
Культуры \ Виды |
Изюбрь |
Косуля |
Кабан |
Колонок |
Лисица |
|
Зерновые культуры |
* |
* |
* |
0,5648 (0,05) |
* |
|
Кукуруза на зерно |
* |
* |
0,4602 (0,05) |
* |
* |
|
Соя |
* |
0,8862 (0,01) |
0,7784 (0,01) |
* |
* |
|
Картофель |
* |
* |
* |
0,6445 (0,05) |
* |
|
Кормовые культуры |
0,6534 (0,01) |
* |
* |
* |
* |
|
Кукуруза на корм |
0,6341 (0,01) |
* |
* |
* |
* |
|
Кормовые бахчевые |
* |
0,5646 (0,05) |
0,565 (0,05) |
* |
0,6 (0,05) |
|
Многолетние травы |
0,6823 (0,01) |
* |
* |
* |
* |
|
Однолетние травы |
* |
0,6838 (0,01) |
0,5615 (0,05) |
* |
0,5 (0,05) |
(*) - незначимый коэффициент корреляции пример [5, 17], в сельскохозяйственных угодьях косуля может обитать постоянно, но численность ее меняется по сезонам и годам в зависимости от наличия соевого жнивья, состояния снежного покрова и преследования человеком. Кабан и изюбрь заходят в сельскохозяйственные поля преимущественно осенью, хотя встречаются здесь и в конце лета, и зимой в случае неурожая основных видов кормов, при этом восполняя дефицит естественных кормов за счет остатков урожая на полях. Из-за преследования человеком и отсутствия убежищ эти животные здесь находятся недолго [17].
В табл. 6 представлены результаты верификации модели (1) с учетом посевных площадей сельскохозяйственных культур.
В соответствие со значениями коэффициентов детерминации и аппроксимации (табл. 6), модель (1) с учетом посевных площадей сои наилучшим образом описывает динамику численности косули и кабана. Несколько хуже модель (1), учитывающая площади кормовых культур, описывает динамику численности изюбря и лисицы. Скорее всего, это связано с незначительной ролью расширения посевных площадей кормовых сельскохозяйственных культур в развитии этих популяций, а также не включенными в модель (1) другими факторами. Согласно оценкам коэффициентов детерминации модель (1) с учетом изменения посевных площадей зерновых культур и картофеля весьма слабо описывает динамику численности популяции колонка, по-видимому, для описания колебаний его численности необходимо учитывать другие факторы.
Комплексное влияние изменения площадей лиственных пород, объема вырубки леса и посевных площадей сельскохозяйственных культур на флуктуации численности охотничьих животных
Как правило, на популяцию действует совокупность факторов внешней среды, поэтому целесообразно анализировать динамику численности животных по нескольким факторам. В табл. 7 представлены оценки параметров модели динамики численности косули, изюбря, кабана, лисицы, медведя бурого, рыси, соболя с учетом различных комбинаций таких показателей, как площадь лиственных пород, объем вырубки и посевная площадь сельскохозяйственных культур.
На рис. 3 представлены фактическая и модельная численности косули (а), соболя (б) и кабана (в) с учетом площади лиственных пород, площади соевых полей и объема вырубки 3-летней давности.
