Об обнаружении Тульского гербария барона Вальтера Вольдемаровича фон Розена

Автор: Щербаков А.В., Бубырева В.А., Хорун Л.В.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 4 т.10, 2016 года.
Бесплатный доступ
Приводятся сведения о находке считавшегося утерянным гербария известного исследователя тульской флоры барона В.В. фон Розена.
Гербарные коллекции тульская область
Короткий адрес: https://sciup.org/148313865
IDR: 148313865 | УДК: 581.9
Текст научной статьи Об обнаружении Тульского гербария барона Вальтера Вольдемаровича фон Розена
Epipogium aphyllum (F.W. Schmidt) Sw. – сапрофитная орхидея, ареал которой простирается от Центральной Европы на западе до острова Тайвань на востоке (Chung, Tzeng, 2001), от Кавказа на юге до Скандинавского полуострова на севере (Taylor, Roberts, 2011). Несмотря на широкий ареал, надбо-родник включен в Красную книгу России (2008) как вид с сокращающейся численностью.
Бóльшую часть жизни надбородник существует в виде подземного корневища, на поверхности появляются только цветоносные побеги (Александров и др., 2004). Растение полностью лишено хлорофилла. Стебель цветоносных побегов безлистный полупрозрачный, палевого цвета. На стебле заметны несколько чешуевидных влагалищ редуциро- ванных листьев (Taylor, Roberts, 2011). Популяции надбородника в большинстве своем малочисленны и нестабильны. Надбородник, как правило, цветет не ежегодно (перерыв может составлять несколько десятков лет) и после отцветания растение может не проявлять своего присутствия на протяжение многих лет (Аверьянов, 1988; Красная книга…, 2008; Taylor, Roberts, 2011). До сих пор не существует определенных данных о влиянии погодных условий на обилие цветения, хотя есть указание, что цветение более обильно в холодные года (Taylor, Roberts, 2011). Цветоносные побеги разлагаются после нескольких недель своего существования (Chung, Tzeng, 2001).
Морфологию и биологию надбородника описал достаточно подробно P. Rohrbach (1866). Впоследствии это описание дополня-
лось публикациями других авторов, из которых особенно значительный вклад внес P. Vermeulen (1965), хотя многие аспекты до сих пор остаются невыясненными (Taylor, Roberts, 2011). Надбородник достаточно активно размножается вегетативным путём (Александров и др., 2004; Красная книга…, 2008) при помощи небольших протокормо-образных почек, формирующихся на столонах (Roy et al., 2009; Taylor, Roberts, 2011). По внешним признакам невозможно достоверно определить происхождение экземпляра (т.е. появился ли он в результате вегетативного или генеративного размножения). В обоих случаях корневище имеет узкую длинную базальную часть с закруглённым кончиком. Однако экземпляры, полученные путём парти-куляции, имеют широкую и как бы обрубленную, иногда с неровным краем, базальную часть (Irmisch, 1853; Ziegenspeck, 1936).
Цветки надбородника оплодотворяются очень редко, а семена вызревают еще реже (Александров и др., 2004). P. Rohrbach (1866) считает самоопыление у E. aphyllum невозможным из-за особенностей строения цветка (Taylor, Roberts, 2011). В цветках E. aphyllum содержится большое количество нектара (Кернер фон Марилаун, 1906). Сложно устроенный цветок надбородника требует, по-видимому, специфичных опылителей, таксономическая принадлежность которых до сих пор не установлена (Taylor, Roberts, 2011). P. Rohrbach (1866) и А. Кернер фон Марилаун (1906) наблюдали, как шмель Bombus lucorum (Linnaeus, 1761) посещал цветки надбородника, при этом перенос поллиниев не был отмечен.
Надбородник безлистный по данным Е.Г. Воробьевой и А.Е. Панарина (1994) является одним из наиболее уязвимых и нуждающихся в охране видов сосудистых растений, произрастающих на территории Кандалакшского заповедника. Популяция надбородника на острове Асафий Кемьлудского архипелага была впервые обнаружена Н.Е. Богдановой и В.Н. Веховым в 1962 г. (Богданова, Вехов, 1969). При повторных поисках популяцию обнаружить не удалось (Воробьёва, Панарин,
1994). В 2001 г. надбородник был вновь найден на острове Асафий участниками Беломорской экспедиции Московской гимназии на Юго-Западе (Сухов, 2001). Существуют сведения об еще одной популяции надбород-ника в Кандалакшском заповеднике, обнаруженной в 1986 г. А.Б. Георгиевским на острове Великий (Воробьёва, Панарин, 1994). Однако, по мнению проф. Д.Д. Соколова (биологический факультет МГУ, личное сообщение), на о. Великий за Epipogium aphyllum была принята Corallozhiza trifida Chatel, поэтому популяция надбородника на о. Асафий является в настоящее время единственной достоверно известной для Кандалакшского заповедника.
