О зависимости адаптации гибридов овса F1 к повышенному содержанию ионов алюминия в культуральной среде от компонентов скрещивания

Автор: Лисицын Е.М., Баталова Г.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Экологические проблемы растениеводства
Статья в выпуске: 3 т.40, 2005 года.
Бесплатный доступ
В лабораторных и полевых опытах определяли физиолого-морфологические параметры алюмоустойчивости у гибридов овса F1 в различных комбинациях скрещивания. Оценивали характер взаимодействия генов, контролирующих развитие растений.
Короткий адрес: https://sciup.org/142133081
IDR: 142133081 | УДК: 633.13:631.527.5
About influence of parent components of cross on adaptation of F1 oat hybrids against heightened contents of aluminum ions in cultural media
Influence of parent components of cross on a development of different physiological and morphological parameters of aluminum resistance of oat F1 hybrids was studied under laboratory and field conditions. The character of genes interrelations determining the plants development was estimated. The significant influence of a direction of cross both on development of the studied parameters, and on nature of a gene interaction encoding their determinants was established.
Текст научной статьи О зависимости адаптации гибридов овса F1 к повышенному содержанию ионов алюминия в культуральной среде от компонентов скрещивания
Эколого-генетический подход к селекции растений позволяет разрабатывать методы и направления рационального использования уже созданных сортов, отбирать новые генотипы, устойчивые к лимитирующим факторам среды, получать информацию о норме реакции последних, а также выявлять причины изменчивости генетических признаков и физиологических свойств растений (1, 2). Однако отсутствие к настоящему времени частной генетики отдельных сельскохозяйственных культур значительно затрудняет задачу правильного подбора родительских компонентов скрещивания для получения высокоустойчивых к стрессовым факторам среды сортов с высокой потенциальной продуктивностью.
Целью нашей работы была оценка влияния родительских компонентов скрещивания на проявление в гибридном потомстве устойчивости к такому стрессовому эдафическому фактору, как повышенное содержание подвижных ионов алюминия в культуральной среде.
Методика . Материалом исследования служили сорта овса ( Avena sativa ) Ar-gamak, Freija и Petra, а также гибриды F1, полученные посредством прямых и обратных скрещиваний этих сортов. Растения выращивали в условиях рулонной культуры по ранее описанной методике (3, 4). Относительную алюмоустойчивость определяли у 5-суточных проростков: I группа (контроль) — рН 6,0, без ионов Al; II группа — рН 4,3, 1 мМ Аl 2 (SO 4 ) 3 . Анализировали следующие показатели: ИДК (соотношение длины наибольшего корня в опыте и контроле); катионообменную емкость корней (5); окислительную активность корней (6); активность кислой фосфатазы (КФ 3.1.3.1.) и кислой липазы (КФ 3.1.1.3.) (7). Долю влияния родительских компонентов рассчитывали методом двухфакторного дисперсионного анализа.
Для оценки генетических систем адаптивности растений родительских компонентов и гибридов овса F1 в полевых условиях использовали графическую методику (1). Растения выращивали в соответствии с методикой государственного сортоиспытания (8): повторность 4-кратная; делянки размещали в один ярус; расположение вариантов внутри повторений рендомизированное; общая площадь делянок — 50 м2, учетных площадок — 10 м2.
Результаты . В таблице представлены физиолого-биохимические показатели растений овса, использованные для характеристики устойчивости зерновых культур к токсичному действию ионов алюминия. Доля влияния обеих родительских форм на длину корня у гибридов F 1 в I группе была практически одинаковой: материнской, отцовской форм и их взаимодействия — соответственно 19,4; 17,0 и 58,9 %. Под действием стрессового фактора влияние родительских форм усиливалось (соответственно 44,1 и 28,8 %) за счет снижения доли влияния их взаимодействия (17,7 %). При этом на показатель ИДК отцовский компонент не оказывал влияния, доля же влияния материнской формы возросла до 51,1 %, а взаимодействия компонентов — до 30,2 %.
