Об экосистемных функциях гумусогенного слоя технозёмов в отдалённый период землевания отвалов угледобычи в Сибири

Автор: Артамонова В.С., Бортникова С.Б., Андроханов В.А.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Почвоведение
Статья в выпуске: 1, 2024 года.
Бесплатный доступ
Создание продуктивных почв для повышения урожайности возделываемых культур известно с доисторических времен. В настоящее время восстановление почвенных свойств актуально для территорий, пострадавших от добычи полезных ископаемых. В Сибири многолетняя добыча угля привела к значительному разрушению почвенного покрова, накоплению на дневной поверхности отвалов вскрышных и вмещающих пород. Это обусловило поиск ресурсосберегающих технологий возмещения почвенных потерь, в том числе путём формирования искусственных почвоподобных тел с нанесённым плодородным слоем для последующего сельскохозяйственного использования. В данной статье изложены результаты исследования физико-химических и микробиологических свойств гумусово-аккумулятивного слоя после 40 лет землевания техногенных отходов в трёх природных зонах (подтаёжной, лесостепной, сухостепной) с целью характеристики экосистемных функций, обеспечивающих плодородие. Установлено, что гумусогенный слой многолетних технозёмов обладает экосистемными функциями фоновых почв, однако в лесостепной и сухостепной зоне обнаруживает признаки снижения устойчивости к повышенным значениям рН, повышения содержания экотоксикантов, уменьшения запаса органического углерода и пула метаболических активных азотфиксирующих бактерий. В аридных условиях диагностируется наибольшее биогенное обеднение технозёмов, проявление инсертационной мутации бактериальных популяций, обеспечивающее их диссоциирование и сохранение в экстремальных местообитаниях. Информация может быть использована при характеристике экосистемных функций и экосистемных сервисов многолетних технозёмов, в прогнозах результативности землевания техногенных отходов в разных природных зонах, учтена при разработке природоподобных технологий.
Почва, природные зоны, экосистемные функции, азотобактер, тяжёлые металлы, мышьяк
Короткий адрес: https://sciup.org/147243413
IDR: 147243413 | УДК: 631.4, | DOI: 10.17072/1994-9952-2024-1-92-106
On ecosystem functions of the technozem humusogenic layer in the remote period of soiling of coal mining waste dumps in Siberia
People strove for creating productive soils to increase the yield of cultivated crops since prehistoric times. Currently, restoration of soil properties is relevant for areas affected by mining. In Siberia, long-term coal mining has led to significant destruction of the soil cover, accumulation of overburden and host rocks on the daytime surface. This initiated the search for resource-saving technologies to compensate for soil losses, including methods of formation of artificial soil-like bodies with a deposited fertile layer for subsequent agricultural use. This article presents the results of a study of physicochemical and microbiological properties of the humus-accumulative layer after 40 years of man-made wasteland soiling in three natural zones (sub-taiga, forest-steppe, dry-steppe) in order to characterize the ecosystem functions that ensure fertility. It was established that the humus layer of perennial technogenic soils (technozems) has ecosystem functions of background soils, however, in the forest-steppe and dry-steppe zones it shows signs of decreased resistance to elevated pH, increased ecotoxicants, reduced organic carbon reserves and a pool of metabolic active nitrogen-fixing bacteria. In arid conditions, the greatest biogenic depletion of technozems is diagnosed, the manifestation of an inertial mutation of bacterial populations, ensuring their dissociation and preservation in extreme habitats. The information can be used to characterize ecosystem functions and ecosystem services of long-term technozems, in forecasts of the effectiveness of man-made wasteland management in different natural zones, and taken into account in the development of nature-like technologies.
Текст научной статьи Об экосистемных функциях гумусогенного слоя технозёмов в отдалённый период землевания отвалов угледобычи в Сибири
С древних времён в земледелии разных стран мира были востребованы искусственно созданные почвоподобные образования агрогенного назначения с добавками аллохтонного (привнесённого) твёрдофазного материала, которые в Мировой системе почвенной классификации входят в группу Антросоли. В ней присутствуют почвы насыпного типа происхождения, например чинампы – на водно-болотных угодьях в Америке, плаггены – на флювиогляциальных или древнеаллювиальных отложениях, суглинках, бурых лесных почвах и подзолах в Северной Европе, хейлуту – на лессе Лессового Плато в бассейне р. Хуанхэ в Китае и другие. Возраст некоторых из них датируются 1–2 тысячелетиями до нашей эры, что подтверждается археологическими раскопками и космическими снимками, полученными с Landsat 7 и мультиспектральных изображений Quickbird VHR, интегрированных в систему ГИС [Hirst, 2021]. Такие объекты включены Организацией Объединённых Наций в состав Всемирного сельскохозяйственного наследия [Всемирным …, 2018].
Примером почв насыпного типа в России являются шиферные почвы Крыма, образованные из плотного юрского глинистого сланца и суглинистых почв – продуктов выветривания этого сланца [Костычев, 1892], на поверхность которых наносились различные почвоулучшители, преимущественно с целью возделывания виноградников.
В ХХ в. список искусственно созданных почв пополнился почвоподобными конструкциями агроген-ного назначения на территориях недропользования в связи острой необходимостью компенсации почв, утраченных при добыче полезных ископаемых. Технологии формирования искусственно созданных почвоподобных тел или образований (ИСПТ) рассматриваются как ресурсосберегающие, поскольку в почвообразование вовлекаются техногенные отходы, например вскрышные и вмещающие породы угледобычи, и отчуждённый ранее плодородный слой зрелых почв. Его привнос предполагает повышение биологической продуктивности нарушенных земель, ускорение восстановления экологических функций почв и продуктивности, а также видового разнообразия экологических систем 1. Период восстановления почвенно-растительного покрова после биологического этапа рекультивации нарушенных земель зависит от природно-климатических условий: скорости и направленности процессов почвообразования, биологической активности почв, гидротермических особенностей, длительности вегетационного периода, мощности и качества нанесенного плодородного слоя почвы, состава растительности прилегающей территории и естественных ландшафтов, последующего хозяйственного использования рекультивируемых земель.