В настоящем сообщении впервые обобщены результаты десятилетних авторских наблюдений за популяцией надбородника на острове Асафий (2001 – 2011 гг.). Некоторые результаты были опубликованы ранее (за 2001 – 2005 гг. – см. Копылов-Гуськов и др., 2007, за 2006 г. – Kopylov-Guskov et al., 2007, за 2007 – 2008 и 2011 гг. – депонированы в Летописи природы Кандалакшского заповедника, см. Волкова, Копылов-Гуськов, 2008, 2009; Волкова и др., 2012). В статье также приведены ранее не публиковавшиеся данные по морфометрии генеративных побегов.
Материал и методика исследований
Все наблюдения проводились в первой половине августа во время массового цветения надбородника (только в 2007 г. наблюдение проведено 29 июля в начале цветения). В течение восьми лет (2001–2004, 2006–2008 и 2011 гг.) подсчитывалось число наземных генеративных побегов в популяции. В 2003, 2004 и 2007 гг. генеративные побеги были закартированы. Картирование проводилось методом обратной засечки (измерение азимута от растения на два репера на местности), что позволяет свести к минимуму повреждения растительного покрова при картировании и нанести растения на схему с ошибкой, не превышающей 10% расстояния от растения до репера (Говорухин и др., 1980); то есть не более двух метров в нашем случае. Побеги отмечались на схеме отдельно, если расстояние между ними превышало один метр. В 2005 г. были зарегистрировали виды сосудистых растений, произраставшие в непосредственной близости (на расстоянии менее 10 см) от измеренных цветоносных побегов. В течение трех лет проводился подсчет цветков в соцветиях и измерялась высота генеративных побегов (в 2005 г. – у 50 случайно выбранных побегов, в 2006 и 2008 гг. – у всех обнаруженных побегов).
В 2005 г. исследовались подземные органы и производилась оценка потенциальной семенной продуктивности надбородника. Для достижения первой задачи были осмотрены подземные органы четырёх одиночных генеративных побегов и четырёх групп побегов по 2 – 4 побега (частицы подстилки были аккуратно приподняты с помощью пинцета и затем возвращены на место). Для достижения второй задачи четыре соцветия (один цветок, два цветка и два соцветия по три цветка в каждом) были зафиксированы 70% этанолом (выбраны соцветия, уже поврежденные животными). Каждая завязь в лабораторных условиях вскрывалась и извлекались семязачатки, которые помещались в смесь глицерина со спиртом и по возможности равномерно распределялись по чашке Петри с нанесенной сеткой. После подсушивания препарата были подсчитаны все семязачатки, причем подсчет повторялся дважды или трижды (средняя погрешность подсчета составила 109 семязачатков).
В 2005–2006 гг. проводились непрерывные наблюдения за несколькими цветущими растениями для выявления потенциальных опылителей. 9–11 августа 2005 г. проводились наблюдения за одиночным цветоносным побегом и за двумя группами побегов различной плотности (3 шт/м2 и 9 шт/м2) с расстояния 1–1,5 метра от соцветия. Всего состоялось пять сеансов непрерывных наблюдений общей продолжительностью 20 часов: два сеанса в первой половине дня (с 9:30 до 13:30), два сеанса во второй половине дня (с 15:30 до 20:30) и один сеанс ночью (с 00:00 до 02:00). Во время наблюдений стояла безвет- ренная ясная погода, температура воздуха колебалась от 17 °C утром и вечером до 27 °C днем. 10–11 августа 2006 г. наблюдалась группа побегов (примерная плотность 9 шт/м2). Состоялось четыре сеанса непрерывных наблюдений общей продолжительностью 11,5 часов: три дневных (с 17:00 до 20:40; с 11:00 до 14:40 и с 16:45 до 19:00) и один ночной (с 0:00 до 2:00). Ночью наблюдения проводили с искусственной подсветкой светодиодным фонарем. Погода во время наблюдений была безветренной; первый день наблюдений был пасмурным и прохладным (температура воздуха около 17 °C), второй день – ясным и теплым (около 24 °C). Во время наблюдений фиксировалось время, когда насекомое посетило цветок, отмечался отряд насекомого, изучалось поведение насекомого на цветке. После того, как насекомое покидало цветок оно, по возможности, отлавливалось с помощью сачка и более точно определялась таксономическая принадлежность по определителям (Определитель насекомых…, 1948; Горностаев, 1999).
Результаты
Фитоценотическая приуроченность и морфологическое описание
Исследуемая популяция надбородника находилась в сыром разнотравном березняке с осиной, примесью рябины и кустарниковых ив с преобладанием в травяном ярусе Athyrium filix-femina , Cirsium heterophyllum и Geranium sylvaticum . В непосредственной близости от цветоносных побегов надбород-ника обычно произрастали Gymnocarpium dryopteris , Filipendula ulmaria , Angelica sylves-tris , Rubus saxatilis и Geranium sylvaticum .