Физиолого-биохимические показатели гибридов овса F1 в различных комбинациях скрещивания под влиянием ионов алюминия (рулонная культура)
Гибрид I группа (контроль) II группа % от контроля
Д л и н а к о р н я, мм
|
Petra x Freija |
118,8 ± 3,5 |
93,6 ± 4,0 |
78,8 ± 3,4 |
|
Freija x Petra |
114,8 ± 4,6 |
101,6 ± 1,8 |
88,5 ± 1,6 |
|
Petra x Argamak |
109,0 ± 2,6 |
85,7 ± 2,7 |
78,6 ± 2,5 |
|
Argamak x Petra |
110,1 ± 2,8 |
81,4 ± 6,6 |
73,9 ± 6,0 |
|
Argamak x Freija |
78,4 ± 1,6 |
66,7 ± 1,2 |
85,1 ± 1,6 |
|
Freija x Argamak |
76,0 ± 2,0 А к т и в н о с т ь к и с л о й л и п а з ы, |
72,0 ± 1,7 мл 0,1 н. NaOH/г сухой массы |
94,8 ± 2,3 |
|
Petra x Freija |
2,23 ± 0,11 |
1,80 ± 0,19 |
80,81 ± 5,02 |
|
Freija x Petra |
2,31 ± 0,06 |
1,16 ± 0,07 |
50,05 ± 1,75 |
|
Argamak x Petra |
1,93 ± 0,13 |
1,40 ± 0,02 |
72,21 ± 0,49 |
|
Petra x Argamak |
1,87 ± 0,09 |
1,38 ± 0,15 |
73,72 ± 4,67 |
|
Argamak x Freija |
2,92 ± 0,22 |
1,48 ± 0,06 |
50,47 ± 1,18 |
|
Freija x Argamak |
2,69 ± 0,09 |
1,15 ± 0,08 |
42,69 ± 1,62 |
|
О |
к и с л и т е л ь н а я а к т и в н о с т ь к о |
р н е й, мг KMnO 4 / г сухой массы |
|
|
Petra x Freija |
5,38 ± 0,51 |
6,00 ± 0,55 |
111,54 ± 10,18 |
|
Freija x Petra |
10,81 ± 0,74 |
10,91 ± 0,33 |
100,96 ± 3,08 |
|
Argamak x Petra |
6,79 ± 0,16 |
6,16 ± 0,23 |
90,79 ± 3,41 |
|
Petra x Argamak |
7,39 ± 0,06 |
10,73 ± 0,45 |
146,17 ± 6,16 |
|
Argamak x Freija |
7,87 ± 0,34 |
9,09 ± 0,10 |
115,46 ± 1,25 |
|
Freija x Argamak |
4,16 ± 0,37 |
6,62 ± 0,54 |
159,21 ± 13,01 |
|
К а т и о н о о б м е н н а я е м к о с т ь к |
о р н е й, мг-экв/г сухой массы |
||
|
Petra x Freija |
62,5 ± 7,7 |
61,1 ± 4,2 |
97,8 ± 6,7 |
|
Freija x Petra |
64,9 ± 3,3 |
31,5 ± 0,6 |
48,5 ± 0,9 |
|
Argamak x Petra |
53,4 ± 1,0 |
65,8 ± 1,1 |
123,2 ± 2,1 |
|
Petra x Argamak |
94,8 ± 1,1 |
90,3 ± 1,1 |
95,3 ± 1,2 |
|
Argamak x Freija |
59,6 ± 0,8 |
70,5 ± 0,6 |
118,3 ± 1,1 |
|
Freija x Argamak |
93,1 ± 0,9 А к т и в н о с т ь к и с л о й ф о с ф а |
65,4 ± 1,0 т а з ы, мкг Р/г сухой массы |
70,3 ± 1,1 |
|
Petra x Freija |
0,724 ± 0,057 |
0,866 ± 0,101 |
119,59 ± 14,02 |
|
Freija x Petra |
0,436 ± 0,095 |
0,601 ± 0,056 |
137,81 ± 12,88 |
|
Argamak x Petra |
0,374 ± 0,043 |
0,433 ± 0,005 |
115,70 ± 1,27 |
|
Petra x Argamak |
0,393 ± 0,037 |
0,460 ± 0,065 |
116,97 ± 16,52 |
|
Argamak x Freija |
0,587 ± 0,054 |
1,169 ± 0,052 |
199,23 ± 8,90 |
|
Freija x Argamak ___________ 0,821 ± 0,030 П р и м е ч а н и е. Описание групп см. в разделе «Методика». |
0,798 ± 0,054 |
97,23 ± 6,57 |
|
Влияние исходных родительских форм на активность кислой фосфатазы в I группе было практически равным: материнской, отцовской форм и их взаимодействия — соответственно 32,8; 34,3 и 16,3 %. Во II группе вклад отцовского компонента возрос до 59,2 %, материнского — оказался статистически недостоверным; несколько усилилось взаимодействие компонентов — 19,3 %. Однако степень изменчивости активности фермента под действием ионов Al контролировалась только материнской формой (37,0 %) и взаимодействием компонентов (36,6 %).