Из табл. 7 видно, что наилучшим образом модель Рикера с учетом комбинаций внешних факторов описывает динамику численности косули (коэффициент аппроксимации варьируется в пределах 8,3-9,8%, а коэффициент детерминации - от 0,86 до 0,9). Как видно из рис. За, модель (2) хорошо улавливает общие тенденции динамики численности косули и описывает основные всплески наблюдаемой численности. Следовательно, можно предполагать, что указанные факторы оказывают наибольшее влияние на динамику численности косули. Действительно, согласно литературным источникам (например, [5, 8]), лучшими местами обитания косули являются леспедециево-лещинные дубняки, разно-
Таблица 6
Оценки параметров модели (1) с учетом изменения посевных площадей сельскохозяйственных культур
|
Вид животного |
Коэффициенты модели (1) |
Яср |
Vcp |
R2 |
А, % |
||
|
«0 |
С |
Ьо |
|||||
|
Косуля (соя) |
1,53 |
0,012624 |
-0,000129 |
2,8 |
0,4 |
0,77 |
10,8 |
|
Изюбрь (кормовые) |
1,63 |
0,015022 |
-0,00021 |
1,9 |
0,54 |
0,43 |
15,8 |
|
Изюбрь (кукуруза) |
1,72 |
0,01909 |
-0,000204 |
1,9 |
0,54 |
0,38 |
17,2 |
|
Изюбрь (травы) |
1,72 |
0,081245 |
-0,000251 |
2,2 |
0,47 |
0,45 |
16,5 |
|
Кабан (соя) |
1,51 |
0,007038 |
-0,00027 |
2,1 |
0,49 |
0,62 |
13,4 |
|
Колонок (зерновые) |
1,19 |
0,01595 |
-0,00018 |
1,9 |
0,54 |
0,15* |
24 |
|
Колонок (картофель) |
0,66 |
0,133777 |
-0,000171 |
1,79 |
0,56 |
0,11* |
23,4 |
|
Лисица (бахчевые) |
1,24 |
1,523541 |
-0,000554 |
1,65 |
0,64 |
0,48 |
23 |
|
Лисица (травы) |
1,37 |
0,05018 |
-0,000517 |
1,59 |
0,66 |
0,53 |
21,4 |
Согласно ^-критерию Фишера R2 является статистически незначимым
Таблица 7
Оценки параметров модели (2) с учетом изменения площадей лиственных пород, объема вырубки, посевных площадей сельскохозяйственных культур
|
Вид |
Коэффициенты модели (2) |
дср |
vcp |
id |
Л, % |
||||
|
«0 |
d |
W |
С |
Ьо |
|||||
|
Косуля |
1,73 |
* |
0,001366 |
0,009546 |
-0,00015 |
3,4 |
0,35 |
0,86 |
8,3 |
|
9,7-10"6 |
0,01281 |
* |
-0,00033 |
-0,00015 |
3,6 |
0,34 |
0,89 |
9,8 |
|
|
5,3-10"5 |
0,01102 |
6,9?10"5 |
* |
-0,000144 |
3,3 |
0,36 |
0,9 |
9,4 |
|
|
1,5-10"4 |
0,00991 |
4710"13 |
0,002989 |
-0,0001525 |
3,3 |
0,34 |
0,9 |
8,9 |
|
|
Изюбрь |
1,71 |
* |
0,00129 |
-0,000527 |
-0,000243 |
1,9 |
0,52 |
0,36 |
15,6 |
|
0,0196252 |
0,00456 |
* |
-0,005216 |
-0,000204 |
1,7 |
0,57 |
0,43 |
16,3 |
|
|
0,00178 |
0,00697 |
-0,00105 |
* |
-0,000177 |
1,6 |
0,62 |
0,46 |
16 |
|
|
0,0015368 |
0,00714 |
-0,00107 |
-0,006083 |
-0,000168 |
1,6 |
0,63 |
0,47 |
16,4 |
|
|
Кабан |
1,47 |
* |
0,000093 |
0,007197 |
-0,000276 |
2,2 |
0,48 |
0,65 |
13,8 |
|
0,015665 |
0,00486 |
* |
0,002359 |
-0,000295 |
2,3 |
0,45 |
0,68 |
13,5 |
|
|
0,000865 |
0,00789 |
-0,00056 |
* |
-0,000271 |
2,1 |
0,48 |
0,66 |
13,9 |
|
|
0,003522 |
0,00638 |
-0,00065 |
0,002897 |
-0,000286 |
2,2 |
0,47 |
0,68 |
13,5 |
|
|
Лисица |
1,5 |
* |
0,00009 |
0,061742 |
-0,000641 |
1,8 |
0,58 |
0,45 |
19,6 |
|
Медведь |
1,04 |
0,001 |
0,00045 |
* |
-0,00173 |
3 |
0,34 |
0,6 |
4 |
|
Рысь |
2,63-10"6 |
0,0136 |
-0,00059 |
* |
-0,01291 |
1,8 |
0,6 |
0,59 |
23,9 |
|
Соболь |
1,7-10"5 |
0,01162 |
0,001945 |
* |
-0,00018 |
2,5 |
0,55 |
0,87 |
21,3 |
- Данный фактор не оценивался в модели
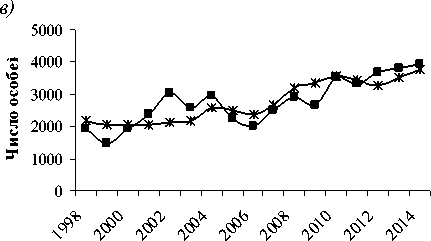
—Ж—Модельная численность —■—Учетная численность
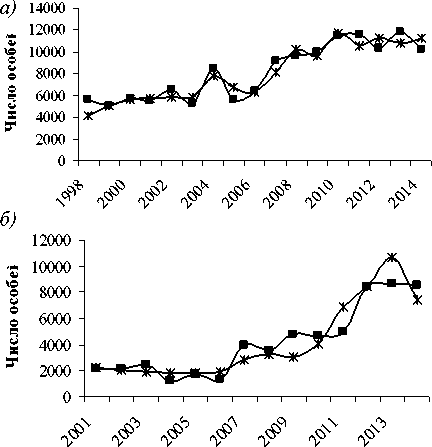
Рис. 3. Результаты моделирования и данные учетной численности популяций:
а) косули (факторы: площади лиственных пород и соевых полей, объем вырубки 3-летней давности), б) соболя (факторы: площади лиственных пород 4 года назад и объем вырубки 6летней давности), в) кабана (факторы: площади лиственных пород и соевых полей, объем вырубки 4-х летней давности)