В Сибири первые рекомендации по созданию ИСПТ, в том числе путём технозёмообразования, были представлены в 70-е годы прошлого века. ИСПТ с горизонтом Апах, так называемые технозёмы гумусовоаккумулятивные, созданные на участках с землеванием спланированных отвалов вскрышных пород в районах добычи каменных углей, встречаются поныне на территории Кузбасса, КАТЭКа, Хакасии. Изначально они предназначались под посев многолетних трав, в том числе бобовых, затем – под сенокосы и пастбища, в настоящее время встречаются зачастую в виде залежей.
В Международной реферативной базе почвенных ресурсов технозёмы занимают классификационное положение Technosols. В 2019 г. реферативные группы Антросоли и Техносоли были объединены М.И. Герасимовой в единую общность на основании ряда схожих диагностических признаков и артефактов [Герасимова, 2019]. Технозёмы, состоящие из насыпного грунта с поверхностным плодородным слоем, формируемые при рекультивации целенаправленно, функционируют как почвы [Герасимова и др., 2003]. Исходя из этого, можно заключить, что технозёмам с интродуцированным плодородным слоем присущи схожие экологические функции почв: экосистемные (биогеоценотические) и глобально-биосферные [Национальный…, 2011]. Первая группа функций в значительной степени обусловливается физикохимическими и микробиологическими свойствами. В последние годы биогеоценотические функции почв оказались объектом научных интересов на бывших сельскохозяйственных территориях и леса, в отношении почвоподобных материалов и техногенных почв [Бондаренко, 2016; Васенев и др., 2018; Ананьева, Иващенко, Сушко, 2021; Ананьева и др., 2023; Довлетярова, 2023]. В некоторых случаях они представлены как экосистемные сервисы. Технозёмы с гумусово-аккумулятивным горизонтом с таких позиций ранее не рассматривались. Сведения об участии азотфиксирующих представителей педобиоты в сохранении и активности биогеоценотических функций в многолетних технозёмах представляются своевременными.
Известно, что многолетняя распашка почв ведёт к снижению его запасов и азота [Назарюк, 2002], потере прочной фиксации свежеобразованных соединений гумусовых соединений, препятствующей разрушительной деятельности микроорганизмов [Шарков и др., 2010], изменению состава термических фракций, уменьшению доли пиролизатов слаборазложенных компонентов, в том числе азотсодержащих соединений [Фарходов, 2022]. Можно предположить, что конверсия гумусово-аккумулятивного горизонта фоновых почв на поверхность техногенных отходов и его вовлечение в распашку также будет сопровождаться истощением запасов исходного лабильного органического азотсодержащего вещества, особенно при доминировании растительных остатков с малым отношением С : N (белковые соединения энергично подвергаются процессам трансформации, вовлекаются в обменные процессы гетеротрофного звена азотного цикла). Результаты исследования 20-летних технозёмов после посевов бобовых культур, когда наблюдался рост количества азотсодержащих органических соединений, свидетельствуют о том, что стабилизации азотного режима к этому сроку не достигалось [Андроханов, Курачёв, 2010]. При этом азот микробных продуцентов, в том числе азотфиксирующей бактерии Azotobacter chroococcum, которая является активным природным поставщиком в корнеобитаемый слой растений доступных форм азота, ростстимулирующих и других экзометаболитов и относится к группе Plant-Growth-Promoting-Rhizobacteria – PGPR [Chennappa et al., 2022], не учитывался.
В последние годы представители рода Azotobacter, особенно широко распространённый вид Az. сhroococcum, находятся в сфере интересов международного научного сообщества, особенно США и Индии. В 1990–2020-е гг. за рубежом наметилась восходящая тенденция публикаций, посвящённых бактерии, о чём свидетельствует библиометрическая информация базы данных Scоpus [Aasfar et al., 2021]. Результаты исследований, освещенные в кластерах «Agricultural and Biological Science» и «Environmental Sсience», занимают 2 и 4-е места соответственно. При этом акцентируется внимание на недостаток сведений о влиянии эдафических свойств почв на развитие популяций диазотрофной бактерии, как и её метаболической активности в корневой зоне растений агроценозов [Rajakaruna, Boyd, 2019; Aasfar et al., 2021].
Пристальное внимание исследователей к данной бактерии обусловлено также тем, что штаммы Az. chroococcum считаются наиболее полезными в качестве фиксатора молекулярного азота атмосферы и экскретора (выделителя) аммиака, который идёт на клеточный синтез аминокислот и затем белка. Аэробное поглощение N 2 азотобактером происходит активнее, чем другими свободноживущими диазотрофными бактериями и, следовательно, приток минерализуемых белков до аминокислот, а затем их дезаминирование с выделением NH 3 представляется значимым.
Абсолютная численность Az. chlorococcum является показателем биологической активности, характеризующим плодородие земель сельскохозяйственного пользования1 и качества (плодородия) почв в пахотном горизонте2, входит в состав показателей эколого-гигиенической оценки опасности отходов про- изводства1. Кроме этого, обогащение корнеобитаемой зоны минеральным азотом бактериального происхождения считается экологически безопасным процессом, поскольку исключается загрязнение объектов окружающей среды, что происходит при использовании технического азота и бесподстилочного навоза [Завалин, Соколов, Шмырева, 2019].
Всё вышеизложенное послужило основанием для исследования физико-химических свойств и популяций гетеротрофной азотфиксирующей бактерии – Az. chlorococcum гумусогенного слоя 40-летних тех-нозёмов подтаёжной, лесостепной и сухостепной зон Сибири с целью выявления в нём экосистемных функций.