По окислительной активности корней действие отцовского и материнского компонентов у гибридов I группы было практически одинаковым (соответственно 13,8 и 14,2 %), а их взаимодействие составляло 61,9 %. При наложении стрессового фактора статистически значимым осталось только влияние взаимодействия компонентов скрещивания (79,3 %); на степень изменчивости показателя под действием ионов Аl оказывал значительное влияние материнский компонент (32,5 %) и взаимодействие обеих форм (41,1 %), влияние отцовского компонента было статистически недостоверным (7,7 %).
Доля влияния материнской формы и взаимодействия компонентов на катионообменную емкость корней в I группе составляла соответственно 33,6 и 35,1 %, влияние отцовской формы было недостоверным — 6,2 %. Примерно такая же картина наблюдалась и при действии стрессового фактора (доля влияния отцовской, материнской форм и их взаимодействия — соответственно 8,6; 37,4 и 42,9 %), а также при оценке степени изменчивости показателя под действием ионов Al (соответственно 8,7; 34,2 и 51,4 %).
По активности кислой липазы у гибридов I группы статистически значимым оказалось только влияние взаимодействия компонентов (93,9 %), которое объясняет практически всю изменчивость в изучаемой популяции. Однако при наложении стрессового воздействия значительно возрастала роль материнского компонента (до 38,2 %), статистически значимой была доля влияния отцовской формы (8,7 %), доля взаимодействия компонентов снижалась в 3 раза (34,5 %). При оценке сте-61
пени изменчивости активности фермента под действием ионов Al вклад этих факторов составлял соответственно 68,8; 5,8 и 10,1 %.
В полевых условиях на нейтральном и кислом участках почвы мы оценивали количественные и качественные сдвиги в работе генетических систем адаптивности и аттракции у родительских сортов овса и их гибридов (рис.). У растений гибрида Petra x Freija выявлена высокая адаптивность на обоих участках, где у родительских форм также отмечены положительные сдвиги по этому признаку. У реципрокных гибридов в этой комбинации скрещивания на нормальной и кислой почве наблюдались соответственно слабые и сильные отрицательные сдвиги по адаптивности. По генетическим системам аттракции у гибридов Petra x Freija и Freija x Petra на нейтральном
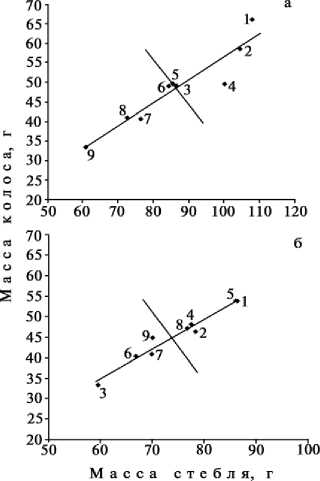
Графический анализ работы генетической системы адаптивности и пластичности сортов и гибридов овса F 1 : а и б — соответственно нейтральная и кислая почва; 1, 2 и 3 — соответственно сорта Argamak, Freija и Petra; 4, 5, 6, 7, 8 и 9 — соответственно комбинации скрещивания Petra x Freija, Freija x Argamak, Freija x Petra, Petra x Argamak, Argamak x Freija и Argamak x Petra.
участке выявлены соответственно положительный и отрицательный сдвиги. Следовательно, первый гибрид имел преимущество перед вторым по работе обеих систем.