Морфологическое описание составлено на основе авторских наблюдений в 2001 – 2008 гг., а также по литературным данным.
Цветоносные побеги надбородника безлистного имеют высоту (5)10–20(30) см (Chung, Tzeng, 2001; Александров и др., 2004; Красная книга…, 2008; Taylor, Roberts, 2011).
Исследования показали, что подземные органы генеративных экземпляров размещались в верхних слоях почвы, на глубине от 2,5
до 5,5 см от поверхности опада, при толщине опада от 2 до 3 – 4 см. Три из четырех одиночных растений имели тонкий базальный кончик, т.е., вероятно, происходят из протокорма или протокормообразной почки. У четвёртого базальная часть была разрушена, что указывает на его больший (по меньшей мере год) возраст. Что касается генеративных побегов, собранных в группы, то только в одном случае от одного корневища отходили два генеративных побега; в остальных случаях каждый из 3 – 4 генеративных побегов группы имел в основании собственное корневище, причем их взаимное расположение и широкий край базальной части заставляли предположить, что эти растения появились путем партикуляции. Во время разбора подстилки также найдены восемь подземных вегетативных экземпляров разных размеров, образовавшихся из семян либо протокормообраз-ных почек. Вегетативные растения размещались в горизонтальной плоскости, одиночно или небольшими группами. Один из самых крупных подземных экземпляров был найден вплотную под корневищем небольшого генеративного побега; другой – между тремя одиночными генеративными экземплярами, на расстоянии от 10 до 30 см от них. Размещались корневища этих растений, как и у генеративных растений, в верхнем слое почвы. Более молодые подземные растения, размеры которых не превышали 1 см, располагались двумя группами – по 4 и 2 растения, на расстоянии 15 – 30 см от ближайших генеративных экземпляров. Обращает на себя внимание тот факт, что корневища этих молодых растений помещались не в почве, а в слое опада, между слаборазложившимися листьями древесных растений. Это позволяет с некоторой достоверностью утверждать, что возраст этих экземпляров – год или, что менее вероятно, два.
Считается, что после образования цветоноса корневище полностью или частично отмирает (Rasmussen, 1995; Александров и др., 2004; Красная книга…, 2008). Однако в 2007 г. была зафиксирована группа цветоносов надбородника, выросших внутри пластиково- го кольца диаметром около 7 см, которым мы окружили группу цветоносов в 2006 г. Это наблюдение косвенно указывает на сохранение корневища и после цветения.
Цветки надбородника довольно крупные (до полутора сантиметров от кончика гиностемия до цветоножки), располагаются по (1)2–4(8) в соцветии (Александров и др., 2004; Красная книга…, 2008). По нашим наблюдениям, высота цветоносных побегов варьирует от 5 до 32 см (в среднем 14±6 см); число цветков на цветоносном побеге колеблется от одного до пяти, чаще всего встречается по два цветка на побеге. Цветоножка изогнута, поэтому цветки направлены вниз. В экспериментах А. Кернера фон Марилауна (1906) цветки после того, как их переворачивали на 180 градусов, в течение 24 часов возвращались в нормальное положение. Губа цветка трёхлопастная, направлена вверх, довольно крупная и окрашена в светло-розовые тона с пурпурным точечным рисунком, на ней виден жидкостный блеск. На краевых участках внутренней стороны поперечного среза губы видны несколько разветвленных выростов, вершины которых составлены клетками с секреторной активностью (Rohrbach, 1866). P. Rohrbach, описавший эти клетки, затруднялся назвать выделяемые ими вещества. Для уверенного определения химической природы этих веществ необходимо провести гистохимические реакции in vivo . По отсутствию достаточно мощных проводящих пучков вблизи от выростов и рыхлой ткани, слагающей губу, можно предположить, что эти клетки выделяют слизеподобные вещества (М.Р. Леонтьева, биологический факультет МГУ, личное сообщение). Остальные пять лепестков желтоватые и узколанцетные; имеют одну жилку (Chung, Tzeng, 2001). Эти лепестки располагаются по бокам и в нижней части цветка. Гиностемий слегка изогнут. На нем так же, как и на губе, заметен жидкостный блеск. На верхней стороне гиностемия возле самого рыльца имеется светлое пятно — клеящее основание ножки поллиниев (Кернер фон Марилаун, 1906).
По нашим наблюдениям, цветки посещались исключительно двукрылыми насекомыми (табл. 1) и только в дневное время суток. Насекомые обычно садились на ярко окрашенную и блестящую сторону губы цветка и активно ползали по ней, собирая с неё нектар, реже зависали в воздухе перед цветком или садились на стебель и ползали по нему. Насекомые, севшие на цветок, взлетали чаще с гиностемия, сползая на него с губы, или – реже – взлетали с самой губы. Мы не зафиксировали ни одного случая взятия насекомыми пол-линиев, хотя у небольшой доли цветков, осмотренных в 2006 и 2007 гг. (9% и 2% от общего числа цветков соответственно), полли-нии отсутствовали. При анализе этих показателей необходимо учитывать, что во время исследований в 2007 г. цветение надбородни-ка только начиналось.