Материалы и методы исследований
Исследованы ИСПТ (Technosols) с привнесённым гумусовым слоем и фоновые почвы, входящие в состав реферативных почвенных групп – Chernozems, Kastanozems [Герасимова, 2019], они расположены в разных природных зонах Сибири: подтаёжной (южная граница Кузнецкой котловины, Кузбасс), лесостепной (Назаровская котловина, КАТЭК Красноярский край), сухостепной (Минусинская котловина, Хакасия). Пробы для исследований отобраны в течение двух суток с постоянных участков наблюдений лаборатории рекультивации ИПА СО РАН (точки отбора: 807, 810, 811, 868, 897, 899). Площадь участков с технозёмами в подтаёжной зоне составляла 50 га, в лесостепной - 90 га, в сухостепной – 0.5 га. Фоновые почвы подтаёжной и лесостепной зон представлены чернозёмом оподзоленным и выщелоченным, в сухостепной зоне – тёмно-каштановой почвой (в соответствии с отечественной классификацией), почвоподобные образования – технозёмами гумусово-аккумулятивными (во всех трёх случаях). Отбор проб осуществлялся из гумусированного слоя технозёмов и фоновых почв. В каждом конкретном случае составляли средний – смешанный – образец, учитывая рекомендации, приведённые в соответствующих ГОСТах2, поскольку нормативные требования к отбору образцов с участков технозёмов в настоящее время не существуют. Смешанный образец готовили из суммы точечных проб, взятых методом конверта в 5-кратной повторности (5 прикопок), которые смешивали и помещали в полиэтиленовые пакеты. Точечные пробы отбирали до глубины 20 см, поскольку при создании почвоподобных образований в Сибири почвенный слой, что наносился на породу, не превышал 20–35 см в связи с тем, что он лимитировался незначительной мощностью гумусовых горизонтов зональных почв, в отличие от почв европейской части страны. Для почвенно-микробиологических анализов отбирали пробы из смешанного образца в повторностях, указанных в статье. Определение содержания С орг , актуальной кислотности, гранулометрического состава проводили традиционными в почвоведении методами с участием инженеров лаборатории географии и генезиса почв ИПА СО РАН. Предпочтение определению элементарных почвенных частиц по Н.А. Качинскому было отдано в связи с преимуществами метода (в сравнении с методом дифрактометрии), высказанными ранее при интерпретации данных [Болдырев и др., 2019].
Результаты определения химического состава почв, почвоподобных тел получены методом энергодисперсионного рентгенофлуоресцентного анализа с использованием синхротронного излучения (РФА-СИ) на станции синхротронного и терагерцового излучения в ИЯФ СО РАН. Ошибка определения для отдельных элементов находится в пределах 5–15%.
Азотобактер выявляли в лабораторных условиях методом обрастания (встречаемости, колонизации) почвенных комочков (порций, крупиц), традиционным в микробиологии. Данный метод наиболее приближен к естественным условиям обитания азотобактера [Aquilanti, Favilli, Clementi, 2004]. Появление обрастаний мелкозёма колониями бактерии учитывали через 12, 24, 36, 48, 72 часов, поскольку сроки обрастания почв, приводимые в литературе, разнятся. Пробы мелкозёма предварительно размещали в 3 чашки Петри (по 50 штук в каждую). Использовали голодную агаризованную питательную среду Эшби с маннитом в качестве источника углерода (спирт способствует затвердению среды и медленнее, чем глюкоза, утилизируется жизнедеятельными бактериями). Фенотипическое разнообразие колоний азотобактера учитывали по присутствию типичной формы и диссоциантов. Регистрировали штаммы, продуцирующие антибиотики и пигменты. Экспериментальные данные статистически обработаны с использованием компьютерной программы (Statistica 6.0).
Результаты и их обсуждение
Исследования показали, что в технозёмах подтайги и лесостепи содержание илистых частиц и физической глины по сравнению с фоновыми почвами увеличивается, в то время как в сухой степи, наоборот, уменьшается (табл. 1). Суммарное количество частиц пыли и ила, имеющих наибольшую развитую поверхность и, следовательно, адгезивную способность, в технозёме и фоновой почве, расположенных в зоне подтайги, оказалось близким, в то время как в технозёме лесостепи увеличилось в 1.4 раза по сравнению с фоном; в сухой степи, наоборот, снизилось в 1.1 раза. Не исключено, что это могло быть обусловлено различным минералогическим составом подстилающих пород. При этом актуальная кислотность технозёмов в зоне подтайги и сухой степи достоверно повысилась по сравнению с фоновыми почвами (табл. 2). Кислотность технозёма лесостепи изменилась по сравнению с фоном незначительно. Судя по ходу кривой кислотности технозёмов (рис. 1), проявляется тенденция к их защелачиванию.
Таблица 1
Гранулометрический состав гумусогенного слоя фоновых почв и технозёмов в разных природных зонах Западной Сибири
[Granulometric composition of the humus of the fire layer of background soils and technozems in different natural zones of Western Siberia]
|
Место отбора Place of sampling |
Размер фракции (в мм и её содержание в (%) Fraction size (in mm and its content in (%) |
||||||
|
1-0.25 |
0.250.05 |
0.05 0.01 |
0.01 0.005 |
0.005 0.001 |
<0.001 |
<0.01 |
|
|
Подтайга/Subtaiga 1 |
|||||||
|
Фон, чернозём оподзоленный, т. 811/Background, chernozem podzo-lized, p. 811 |
0.07 |
20.19 |
39.72 |
10.52 |
18.88 |
10.62 |
40.02 |
|
Технозём,т. 810/ Technozem, p. 810 |
0.12 |
17.5 |
40.46 |
10.24 |
17.72 |
13.96 |
41.92 |
|
Лесостепь/Forest steppe |
|||||||
|
Фон, чернозём выщелоченный, т. 807/Background, сhernozem leached, p. 807 |
0.13 |
15.91 |
44.04 |
9.40 |
16.76 |
13.76 |
39.92 |
|
Технозём, т. 899/ Technozem, p. 899 |
0.09 |
4.31 |
41.72 |
11.48 |
22.36 |
20.04 |
53.88 |
|
Сухая степь/Dry Steppe |
|||||||
|
Фон, тёмно-каштановая почва, т. 868/ Background, dark chestnut soil, p. 868 |
3.05 |
22.95 |
31.62 |
12.44 |
19.32 |
10.62 |
42.38 |
|
Технозём, т. 894/ Technozem, p. 894 |
10.67 |
40.73 |
14.84 |
7.12 |
15.84 |
10.80 |
33.76 |
В свою очередь, содержание С орг в технозёмах лесостепной и степной зон снижается, в наибольшей степени в степи – до 2.3% по сравнению с фоном (табл. 2).