У растений гибрида Petra x Argamak на обоих участках наблюдались отрицательные сдвиги в работе генетических систем как адаптивности, так и аттракции. У растений реципрокного гибрида по адаптивности отмечены отрицательные сдвиги, хотя при выращивании на кислой почве показатели значительно возрастали. По работе системы аттракции в нейтральной среде этот гибрид имел средний для выборки показатель (располагался на оси), а в кислой среде — положительные сдвиги.
По адаптивности в нейтральной и кислой среде растения гибрида Argamak x Freija имели соответственно отрицательный и положительный сдвиги. У реципрокных гибридов в обеих средах проявился положительный сдвиг по адаптивности, причем в кислой среде — наибольший в выборке.
Следовательно, по работе генетических систем адаптивности положительные сдвиги при выращивании на кислой почве имели гибриды Freija x Argamak и Argamak x
Freija. Гибрид Petra x Freija располагался в положительной зоне адаптивности, однако величина сдвига последней в этой комбинации бы- ла ниже, чем у других гибридов. В целом сорт Freija, будучи отцовским компонентом, обусловливал положительные сдвиги в генетической системе адаптивности гиб- ридов.
У анализируемых сортов практически отсутствовал полиморфизм по генам аттракции; при выращивании на кислой почве только у растений гибрида Argamak x Petra выявлены положительные сдвиги в работе этой генетической системы.
Среди родительских форм наибольшей адаптивностью обладал сорт Argamak, а наименьшей — Petra; сорт Freija занимал промежуточное положение. Оба гибрида между сортами Petra и Argamak в нейтральных условиях проявляли отрицательный гетерозис по генетическим системам адаптивности, на кислых почвах — промежуточное наследование по этому признаку с отклонением в сторону худшей родительской формы. Отклонение по работе генетической системы адаптивности шло в сторону родительской формы Petra.
У прямых и обратных гибридов между сортами Petra и Freija при выращивании на нейтральной почве выявлено соответственно промежуточное наследование системы адаптивности и слабый отрицательный гетерозис, на кислой почве — соот- ветственно полное доминирование и промежуточное наследование с небольшим отклонением в сторону худшей родительской формы. У обоих гибридов отклонение по работе генетической системы адаптивности шло в сторону отцовского компонента скрещивания. У гибридов между сортами Argamak и Freija на нейтральной почве проявлялся сильный отрицательный гетерозис по генам адаптивности, на кислой почве оба гибрида были практически идентичны по работе генетической системы адаптивности со своими отцовскими компонентами.
Таким образом, на алюмоустойчивость гибридов овса F1 и работу генетической системы адаптивности достоверное влияние оказывает материнский компонент скрещивания. Материнская форма в большей мере влияет на развитие корневой системы гибридов, а отцовская — на параметры работы генетической системы адаптивности. Изменение направления скрещивания отражается на характере взаимодействия генов, кодирующих физиолого-биохимические признаки алюмоустойчивости.
Л И Т Е Р А Т У Р А
-
1. Д р а г а в ц е в В.А. Алгоритмы эколого-генетической инвентаризации генофонда и конструирования сортов сельскохозяйственных растений по урожайности, устойчивости и качеству. СПб, 1997.
-
2. Ж у ч е н к о А.А. Адаптивная система селекции растений (эколого-генетические основы). М., 2001, т. 1.
-
3. Л и с и ц ы н Е.М. Методика лабораторной оценки алюмоустойчивости зерновых культур. Докл. РАСХН, 2003, 3: 5-7.
-
4. L i s i t s y n E.M. Intravarietal level of aluminum resistance in cereal crops. J. Plant Nutrition, 2000, 23(6): 793-804.
-
5. П е т е р б у р г с к и й А.В. Практикум по агрономической химии. М., 1975.
-
6. К л и м а ш е в с к и й Э.Л., Ч у м а к о в с к и й Н.Н. Идентификация форм растений, отзывчивых на
удобрение. Вест. с.-х. науки, 1986, 3: 71-77.
-
7. Методы биохимического исследования растений. Л., 1972.
-
8. Методика государственного сортоиспытания сельскохозяйственных культур. М., 1989.