травно-кустарниковые лесистые редки в окружении травяных лугов, небольшие леса (колки) вокруг сельскохозяйственных полей, молодняки (осиново-березовое возобновление) на гарях и лесосеках. Завышенный коэффициент пополнения популяции (3,3-3,6), характеризует, скорее всего, кроме естественного прироста, также и миграционную активность зверей. По-видимому, на территории автономии сложились благоприятные условия для обитания косули, что привлекает особей из сопредельных области территорий. Для других видов животных модель Рикера (2) несколько хуже описывает динамику их численности (табл. 7). Скорее всего, это связано с неучтенными в модели показателями, которые оказывают наиболее существенное влияние на динамику численности животных, чем рассмотренные факторы.
Сравнение коэффициентов, характеризующих интенсивности воздействия рассмотренных факторов (табл. 7), выявило, что на динамику численности популяции косули и кабана в первую очередь оказывают воздействие изменения площади лиственных пород, затем вариации посевных площадей сои, а уже потом объем лесозаготовок. Наибольшее влияние на колебания численности изюбрей - динамика площади лиственных пород, затем вырубки леса и наименьшее воздействие оказывает изменение посевных площадей кормовых культур. На динамику численности лисицы в большей степени воздействует расширение посевных площадей однолетних трав, а на колебания численности медведя, рыси, соболя в большей степени воздействует изменение площади лиственных насаждений.
Заключение
Таким образом, в статье показано, что увеличение площадей лиственных лесов в ЕАО, происходящее вследствие совокупной разнородной хозяйственной деятельности человека, создает благоприятные условия для охотничьих животных (косули, изюбря, кабана, бурого медведя, рыси и соболя) и способствует росту их
численности. Увеличение площади хвойных лесов в период 1997-2003 гг. также благоприятно повлияло на развитие популяционной динамики рассмотренных видов (кроме изюбря). Вместе с тем, с 2004 г. по настоящее время отмечается негативная тенденция сокращения площади хвойных лесов, которая может привести к уменьшению численности животных в среднесрочной и долгосрочной перспективе. Тем не менее, в настоящее время для них в пределах автономии так сложились условия среды обитания, что уменьшение площади хвойных насаждений пока что не препятствует росту их численности.
Анализ отдельных видов хозяйственной деятельности человека (рубок леса главного пользования и эксплуатации угодий сельскохозяйственными предприятиями) показал, что рассмотренные направления деятельности в целом улучшают условия обитания животных. Выявлено, что по мере сукцессии лесной растительности после рубок леса главного пользования численность копытных, рыси, соболя, лисицы и зайцев возрастает. Установлено, что увеличение посевных площадей сои, зерновых и кормовых культур способствует росту численности копытных (изюбря, кабана, косули), лисицы и колонка.
В результате модельного анализа установлено, что из рассмотренных показателей наибольшее влияние на колебания численностей косули, изюбря, кабана, медведя, рыси, соболя оказывает динамика площади лиственных пород, а на динамику численности лисицы -расширение посевных площадей сельскохозяйственных культур. Второстепенное значение на динамику численности косули и кабана оказывает изменение посевных площадей сои, а на колебания численности изюбря, медведя, рыси, соболя и лисицы - рубки леса. Наименьшее воздействие на флуктуации численности косули и кабана оказывает изменение вырубок леса, а на изменение численности изюбря - расширение посевных площадей кормовых культур.