Таблица 2
Показатели рН и С орг технозёмов в сравнении с фоном (n=3)
[pH and C organic parameters of technozems in comparison with the background (n=3)]
|
Показатель/ Parameter (M ± m) |
Подтайга/ Subtaiga |
Лесостепь/ Forest steppe |
Степь/ Steppe |
|||
|
Чернозем оподзолен-ный / Сherno-zem podzolized |
Технозем/ Technozem |
Чернозем выщелоченный/ Сhernozem leached |
Технозем/ Techno-zem |
Темнокаштановая почва/ dark chestnut soil |
Технозем/ Techno-zem |
|
|
рН водн. |
5.83±0.01 |
6.45±0.02 |
6.75±0.03 |
6.37±0.01 |
7.8±0.01 |
9.08±0.01 |
|
С орг |
4.91±0.05 |
6.32±0.04 |
7.53±0.04 |
4.70±0.14 |
4.2±0.05 |
2.34±0.05 |
Этому не способствует исходно низкая доля корней, участвующих в формировании общего запаса подземного растительного вещества в фоновых почвах степей Минусинской котловины [Швабенланд, 2002]. Помимо этого, синтез и выделение корневых метаболитов в корнеобитаемый слой ослаблены, поскольку уменьшилась обводнённость протоплазмы наземных частей растений, которая рассматривается как физиологическая защита ксерофитов.
Ход кривой С орг технозёма сухой степи и лесостепи свидетельствует о резком снижении в них биогенного элемента по сравнению с фоновыми почвами (рис. 2). В технозёме подтайги потери содержания биогена не произошли.
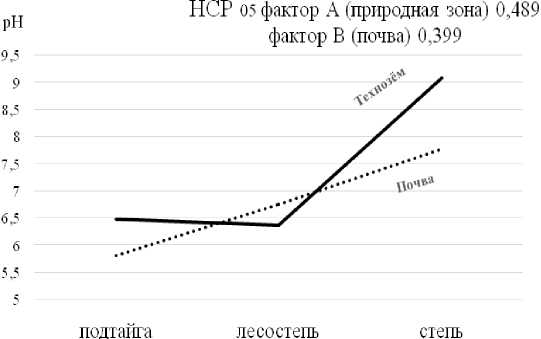
Рис. 1. Ход изменения рН в технозёмах и почвах в разных природных зонах [The course of pH changes in technozems and soils in different natural zones ]
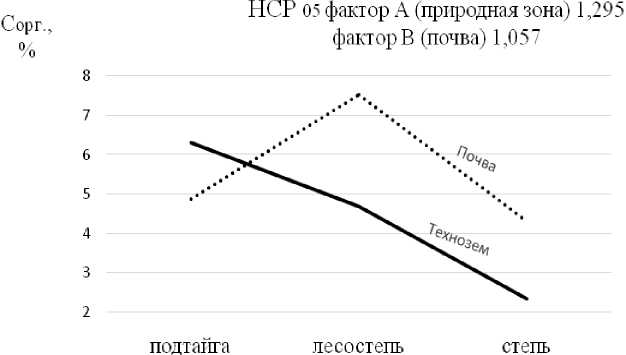
Рис. 2. Ход изменения С орг в технозёмах и почвах в разных природных зонах
[The course of changes in organic matter in technozems and soils in different natural zones ]
Сравнение содержания гетеротрофного азотобактера в технозёмах и фоновых почвах трёх природных зон показало, что наибольшее количество метаболически активных клеток присуще подтаёжной зоне. Наибольшие темпы обрастания проб мелкозёма колониями бактерии присущи фоновой почве подтайги, где повсеместная (100%-ная) встречаемость зарегистрирована уже через 12 ч., но пробы технозёма обрастали медленнее – 24 ч. В лесостепной зоне обрастание мелкозёмистых частиц технозёма занимало также 24 ч., как и в фоновой почве, в степной зоне (в тех же вариантах) – 36 ч. Следует сказать, что значения кислотности технозёмов и фоновых почв трёх природных зон располагались в диапазоне рН = 5.8– 9.09, который соответствовал диапазону роста азотобактера (рН = 4.5–9.0). Оптимальной кислотностью для бактерии считается рН = 6.8–7.2, что зарегистрировано лишь в фоновых почвах лесостепи и сухой степи. Но их заселённость бактерией оказалась ниже, чем в фоновой почве подтайги, кислотность которой оказалась ниже границы оптимального роста. Вероятно, в лесостепи и степи содержание метаболически активных клеток сдерживалось щелочностью, которая сопровождается увеличением времени генерации, снижением числа делений клеток в сутки. При этом не следует исключать пребывание азотобактера в анабиотическом состоянии (от греч. Anabíōsis – «возвращение к жизни»), названным как «немножко умереть» [Пучков, 2015]. Прорастание покоящихся форм бактерии (цист) происходит в течение 4–6 ч. [Wyss, Neumann, Socolofsky, 1961], деление вегетативных клеток – 40–70 мин., т.е. переход жизнеспособных клеток в статус жизнедеятельных присутствовал при инкубации проб в течение 24–36 ч. Это свидетельствует о том, что технозёмы этих зон сохранили функцию депонирования клеток азотобактера, как и фоновые почвы. Однако существенное снижение содержания органического углерода, необходимого для С-гетеротрофии бактерии, как и защелачивание среды обитания, наиболее выраженное в технозёме сухой степи, не способствовали активной жизнедеятельности бактерии в них. К тому же возрастание щелочности технозёмов сопровождалось поглощением катионов металлов и неметаллов гумусогенным слоем, превышающем фоновые почвы в большинстве случаев (табл. 3). По валовому количеству тяжёлых металлов и неметаллов, определённому ранее для территории нашего обследования [Богуславский и др., 2021], лидировали Zn, Pb, Cu, Ni, V и As, числовые значения которых превышали их кларки [Архипов, 2011; Касимов, Власов, 2015] и ОДК/ПДК1. О присутствии металлов и неметаллов в почвах степной зоны сообщалось ранее [Язиков, Худяков, Азарова, 2002; Архипов, 2011; Швабенланд, Можайский, 2011], в том числе в биотоксичных концентрациях [Азарова, Васиков, Ильинских, 2004]. Средние значения содержания мышьяка на рекультивируемом отвале в Минусинской котловине и в почвенном горизонте АВ достигали 12 г/т [Язиков, Худяков, Азарова, 2002].