ГУ Зональный НИИ сельского хозяйства Поступила в редакцию 24
Северо-Востока им. Н.В. Рудницкого, января 2005 года
610007, Киров, ул. Ленина, 166-а
ABOUT INFLUENCE OF PARENT COMPONENTS OF CROSS
ON ADAPTATION OF F 1 OAT HYBRIDS AGAINST HEIGHTENED CONTENTS OF ALUMINUM IONS IN CULTURAL MEDIA
Е.М. Lisitsyn., G.A. Batalova
S u m m a r y
Influence of parent components of cross on a development of different physiological and morphological parameters of aluminum resistance of oat F 1 hybrids was studied under laboratory and field conditions. The character of genes interrelations determining the plants development was estimated. The significant influence of a direction of cross both on development of the studied parameters, and on nature of a gene interaction encoding their determinants was established.
Новые книги
А б а к у м о в В.А., К а л а б е к о в А.Л. Планетарная экологическая система. М., 2002, 674 с.
В монографии сформулирована концепция биосферы как планетарной экологической системы. Рассматриваются пространственная и временная структуры этой системы, а также уровни ее организации. Отражены особенности термодинамики планетарной экологической системы и биологической продуктивности биосферы. Обсуждаются проблемы нерационального использования ресурсов и загрязнения окружающей среды. Проанализированы закономерности изменения биоценозов в условиях антропогенного воздействия. Представлена концепция экологического мониторинга и экологиче- ского нормирования антропогенных воздействий.
Рекомендации по применению препарата Иммуноцитофит в системах защиты зерновых, зернобобовых культур, сахарной свеклы, подсолнечника и плодово-ягодных насаждений от болезней и стрессовых факторов (на примере Черноземной зоны РФ). Пущино, 2003, 38 с.
В рекомендациях обобщены результаты многолетних исследований (1994-2002 годы), проводимых на растениях зерновых, зернобобовых культур, сахарной свеклы, подсолнечника, гороха, яблони и черной смородины, по использованию многоцелевого стимулятора защитных реакций, роста и развития растений, полученно- го на основе арахидоновой кислоты, — имму-ноцитофита. Обсуждается возможность фито-санитарного, ростактивирующего и антистрессового воздействия препарата при наиболее вероятных фитосанитарных и агроэкологических ситуациях. Продемонстрирована целесообразность применения иммуноцитофита в системах защиты различных селькохозяйствен-ных культур от комплекса вредоносных объектов, в том числе в составе баковых смесей с гербицидами, инсектицидами, фунгицидами, удобрениями. Приведены схемы использования препарата, разработанные на основе апробации в производственных условиях Черноземной зоны РФ. Отмечена универсальность препарата и возможность применения в других агроэкологических зонах.
З а р и п о в а О.А., К и р е й ч и к о-в а Л.В., М о ж а й с к и й Ю.А. Микробоценоз почвы при разных уровнях антропогенного воздействия . Рязань, 2004, 162 с.
В монографии представлены результаты многолетних натурных исследований микробиологической активности почвы при разных уровнях антропогенного воздействия (на примере техногенного воздействия Рязанской ГРЭС и орошения сточными водами ОАО «Рязанский свинокомплекс»). Описаны природно-клима-ические условия Рязанской области. Дано обоснование необходимости орошения агроландшафтов природными и сточными водами. Показано, что эффективность использования сточных вод свинокомплексов во многом зависит от выбора технологии, норм, срока, способа полива растений и соблюдения природоохранных мероприятий. Приведены сведения о функционировании и устойчивости микробоценозов. Выявлена взаимосвязь между устойчивостью микробоценозов и количеством органического вещества в почве, которое не только служит субстратом для микроорганизмов, но и обусловливает буферность почв по отношению к различным видам загрязнения. Дана оценка влияния антропогенных факторов на микробо-ценозы серой лесной почвы. Представлены результаты оценки целлюлозоразрушающей активности почвы при агро- и техногенном загрязнении. Проанализировано санитарно-эпидемиоло-ическое состояние орошаемой сточными водами почвы. Уделено внимание оценке микробиологической активности почвы при использовании смеси эффективных микроорганизмов «Байкал ЭМ-1».