Таблица 1. Частота посещения цветков Epipogium aphyllum насекомыми разных семейств отряда Diptera
Frequency of different families of Diptera visits to flowers of Epipogium aphyllum
|
Семейство |
Частота посещений: число посещений (доля от общего числа посещений в этот год, %) |
||
|
2005 г. |
2006 г. |
||
|
Syrphidae |
Baccha elongata (Fabricius, 1775) |
2 (4) |
0 |
|
Cheilosia sp. |
1 (2) |
0 |
|
|
Не определено до рода |
14 (24) |
2 (11) |
|
|
Anthomyiidae |
3 (5) |
0 |
|
|
Muscidae |
3 (5) |
1 (6) |
|
|
Lauxaniidae |
самка Meiosimyza decipiens (Loew, 1847), или M. laeta (Zetterstedt, 1847) |
2 (4) |
0 |
|
Culicidae |
1 (2) |
1 (6) |
|
|
Sphaeroceridae |
1 (2) |
0 |
|
|
Не определено до семейства |
30 (52) |
14 (77) |
|
Динамика численности
НАДЗЕМНЫХ ПОБЕГОВ
Данные по численности генеративных побегов представлены в табл. 2 и обобщены на рис. 1. В 2001 г. при беглом осмотре найдено 34 генеративных побега (8 одиночных и 3 группы из 5 – 10 побегов). В 2002 г. при подробном осмотре обнаружено 68 генеративных побегов (9 одиночных и 6 групп из 4 – 20
Часть цветков надбородника поедается слизнями, что было отмечено и в других популяциях (Taylor, Roberts, 2011).
Число семязачатков в завязях составило от 1412 до 4665 (в среднем – 3210 ±1239) шт. Даже немногочисленный материал позволяет выявить зависимость числа семязачатков от положения цветка в соцветии. У некрупного растения с одиночным цветком завязь содержала 1412 семязачатков. Завязь первого (нижнего) и второго (среднего в трехцветковых соцветиях и верхнего в двухцветковом соцветии) цветков содержала сходное число семязачатков (в среднем 3997±206 и 4016±594 соответственно). У обоих растений с тремя цветками третий (верхний) цветок имел в среднем 1719±107 семязачатков.
побегов). В 2003 г. – 151 генеративный побег (4 одиночных и 4 группы из 4 – 70 побегов). В 2004 г. – 133 генеративных побега (7 групп из 2 – 48 побегов). В 2006 г. – 90 побегов (28 одиночных и 20 групп из 2 – 9 побегов). В 2007 г. – 138 побегов (2 одиночных и 19 групп из 2-21 побегов). В 2008 г. – 73 побега (14 одиночных и 16 групп из 2 – 11 побегов). В 2011 г. – 219 побегов (28 одиночных и 32 группы из нескольких побегов каждая).
Таблица 2. Число и высота генеративных побегов в популяции надбородника безлистного Epi-pogium aphyllum и число цветков, о. Асафий, Кемьлудский архипелаг, 2002 – 2008 гг.
Number and height of generative shoots and flower number in population of Epipogium aphyllum , Asafij island, Kemludsky archipelago, 2002 – 2008