Таблица 3
Валовое содержание некоторых металлов (мг/кг) в почве и технозёме степной зоны (в районе Черногорского угольного месторождения, Хакасия)
[The total content of some metals (mg/kg) in the soil and technozem of the dry steppe zone (in the area of the Chernogorsky coal deposit, Khakassia)]
|
Почва/Soil |
Металлы/Metals |
|||||
|
Cu |
Ni |
Pb |
Zn |
V |
As |
|
|
Региональный кларк, каштановая почва, мг/кг [Архипов, 2011]/ Regional Clark, chestnut soil, mg/kg |
31.4 |
26.8 |
13.3 |
62.8 |
119 |
Нет |
|
Кларк по Виноградову [Касимов, Власов, 2015]/ Clark by Vinogradov |
47 |
58 |
16 |
83 |
90 |
1.7 |
|
ПДК в почвах / MPC in soils |
3.0 |
Нет |
6.0 |
23.0 |
Нет |
2.0 |
|
ПДК(ОДК) в почвах2 / MPC(ODC) in soils |
132 |
80 |
130 |
220 |
Нет |
10.0 |
|
Самозарастающий участок отвала/ Selfhealing section of the dump* |
61 |
98 |
19 |
127 |
122 |
5.3 |
|
Почва/ Soil* |
51 |
78 |
20 |
106 |
180 |
9.6 |
|
Технозём/ Technozem* |
58 |
101 |
19 |
121 |
151 |
11.3 |
Примечание. * –Богуславский и др. [2021].
К такому химическому составу азотобактер отчасти проявляет определённую толерантность, благодаря механизмам защиты. Установлено [Herter et al., 2013], что высокие концентрации сульфата меди активизируют синтез катехолатных сидерофоров азотфиксирующими меланогенными клетками, обеспечивающими детоксикацию меди. Устойчивость бактерии к Pb(II) достигается адсорбцией металла внеклеточными полисахаридами, исключением свинца из клеток, связыванием его в виде нерастворимых фосфатов, оттоком ионов наружу, экспрессией ионов регуляторными белками. Сообщается [Маlynovska, 2017], что показатель содержания и биохимической активности клеток азотобактера можно считать индикаторным при уровнях загрязнения 5–100 предельно допустимых концентраций Pb в отсутствие фитоценоза, при уровнях загрязнения 10–100 ПДК — на почвах с фитоценозом. Показано [Ngumah et al., 2018], что высокие концентрации ZnSO 4 не снижают биомассу азотобактера, не подавляют азотфикса-цию. В нашем случае бактерия развивалась на фоне высокого содержания кальция в технозёме и почве. Он способен задерживать поступление Zn внутрь бактерий и растений, т.к. эти катионы являются антагонистами. Несмотря на высокую растворимость солей Zn в щелочной среде, металл образует с Ca малорастворимые цинкаты кальция [Шаврина, 2021].
Кроме того, в поддержании диазотрофного питания бактерии мог участвовать ванадий, недостатка которого в технозёме и фоновой почве сухой степи нет. Он входит в состав V-содержащей нитрогеназы, кислородоустойчивой в аэробных условиях. V-нитрогеназа, в отличие от белково-полиметальных кластеров нитрогеназ (Mo-Fe-содержащей и альтернативной Fe- содержащей), наиболее активна в условиях пониженных температур (вплоть до 5º), которые характерны для резко континентального климата Хакасии. А температурный минимум мезофильного азотобактера соответствует 10–15°C, поэтому значение V-нитрогеназы велико. Помимо этого, V является парамагнитным и азотконцентрирующим (в больших количествах) элементом. Находясь в слизи и в цитоплазме бактерии, он ослабляет образование возбуж- дённых молекул кислорода, снижает его негативное воздействие. В свою очередь, углеродсодержащие соединения, присутствующие в полисахаридной оболочке бактерии, выступают агентами гелеобразования, обеспечивая поглощение большого количества воды в присутствии (как правило) двухвалентных ионов. Сорбционная ёмкость полисахаридов по отношению к металлам значительно превышает таковую препаратов угля и лигнина [Хотимченко, 2011]. Это также играет важную роль в сохранении жизнедеятельности влаголюбивого азотобактера.
Особого внимания заслуживают ответные реакции азотобактера на мышьяковистое окружение. As – биотоксикант кумулятивного действия, он ингибирует процессы субстратного и окислительного фосфо-лирирования, блокирует SH-группы белков. Доказано, что бо́льшую чувствительность к высоким концентрациям As проявляют протеолитические микроорганизмы [Окунев, Сунгатуллина, Григорьян, 2015], которые участвуют в расщеплении белков, пептидов, аминокислот растительных остатков. Азотобактер проявляет различные стратегии защиты к мышьяку. Арсенат может осаждаться и на поверхности клеток в виде частиц минерала скородита (FeAsO 4 x 2H2O), который слаборастворим. Экспериментально доказано, что сорбция As полисахаридами цист Azotobacter достигает 65%, в комплексе с керамическими гранулами – 96% [Gauri et al., 2011]. Не исключено, что оксидоредуктазы азотобактера подобно близкородственным микроорганизмам – Pseudomonas aeruginosa и цианобактериям – восстанавливают ион арсената до иона арсенита. Этот путь считается критическим, но он позволяет образовать комплекс со специфическими сульфгидрилсодержащими белками, обеспечивая детоксикацию металлоида. Известно также, что азотобактер способен окислять и метилировать арсенит, восстанавливать арсенат в цитоплазме, что обеспечивает сохранение жизнеспособности бактерии, как и других микроорганизмов [Dhuldhaj et al., 2013]. Кислые гетерополисахариды оболочки частично могут связывать 5-валентный As и вовлекать его в восстановительное метилирование с последующим улетучиванием алкилсерина. Тем не менее, полностью механизмы устойчивости бактерии к металлоиду не раскрыты.
Что касается негативного влияния никеля на бактерию, содержание которого было в технозёме выше, чем в почве, то оно изучено недостаточно. Можно предположить, что его токсичность в щелочном растворе технозёма, как и фоновой тёмно-каштановой почве, мала, поскольку металл обладает пассивацией, т.е. способностью образовывать защитную оксидную плёнку. Водные растворы 2-х валентного металла содержат аква-ион, который осаждается в виде гидроксида никеля.
В свою очередь, факты диссоциирования популяции азотобактера свидетельствуют о присутствии техногенного загрязнения среды обитания бактерии. Причиной SR-диссоциации являются инсерционные мутации, научный интерес к которым в последнее время растёт. Способность бактерий к расщеплению популяций проявляется в разнообразных экологических нишах [Maia, Sanchez, Vela, 1988; Пищик и др., 2016]. Диссоциирование популяции Az. chlorococcum зарегистрировано на отвалах добычи антрацита [Артамонова, Бортникова, Оплеухин, 2016], на отходах цианирования золотосодержащей руды [Артамонова, Бортникова, Оплеухин, 2020].