Показано, что наилучшим образом модифицированный вариант модели Рикера с учетом изменения площадей лесных насаждений, посевных площадей сельскохозяйственных культур и объема рубок леса главного пользования описывает динамику численности популяции косули. По-видимому, рассмотренные факторы оказывают наибольшее влияние на кормовые и защитные условия среды обитания косули и, следовательно, на динамику ее численности.
Исследование выполнено при финансовой поддержке Комплексной программы фундаментальных исследований «Дальний восток».
Список литературы О возможности использования эффекта кластеризации в системах связанных отображений для описания динамики метапопуляций
- Иванова А.С., Кузнецов С.П. Волны кластеризации в цепочке систем, каждая из которых содержит набор элементов с внутренней глобальной связью//Изв. вузов «ПНД». 2003. т. 11, № 4-5. С. 80-88.
- Кузнецов С.П. Универсальность и подобие связанных систем Фейгенбаума//Изв. Вузов: Радиофизика. 1985. Т. 27, № 8. с. 991-1007.
- Кулаков М.П., Аксенович Т.И., Фрисман Е.Я. Подходы к описанию пространственной динамики миграционно-связанных популяций: анализ синхронизации циклов//Региональные проблемы. 2013. Т. 16, № 1. с. 5-15.
- Кулаков М.П., Ревуцкая О.Л. Применение метапопуляционного подхода к анализу пространственно-временной динамики промысловых животных//Фундаментальные и прикладные исследования в математической экологии и агроэкологии. Материалы международной школы-семинара. 2012. с. 94-98.
- Кулаков М.П., Ревуцкая О.Л. Применение метапопуляционного подхода к анализу пространственно-временной динамики промысловых животных (на примере популяций кабана и изюбря)//Региональные проблемы. 2011. Т. 14, № 2. с. 12-20.
- Кулаков М.П., Фрисман Е.Я. Синхронизация 2-циклов в системе симметрично связанных популяций, запас-пополнение в которых описывается функцией Рикера//Изв. вузов «ПНД». 2010. Т. 18, № 6. с. 25-41.
- Таборов А.В. Частичная хаотическая синхронизация в системах связанных логистичеких отображений//Динам. системы. 2001. Вып. 17. с. 29-35.
- Ecology, Genetics and Evolution of Metapopulations/Edited by Hanski Ilkka, Gaggiotti Oscar, London: Academic Press. 2004. 696 p.
- Kaneko K. Clustering, coding, switching, hierarchical, ordering, and control in network of chaotic elements, Physica D. 1990. vol. 41. p. 137-172.
- Kaneko, K. Relevance of dynamic clustering to biological network, Phisica D. 1994. vol. 75. p. 55-73.
- Kulakov M., Revutskaya O., Frisman E. Application of a metapopulation approach for analysis of space-time population dynamics (case study using ungulates)//iEMSs 2012 -Managing Resources of a Limited Planet: Proceedings of the 6th Biennial Meeting of the International Environmental Modelling and Software Society. 2012. p. 250-255.
- Omelchenko I., Maistrenko Yr., Mosekilde E. Synhronization in ensembles of coupled maps with a major element//Discrete Dynamics in Nature and Society. 2005. no. 3. p. 239-255.
- Opdam P. Metapopulation theory and habitat fragmentation: a review of holarctic breeding bird studies//Landscape Ecology. 1991. vol. 5, no. 2. p. 93-106.
- Oppo G.-L., Kapral R. Discrete models for the formation and evolution of spatial structure in dissipative systems//Phys. Rev. A. 1984. vol. 33, no. 6. p. 4219-4231.
- Popovych O., Pikovsky A., Maistrenko Yu. Cluster-splitting bifurcation in a system of coupled maps//Physica D. 2002. vol. 168-169. p. 106-125.
- Udwadia F.E., Raju N. Dynamics of Coupled Nonlinear Maps and Its Application to Ecological Modeling//Applied mathematic and computation.1997. vol. 82. p. 137-179.
- Vanderlei Manica, Jacques A.L. Silva. Population distribution and synchronized dynamics in a metapopulation model in two geographic scales//Mathematical Biosciences. 2014. vol. 250. p. 1-9.
- Waller I., Kapral R. Spatial and temporal structure in systems of coupled nonlinear oscillators//Phys. Rev. A. 1986. vol. 30, no. 4. p. 2047-2055.
- Wysham D.B., Hastings A. Sudden Shift Ecological Systems: Intermittency and Transients in the Coupled Riker Population Model//Bulletin of Mathematical Biology. 2008. vol. 70. p. 1013-1031.