|
Группа |
о в « о о 9 2 о к |
л 3 № о с |
Количество |
Группа |
о о к |
л 3 № о с |
Количество |
|||||
|
о м |
в Й |
Й СТ О |
в Й |
Й ь О |
||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
1 |
2 |
3 |
4 |
5 |
6 |
|
|
2002 г. 04.08.2002 |
6 |
2 |
13 |
3 |
||||||||
|
1 |
6 |
7 |
2 |
9 |
3 |
|||||||
|
2 |
2 |
7 |
2 |
12 |
3 |
|||||||
|
3 |
2 |
8 |
1 |
10 |
2 |
|||||||
|
4 |
2 |
9 |
2 |
15 |
3 |
|||||||
|
5 |
1 |
9 |
2 |
16 |
4 |
|||||||
|
6 |
10 |
10 |
1 |
10 |
1 |
|||||||
|
7 |
13 |
11 |
1 |
12 |
1 |
|||||||
|
8 |
6 |
12 |
1 |
11 |
1 |
|||||||
|
9 |
4 |
13 |
3 |
14 |
2 |
|||||||
|
10 |
2 |
13 |
3 |
15 |
2 |
|||||||
|
11 |
20 |
13 |
3 |
15 |
3 |
|||||||
|
2003 г.: 10.08.2003 (рис. 2) |
14 |
4 |
9 |
3 |
||||||||
|
1 |
10 |
14 |
4 |
11 |
2 |
|||||||
|
2 |
4 |
14 |
4 |
21 |
3 |
|||||||
|
3 |
2 |
14 |
4 |
25 |
3 |
|||||||
|
4 |
63 |
15 |
1 |
9 |
2 |
|||||||
|
5 |
1 |
16 |
2 |
12 |
1 |
|||||||
|
6 |
1 |
16 |
2 |
17 |
1 |
|||||||
|
7 |
70 |
17 |
4 |
13 |
2 |
|||||||
|
2004 г.: 10.08.2004 (рис. 2) |
17 |
4 |
15 |
2 |
||||||||
|
1 |
12 |
17 |
4 |
15 |
3 |
|||||||
|
2 |
19 |
17 |
4 |
15 |
2 |
|||||||
|
3 |
2 |
18 |
1 |
14 |
2 |
|||||||
|
4 |
14 |
19 |
3 |
13 |
2 |
|||||||
|
5 |
12 |
19 |
3 |
13 |
2 |
|||||||
|
6 |
48 |
19 |
3 |
14 |
3 |
|||||||
|
7 |
6 |
20 |
2 |
15 |
3 |
|||||||
|
2005 г.: Данных нет |
20 |
2 |
16 |
3 |
||||||||
|
2006 г.: 10–12.08.2006 |
21 |
1 |
16 |
2 |
||||||||
|
1 |
1 |
17 |
3 |
22 |
1 |
15 |
1 |
|||||
|
2 |
1 |
18 |
4 |
23 |
1 |
14 |
2 |
|||||
|
3 |
2 |
15 |
3 |
24 |
1 |
21 |
3 |
|||||
|
3 |
2 |
16 |
4 |
25 |
9 |
24 |
5 |
|||||
|
4 |
3 |
7 |
1 |
25 |
9 |
24 |
3 |
|||||
|
4 |
3 |
7 |
3 |
25 |
9 |
24 |
3 |
|||||
|
4 |
3 |
12 |
3 |
25 |
9 |
25 |
3 |
|||||
|
5 |
1 |
7 |
1 |
25 |
9 |
27 |
3 |
|||||
|
6 |
2 |
13 |
4 |
25 |
9 |
30 |
4 |
|||||
|
6 |
2 |
13 |
3 |
25 |
9 |
30 |
3 |
|||||
|
7 |
2 |
9 |
3 |
25 |
9 |
30 |
4 |
|||||
|
7 |
2 |
12 |
3 |
25 |
9 |
32 |
4 |
|||||
|
8 |
1 |
10 |
2 |
26 |
1 |
13 |
2 |
|||||
|
9 |
2 |
15 |
3 |
27 |
1 |
5 |
2 |
|||||
Продолжение таблицы 2
|
1 |
2 |
3 |
4 |
5 |
6 |
1 |
2 |
3 |
4 |
5 |
6 |
|
|
28 |
2 |
8 |
2 |
13 |
15 |
|||||||
|
28 |
2 |
9 |
2 |
14 |
1 |
|||||||
|
29 |
1 |
14 |
2 |
15 |
1 |
|||||||
|
30 |
1 |
31 |
4 |
16 |
14 |
|||||||
|
31 |
1 |
8 |
1 |
17 |
2 |
|||||||
|
32 |
1 |
6 |
1 |
18 |
1 |
|||||||
|
33 |
1 |
11 |
1 |
20 |
6 |
|||||||
|
34 |
1 |
11 |
2 |
21 |
3 |
|||||||
|
35 |
5 |
10 |
2 |
2008: 11.08.2008 |
||||||||
|
35 |
5 |
13 |
1 |
1 |
3 |
6 |
0 |
2 |