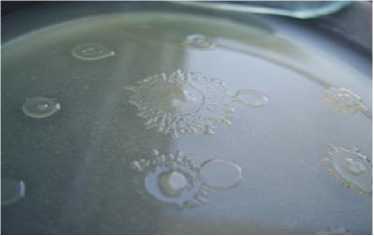
В технозёме и фоновой почве сухой степи диссоциирование проявляется в присутствии атипичных R-, М- и Р- колоний наряду с типичной S-формой (рис. 3). Для P-диссоциантов характерно наличие гидрофобных оболочек клеток и каротиноидных пигментов, которые повышают толерантность клеток азотобактера к токсичным агентам. Гидрофобность клеточных оболочек обусловлена стабильностью отдельных областей молекулы белка, в том числе путём её сворачивания, уплотнения молекул воды, а также образованием мембранных липидов, что ограничивает проникновение водорастворимых металлов внутрь клетки. Каротиноиды поглощают энергию возбуждения электрона без каких-либо химических превращений, возвращают кислород в основное триплетное состояние без повреждения биологических систем клетки, что важно для сохранения нитрогеназы и выживания при недостатке азотного питания. Ранее пигментированный P-дисссоциант – Pseudomonas aurantica, – близкородственный вид азотобактеру, регистрировался на бедной среде с двукратным лимитом источника азота [Мулюкин, Козлова, Эль-Регистан, 2008]. Кроме того, в технозёме сухой степи выявлены диссоцианты с пигментацией и антибиотической активностью. Выделение экзометаболита, очевидно, В 12 , способствовало сохранению способности клеток синтезировать цианокобаламин, поскольку его повышенные концентрации в цитоплазме клетки могут полностью репрессировать синтез В 12 способствовало сохранению цианокобаламина обеспечивает метионин в присутствии кислорода [Sekowska et al., 2004]. Цианокобаламин термостабилен, что проявляется даже в энзиматически неактивной биомассе. Он обладает антимикробным эффектом, повышая активную конкуренцию гетеротрофного микроба за ресурсы питания. Возможно и другое значение «сброса» метаболита за пределы клетки, связанное с хелатной природой соединения, что требует дополнительных исследований для подтверждения предположения. Присутствие в технозёме сухой степи штаммов с антибиотическими свойствами подтверждает конкуренцию гетеротрофов за дефицитную органическую пищу. Следовательно, гумусогенный слой технозёма сухой степи, несмотря на максимальное защелачивание, техногенное загрязнение, подавление илообразования, дефицит органического углерода, снижение биогенности, обеспечил сохранность популяции азотфиксирующей бактерии как неотъемлемой части экосистемы.
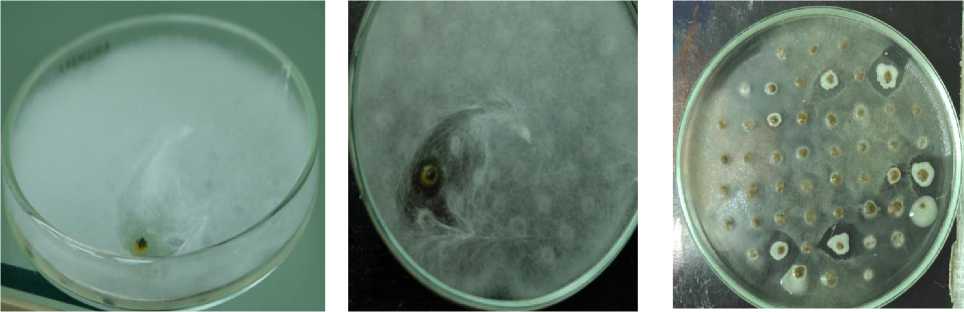
технозём technozem
подстилающая порода bedrock
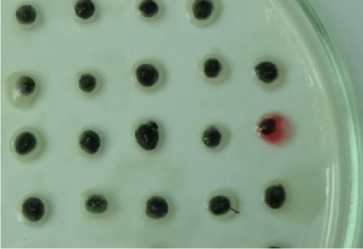

технозём technozem
подстилающая порода bedrock
Диссоцианты/dissociants

S-тип/S-type
R-тип/R-type
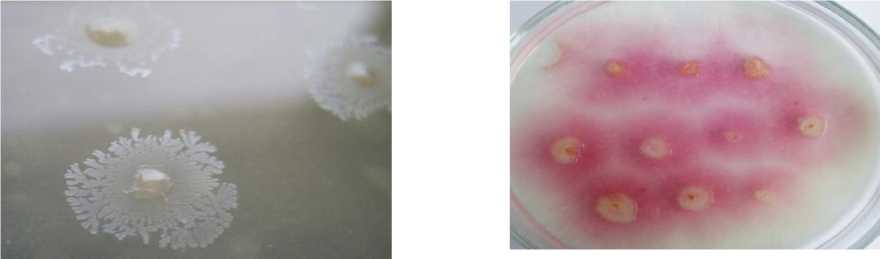
Р-тип/P-type М-тип/M-type
Рис. 3. Внутривидовое разнообразие популяции Az. chroococcum [Intraspecific diversity of the population Az. chroococcum ]
Заключение
После 40 лет землевания отвалов угледобычи в гумусогенном слое технозёмов разных природных зон Сибири сохраняются экосистемные функции фоновых почв. Они проявились на уровне разных категорий: физической, физико-химической, биохимической, целостной. Гумусогенный слой технозёмов сохранил способность к депонированию мелкозёмом клеток азотобактера, обеспечил сохранение его популяций, как и накопление органического углерода, сорбцию химических элементов, илообразование, буферность и другие. Однако в технозёме сухостепной зоны обнаружены некоторые нежелательные признаки. В нём существенно снизилось содержание фракции ила, возросли защелачивание и количество Cu, Zn, Pb, Ni, V, As, уменьшились запас органического углерода, биогенность. Снижение количества илистых частиц, поглотительная способность которых в отношении тяжёлых металлов уменьшается в ряду Pb-Cu-Zn (по мере перехода к цинку) [Пинский и др., 2019], обусловило лидерство Cu и Zn. Присутствие экотоксикантов не оказало бактерицидного действия на популяцию азотобактера, но отразилось на диссоциировании популяции, что можно рассматривать как повышение устойчивости к полиметалльному окружению. Защелачивание снизило заселённость мелкозёма бактерией в связи с уменьшением размножения вегетативных клеток, что не способствует пополнению минеральных форм азота, доступных растениям. Повысить их пул можно путём использования «зелёных» технологий ускоренной рекультивации, в частности с привлечением сидератов, компостов, навоза, торфа, соломы, сапропелей, растительных остатков от пожнивных посевов и покровных культур.
Не следует исключать зарубежные подходы применения местных метаболически активных штаммов азотобактера, адаптированных к техногенному загрязнению и востребованных в фитовосстановлении участков на месте складирования угля [Barman, Dutta, Jha, 2022; Guzmán-Moreno et al., 2022]. Скрининг штаммов местных популяций азотобактера, устойчивых к экотоксикантам и сохраняющих активность размножения, снизит остроту проблемы дефицита биогенного азота в гумусогенном слое сухой степи.
Таким образом, гумусогенный слой многолетних технозёмов гумусово-аккумулятивных проявляет экосистемные функции, свойственные фоновым почвам исследованных природных зон. Однако способность данного слоя противостоять защелачиванию, техногенному загрязнению, а также снижению органического углерода и численности продуцентов биогенного азота в них разная. Наименьшие возможности для экосистемного сервиса складываются в технозёме сухой степи, что следует учитывать при разработке природоподобных технологий.
Список литературы Об экосистемных функциях гумусогенного слоя технозёмов в отдалённый период землевания отвалов угледобычи в Сибири
- Азарова С.В., Васиков У.Г., Ильинских Н.Н. Оценка экологической опасности отходов горнодобывающих предприятий республики Хакасия с применение метода биотестирования // Известия Томского политехнического ун-та. 2004. Т. 307, № 4. С. 55-59.
- Ананьева Н.Д., Иващенко К.В., Сушко С.В. Микробные показатели городских почв и их роль в оценке экосистемных сервисов (обзор) // Почвоведение. 2021. № 10. С. 1231-1246.
- Ананьева Н.Д. и др. Биофильные элементы (С, N, Н) и дыхательная активность микробного сообщества лесопарков Москвы и пригородных лесов // Почвоведение. 2023. № 1. С. 102-117. DOI: 10.31857/S0032180X22600780
- Андроханов В.А., Курачёв В.М. Почвенно-экологическое состояние техногенных ландшафтов: динамика и оценка / отв. ред. А.И. Сысо. Новосибирск: Изд-во СО РАН, 2010. 224 с.
- Артамонова В.С., Бортникова С.Б., Оплеухин А.В. Техногенное загрязнение почв подотвальными водами в районе угледобычи // Известия Коми научного центра УрО РАН. 2016. Вып. 4(28). С. 38-45.
- Артамонова В.С., Бортникова С.Б., Оплеухин А.В. Фитотоксичность лежалых отходов цианирования золотосодержащей руды на территории накопленного экологического ущерба // Вестник Пермского университета. Сер. Биология. 2020. Вып. 1. С. 33-40.
- Архипов А.Л. Геоэкологическое и экогеохимическое состояние геоэкологической среды ЮжноМинусинской котловины (республика Хакасия): автореф. дис. ...канд. геол.-минер. наук. Томск, 2011. 23 с.
- Богуславский А.Е. и др. Геохимический фон тяжёлых металлов в почвах и растениях на участках отвалов угольных месторождений // Известия Алтайского отделения Русского географического общества. Сер. Геология. 2021. № 2(61). С. 40-49.
- Болдырев В.А. и др. Интерпретация результатов определения гранулометрического состава почв методом лазерной дифракции // Фундаментальные концепции физики почв: развитие, современные приложения и перспективы: материалы Междунар. конф. М., 2019. C. 114-118. DOI: 10.31453/kdu.ru.91304.0065.
- Бондаренко Е.В. Опыт учёта экосистемных сервисов почв при оценке деградации земель (на примере УО ПЭЦ МГУ): дис. ... канд. биол. наук. М., 2016. 121 с.
- Васенев В.И. и др. Экологические функции и экосистемные сервисы городских и техногенных почв: от теории к практическому применению (обзор) // Почвоведение. 2018. № 10. С. 1177-1191.
- Всемирным сельскохозяйственным наследием признаны в мире 52 объекта. 2018. URL: https://foodindustry.kz/vsemirnym-selskohozyajstvennym-naslediem-priznany-s-mire-52-obekta/?ysclid=lifr2l8c 18 44759192 (дата обращения: 18.05.2023).
- Герасимова М.И. и др. Антропогенные почвы: генезис, география, рекультивация / отв. ред. Г.В. Добровольский. Смоленск: Ойкумена, 2003. 268 с.
- Герасимова М.И. Международная классификация почв и возможности её применения в географических исследованиях // Вестник Московского ун-та. Сер. 5. География. 2019. № 3. С. 49-56.
- Довлетярова Э.А. Функционально-экологическая оценка почв в условиях антропогенной нагрузки мегаполиса и промышленного предприятия: автореф. дис. ... д-ра биол. наук. М., 2023. 50 с.
- Завалин А.А., Соколов О.А., Шмырева Н.А. Экология азотфиксации. М., 2019. 252 с.
- Касимов Н.С., Власов Д.В. Кларки химических элементов как эталоны сравнения в экогеохимии // Вестник Московского ун-та. Сер. 5. География. 2015. № 2. С. 7-17.
- Костычев П.А. Изслъдоваше почвъ изъ виноградниковъ Крыма и Кавказа //Въестникъ Винодълiя. 1892. № 1, 2. С. 1-23.
- Мулюкин А.Л., Козлова Н.Н., Эль-Регистан Г.И. Свойства фенотипических диссоциантов бактерий Pseudomonas aurantica и Pseudomobas fluorescens // Микробиология. 2008. Т. 77, № 6. С. 766-776.
- Назарюк В.М. Баланс и трансформация азота в агроэкосистемах / отв. ред. В.Б. Ильин. Новосибирск: Изд-во СО РАН, 2002. 257 с.