0 |
|||
|
35 |
5 |
13 |
3 |
1 |
3 |
12 |
0 |
2 |
0 |
|||
|
35 |
5 |
18 |
2 |
1 |
3 |
16 |
0 |
2 |
0 |
|||
|
35 |
5 |
21 |
3 |
2 |
3 |
9 |
0 |
2 |
0 |
|||
|
36 |
2 |
14 |
3 |
2 |
3 |
14 |
0 |
2 |
0 |
|||
|
36 |
2 |
25 |
3 |
2 |
3 |
19 |
0 |
2 |
0 |
|||
|
37 |
1 |
10 |
2 |
3 |
3 |
12 |
0 |
3 |
0 |
|||
|
38 |
1 |
20 |
3 |
3 |
3 |
17 |
0 |
2 |
0 |
|||
|
39 |
3 |
5 |
1 |
3 |
3 |
20 |
0 |
3 |
0 |
|||
|
39 |
3 |
6 |
1 |
4 |
1 |
6 |
0 |
2 |
0 |
|||
|
39 |
3 |
9 |
2 |
5 |
11 |
13 |
1 |
2 |
0 |
|||
|
40 |
3 |
21 |
3 |
5 |
11 |
11 |
0 |
2 |
0 |
|||
|
40 |
3 |
21 |
3 |
5 |
11 |
5 |
2 |
0 |
0 |
|||
|
40 |
3 |
22 |
3 |
5 |
11 |
14 |
1 |
2 |
0 |
|||
|
41 |
1 |
9 |
2 |
5 |
11 |
6 |
0 |
1 |
0 |
|||
|
42 |
4 |
10 |
2 |
5 |
11 |
5 |
1 |
0 |
0 |
|||
|
42 |
4 |
13 |
2 |
5 |
11 |
11 |
1 |
2 |
0 |
|||
|
42 |
4 |
14 |
2 |
5 |
11 |
9 |
2 |
0 |
0 |
|||
|
42 |
4 |
18 |
4 |
5 |
11 |
10 |
2 |
0 |
0 |
|||
|
43 |
1 |
15 |
1 |
5 |
11 |
5 |
3 |
0 |
0 |
|||
|
44 |
2 |
18 |
3 |
5 |
11 |
7 |
3 |
0 |
0 |
|||
|
44 |
2 |
19 |
3 |
6 |
1 |
9 |
0 |
1 |
0 |
|||
|
45 |
1 |
20 |
3 |
7 |
1 |
17 |
0 |
1 |
0 |
|||
|
46 |
2 |
12 |
1 |
8 |
3 |
6 |
1 |
1 |
0 |
|||
|
46 |
2 |
17 |
1 |
8 |
3 |
6 |
1 |
0 |
0 |
|||
|
47 |
1 |
11 |
2 |
8 |
3 |
7 |
1 |
0 |
0 |
|||
|
45 |
1 |
20 |
3 |
9 |
2 |
10 |
0 |
3 |
0 |
|||
|
46 |
2 |
12 |
1 |
9 |
2 |
10 |
1 |
0 |
0 |
|||
|
46 |
2 |
17 |
1 |
10 |
1 |
9 |
0 |
1 |
0 |
|||
|
47 |
1 |
11 |
2 |
11 |
1 |
19 |
0 |
3 |
0 |
|||
|
48 |
1 |
17 |
3 |
12 |
4 |
7 |
0 |
1 |
0 |
|||
|
2007: 29.07–01.08.2007 (рис. 3) |
12 |
4 |
7 |
0 |
0 |
0 |
||||||
|
1 |
2 |
12 |
4 |
11 |
0 |
4 |
0 |
|||||
|
2 |
21 |
12 |
4 |
6 |
0 |
2 |
0 |
|||||
|
3 |
8 |
13 |
4 |
8 |
2 |
1 |
0 |
|||||
|
4 |
10 |
13 |
4 |
5 |
0 |
0 |
||||||
|
5 |
1 |
13 |
4 |
8 |
1 |
0 |
||||||
|
6 |
7 |
13 |
4 |
6 |
0 |
0 |
||||||
|
7 |
13 |
14 |
1 |
10 |
1 |
0 |
0 |
|||||
|
8 |
3 |
15 |
3 |
21 |
1 |
3 |
0 |
|||||
|
9 |
10 |
15 |
3 |
16 |
1 |
2 |
0 |
|||||
|
10 |
2 |
15 |
3 |
17 |
2 |
1 |
0 |
|||||
|
11 |
9 |
16 |
1 |
22 |
0 |
4 |
0 |
|||||
|
12 |
3 |
17 |
1 |
11 |
0 |
2 |
0 |
|||||
Окончание таблицы 2
|
1 |
2 |
3 |
4 |
5 |
6 |
1 |
2 |
3 |
4 |
5 |
6 |
|
|
18 |
4 |
7 |
0 |
2 |
0 |
22 |
2 |
6 |
0 |
1 |
0 |
|
|
18 |
4 |
9 |
0 |
1 |
2 |
23 |
1 |
7 |
0 |
1 |
1 |
|
|
18 |
4 |
11 |
0 |
2 |
0 |
24 |
1 |
10 |
0 |
2 |
0 |
|
|
18 |
4 |
10 |
0 |
3 |
0 |
25 |
2 |
14 |
0 |
3 |
0 |
|
|
19 |
1 |
11 |
0 |
1 |
0 |
25 |
2 |
19 |
0 |
3 |
0 |
|
|
20 |
1 |
15 |
0 |
2 |
0 |
26 |
2 |
15 |
0 |
3 |
0 |
|
|
21 |
8 |
8 |
0 |
1 |
0 |
26 |