- Национальный атлас почв Российской Федерации / отв. ред. чл.-корр. РАН С.А. Шоба. М.: Аст-рель, АСТ, 2011. 632 с.
- Окунев Р.В., Сунгатуллина Л.М., Григорьян Б.Р. Влияние арсената (V) натрия на содержание свободных и связанных аминокислот растворённого органического вещества почвы // Вестник Крас. ГАУ. 2015. № 6. С. 9-12.
- Пинский Д.Л. и др. Особенности сорбции меди чернозёмами различного гранулометрического состава // Фундаментальные концепции физики почв: развитие, современные приложения и перспективы: материалы Междунар. конф. М., 2019. C. 385-389. DOI: 10.31453/kdu.ru.91304.0065.
- Пищик В.Н. и др. Механизмы адаптации растений и микроорганизмов в растительно-микробных системах к тяжёлым металлам // Микробиология. 2016. Т. 85, № 3. С.232-247.
- Пучков Е.О. Немножко умереть // Химия и жизнь. 2015. № 1. С. 1-23.
- Фарходов Ю.Р. Молекулярный состав лабильного и стабильного органического вещества типичных чернозёмов разного вида использования: автореф. дис. . канд. биол. наук. М., 2022. 24 с.
- Хотимченко М.Ю. Сорбционные свойства и фармакологическая активность некрахмальных полисахаридов: дис. ... д-ра мед. наук. Владивосток, 2011. 327 с.
- Шаврина К.Ф. Конкурентное взаимодействие цинка, кальция и магния в системе дерново-подзолистая почва - растения: автореф. ... дис. канд биол. наук. СПб., 2021. 26 с.
- Шарков И.Н. и др. Воспроизводство гумуса как составная часть системы управления плодородием почвы: метод. пособие. Новосибирск, 2010. 36 с.
- Швабенланд И.С. Запас лабильного органического вещества в почвах Хакасии // Геоэкологические проблемы почвоведения и оценки земель: материалы Междунар. науч. конф. Томск, 2002. С. 210-211.
- Швабенланд И.С., Мажайский Ю.А. Геолого-почвенная характеристика Минусинского прогиба на примере Черногорского каменноугольного месторождения // Агрохимический вестник. 2011. № 6. С. 2-4.
- Язиков Е.Г., Худяков В.М., Азарова С.В. Геоэкологические проблемы угледобывающих предприятий и геохимическая оценка воздействия отвалов на почвы (на примере угольного разреза Чалпан, республика Хакасия) // Известия Томского политехнического университета. 2002. Т. 305, вып. 6. С. 433-445.
- Aasfar А. et al. Nitrogen Fixing Azotobacter Species as Potencial Soil Biological Enhancers for Crop Nutrition and Yield Stability // Front. Microbiol. 2021. Vol. 12. Р. 1-19. DOI: 10.3389/fmicb.2021.628379.
- Aquilanti L., Favilli F., Clementi F. Comparison of different strategies for isolation and preliminary identification of Azotobacter from soil samples // Soil Biology and Boochemistry. 2004. Vol. 36. P. 1475-1483.
- Barman D., Dutta I., Jha D.K. Heavy metal resistant bacteria from coal dumping site with plant growth promoting potentials // Biologia. 2022. Vol. 77. P. 533-545. URL: https://doi.org/10.1007/s11756-021-00963-y.
- Chennappa Gurikar C. et al. Azotobacter - a potential symbiotic rhizosphere // Rhizosphere Engineering: Academic press, 2022. P. 97-112. URL: https://doi.org/10.1016/B978-0-323-89973-4.00010-7.
- Dhuldhaj U.P. et al. Microbial interactions in the arsenic cycle: adoptive strategies and applications in environmental management // Reviews Environmental Contamination and Toxicology. 2013. Vol. 24. P. 1-38. DOI: 10.1007/978-1-4614-5 882-1 1.
- Gauri Samiran S. et al. Removal of arsenic from aqueous solution using pottery granules coated with cyst of Azotobacter and portland cement: characterization, kinetics and modeling // Bioresource Technology. 2011. 102(10). P. 6308-6312. DOI: 10.1016/j.biortech.2011.02.037.
- Guzman-Moreno J. et al. Bacillus megaterium HgT21: a Promising Metal Multiresistant Plant Growth-Promoting Bacteria for Soil Biorestoration // Microbiology Spectrum. 2022. Vol. 10, № 5: e0065622. DOI: 10.1128/spectrum.00656-22. URL: https://pubmed.ncbi.nlm.nih.gov/35980185/.
- Herter S. et al. Investigating the effects of metals on phenol oxidaseproducting nitrogen-fixing Azotobacter chroococcum // J. Basic Microbiol. 2013. Vol. 53, № 6. P. 509-517. DOI: 10.1002/jobm.201100443.
- Hirst K. Kris. Chinampa: Raised Field Agriculture in the Americas // Chinampa of Floating Gardens. Thought Co, 2021. URL: https://www.thoughtco.com/chinampa-floating-gardens-170337 (дата обращения: 18.05.2023).
- Maia M., Sanchez J.M., Vela G. Plasmids of Azotobacter vinelandii // Journal of Bacteriology. 1988. Vol. 170. № 4. P. 1984-1985.
- Маlynovska 1.М. Effect of heavy metal ions on the numberand activity of Azotobacter and melanin synthesizing micromycetes // Biotechnologia Acta. 2017. Vol. 10, № 3. P. 65-71.
- Ngumah C. et al. Impacts of different concentrations of Copper and Zinc on in vitro responses of Azotobacter chroococum in biomass and nitrogen fixing outputs // Ecotoxicol. Environ. Contam. 2018. Vol. 13, № 1. P. 79-83. DOI: 10.5132/eec.2018.01.09.
- Rajakaruna H., Boyd R.S. Edaphic Factor // Encyclopedia of Ecology, 2019. Bd. 3. P. 361-367.
- Sekowska A. et al. Bacterial variations on the methionine salvage pathway // BMC Microbiology. 2004. Vol. 4, № 9. P. 1-17. URL: https://doi.org/10.1186/1471-2180-4-9.
- Wyss O., Neumann M.G., Socolofsky M.D. Development and germination of the Azotobacter cyst // Journal of Biophysical and Biochemical Cytology. 1961. Vol. 10. P. 555-565.