2 |
8 |
0 |
0 |
||
|
21 |
8 |
17 |
0 |
3 |
0 |
27 |
2 |
10 |
0 |
3 |
0 |
|
|
21 |
8 |
14 |
0 |
3 |
0 |
27 |
2 |
10 |
0 |
3 |
0 |
|
|
21 |
8 |
20 |
0 |
3 |
0 |
28 |
1 |
7 |
2 |
0 |
0 |
|
|
21 |
8 |
9 |
0 |
2 |
0 |
29 |
3 |
14 |
0 |
2 |
0 |
|
|
21 |
8 |
9 |
0 |
2 |
0 |
29 |
3 |
15 |
0 |
3 |
0 |
|
|
21 |
8 |
12 |
0 |
2 |
0 |
29 |
3 |
11 |
0 |
2 |
0 |
|
|
21 |
8 |
11 |
0 |
2 |
0 |
30 |
1 |
7 |
0 |
1 |
0 |
|
|
22 |
2 |
8 |
0 |
1 |
0 |
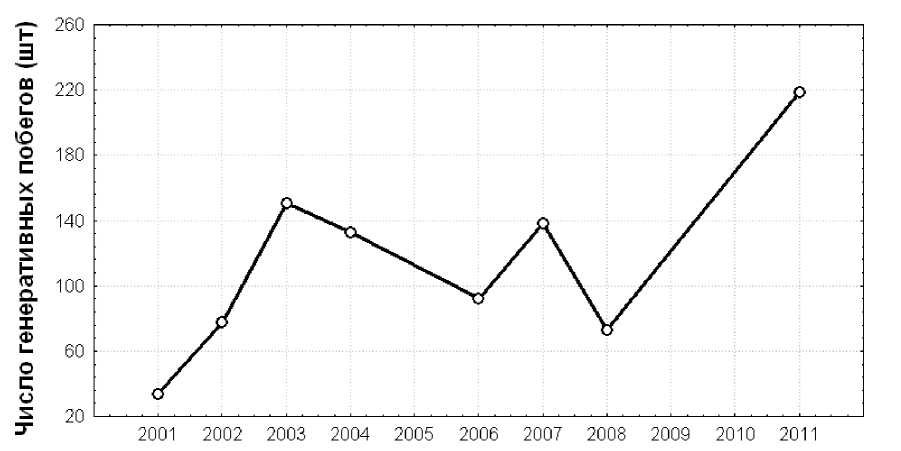
Год наблюдения
Рис. 1. Число генеративных побегов Epipogium aphyllum в разные годы Number of generative shoots of Epipogium aphyllum in different years
Как видно из приведенных данных, численность надземной части популяции увеличилась в 2002 г. по сравнению с 2001 г. и достигла максимума в 2003 г. В 2004 г. наблюдалось небольшое уменьшение общего числа генеративных побегов (по сравнению с 2003 г.) на фоне расширения площади надземной части популяции в северо-восточном направлении (рис. 2). При этом ее примерная площадь увеличилась вдвое (с 50 м2 до 100 м2). Повторное картирование надземной части популяции показало также, что местополо- жение генеративных побегов в 2003 г. и в 2004 г. различно (рис. 2). В 2006 г. число генеративных побегов стало еще меньше, чем в 2004 г. В 2007 г. отмечено значительное увеличение числа генеративных побегов почти до максимального значения, зарегистрированного в 2003 г., при этом площадь популяции составила около 1500 м2 (рис. 3). В 2008 г. численность генеративных побегов вновь заметно уменьшилась, а в 2011 г. их численность достигла максимального значения за все время наблюдений.
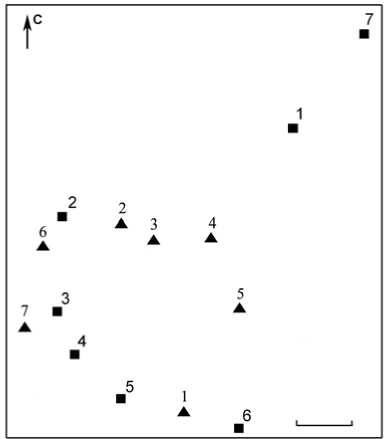
Рис. 3. Картосхема надземной части популяции Epipogium aphyl-lum на о. Асафий архипелага Кемь-Луды в 2007 г.
Map of the aboveground part of the Epipogium aphyllum population on the Asafij island (Kemludsky archipelago) in 2007
Условные обозначения. Длина масштабного отрезка – 2 м. Нумерация групп генеративных побегов соответствует табл. 2.
Рис. 2. Картосхема надземной части популяции Epipogium aphyllum на о. Асафий архипелага Кемь-Луды в 2003 – 2004 гг.
Map of the aboveground part of the Epipogium aphyllum population on the Asafij island (Kem-ludsky archipelago) in 2003 – 2004
Условные обозначения. Длина масштабного отрезка – 2 м. Форма символа, обозначающего группу генеративных побегов, соответствует году картирования: треугольник – 2003 г., квадрат – 2004 г. Нумерация групп генеративных побегов соответствует табл. 2.

Обсуждение результатов
Результаты морфометрии надбородника безлистного на острове Асафий согласуются с литературными данными (Chung, Tzeng, 2001; Александров и др., 2004; Красная книга…, 2008), а в некоторых случаях позволяют их дополнить.
Численность обнаруженных генеративных побегов в разные годы различалась в несколько раз, что позволяет предположить либо активное вегетативное или генеративное размножение, либо значительный «запас» нецветущих растений в почве. О подобном «запасе» говорят также изменения в расположении генеративных побегов в 2003, 2004 и 2007 гг., а также результаты исследования подземных органов надбородника. Отметим, что снижение числа надземных побегов не означает снижение численности растений в популяции. Кроме того, число обнаруженных побегов зависит и от «человеческого фактора» (интенсивности их поиска, которая в разные годы была неодинаковой).
Осуществленные наблюдения подтвердили данные А. Кернера фон Марилауна (1906) о наличии нектара в цветках надбородника. Этот факт говорит в пользу опыления насекомыми и согласуется с мнением, которое ранее высказал P. Rohrbach (1866). Наши собственные эксперименты показали, что поллинии могут быть отделены от цветка только при движении инородного тела сверху вниз. Такое движение насекомые могли бы совершить, переходя на гиностемий с губы. В то же время нами не было зафиксировано ни одного случая взятия насекомыми поллиниев. Это, вероятно, связано с тем, что насекомые улетали, не достигая клеящего основания поллиниев на гиностемии. Днем в пределах популяции надбородника летало множество шмелей, но ни один из них не пытался сесть на цветок надбородника. P. Rohrbach (1866), напротив, наблюдал посещение надбородни-ка шмелями. Возможно, потенциальные опылители надбородника отсутствовали среди насекомых, посетивших это растение во время наших наблюдений. Интересным представляется наличие завязавшихся семян во вскрытых нами завязях надбородника в первой декаде августа 2004 и 2006 гг., в связи с чем необходимо предположить наличие опылителей в районе исследований, учитывая невозможность самоопыления надбородника. В пользу этого свидетельствует отсутствие пол-линиев в некоторых цветках. Несмотря на выявленную значительную потенциальную семенную продуктивность, полученные данные не позволяют утверждать, что семенное возобновление играет значительную роль в поддержании популяции. Напротив, установлено, что наиболее крупные подземные экземпляры претерпевают партикуляцию перед образованием генеративного побега, а образование почек на столонах происходит достаточно регулярно. Однако отсутствие на нынешнем этапе исследований достоверных данных о проценте плодообразования не по- зволяет сделать окончательные выводы. В литературе имеются сведения о чрезвычайно низкой семенной продуктивности надбород-ника и о редкости плодоносящих растений в природе (Taylor, Roberts, 2011).
Полевые исследования проводили во время Беломорской экспедиции Московской гимназии на Юго-Западе (№ 1543), проходившей под руководством С.М. Глаголева, в рамках договора о сотрудничестве с Кандалакшским государственным природным заповедником. Авторы благодарят Л.А. Абрамову, П.Б. Борисову, Е.В. Елисееву и Д.А. Чудаева за помощь на полевом этапе исследования, С.А. Брискер и Е.А. Костереву за помощь в работе с собранным материалом по насекомым, Г.В. Фарафонову и А.И. Шаталкина за помощь в определении насекомых, К.А. Астафьева за изготовление срезов губы цветка, В.P. Филина и М.Р. Леонтьеву за консультации по морфологии и анатомии объекта исследований, а также В.Я. Бергера и В.И. Лю-ляева за помощь в решении транспортных проблем.
Список литературы Об обнаружении Тульского гербария барона Вальтера Вольдемаровича фон Розена
- Кожевников Д.А., Цингер В.Я. Очерк флоры Тульской губернии. Тр. Санкт-Петербургск. об-ва естествоисп., 1880, т. 11, вып. 1 [разд. пагин.], с. 37-150.
- Маевский П.Ф. Флора средней полосы европейской части России. Изд. 11-е, испр. и доп. М.: Тов-во науч. изданий КМК, 2014, 635 с.
- Назаров М.И. Каталог коллекций, поступивших в Гербарий Московского Государственного университета с 1813 по 1939 год. М., 1939, 228 с. [рукопись на каф. геоботаники МГУ]
- Перечень награжденных знаками отличия Русского географического общества (1845-2012). М.: Исполнительная дирекция РГО, 2012, 49 с.
- Розен В.В. Список растений, найденных в Тульской губернии до 1916 года. Изв. Тульск. об-ва любит. естествозн. Тула, 1916, вып. 4, с. 1-282, I-IV.
- Шереметьева И.С., Хорун Л.В., Щербаков А.В. Конспект флоры сосудистых растений Тульской области. М.: Изд-во Бот. сада Московск. ун-та; Тула: Гриф и К, 2008, 274 с.
