Об участии гетеротрофных микроорганизмов в начальном почвообразовании на отходах агломерации железных руд

Автор: Артамонова В.С., Булавина М.И.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 1, 2021 года.
Бесплатный доступ
Представлены результаты исследований развития гетеротрофных микроорганизмов в эмбриозёмах, формирующихся на песчаных отходах агломерации железных руд в Западной Сибири. На начальном этапе почвообразования - в инициальных эмбриозёмах обнаружены низкие значения актуальной кислотности и очень низкие показатели гумуса. Установлено, что такие среды обитания фитотоксичны, что подтверждают данные о всхожести семян и роста проростков: овса посевного, редьки масличной и горчицы белой. Доказано, что гетеротрофные микроорганизмы адаптированы к жизни в эмбриозёмах. Азотобактерии сохраняют жизнедеятельность, благодаря токсино- и слизеобразованию, и активно размножаются. Микромицеты проявляют диморфизм - дрожжевой и мицелиальный типы роста, что расширяет возможности их вегетативного размножения и сохранения популяции. Выявлено, что органо-аккумулятивные эмбриозёмы наиболее жизнепригодны для растений и микроорганизмов. Впервые зарегистрировано свечение плесневых грибов и азотобактера под злаковыми растениями и сосновыми насаждениями. Высказано предположение, что выброс световой энергии, присутствие оксидаз микроорганизмов и лигнина растений способствует гумификации в олиготрофной среде.
Бактерии, плесневые грибы, адаптация, почвообразование, техногенные отходы
Короткий адрес: https://sciup.org/147235444
IDR: 147235444 | УДК: 504.75+631.4 | DOI: 10.17072/1994-9952-2021-1-61-69
On the participation of heterotrophic microorganisms in initial soil formation on waste from iron ore agglomeration
The results of studies of the development of heterotrophic microorganisms in embryosemes formed on the sand waste of iron ore agglomeration in Western Siberia are presented. At the initial stage of soil formation, low values of actual acidity and very low humus values were found in the initial embryosemes. It is established that such habitats are phytiotoxic, which is confirmed by data on the germination of seeds and the growth of seedlings: oats, oilseed radish and white mustard. It is proved that heterotrophic microorganisms are adapted to life in embryos. Azotobacteria retain vital activity, thanks to toxin and mucus formation, and actively multiply. Micromycetes exhibit dimorphism-yeast and mycelial growth types, which expands the possibilities of their vegetative reproduction and population preservation. It was revealed that organo-accumulative embryos are the most viable for plants and microorganisms. For the first time, the glow of mold fungi and Azotobacter under cereal plants and pine plantations was recorded. It is suggested that the emission of light energy, the presence of oxidases of microorganisms and plant lignin contribute to humification in an oligotrophic environment.
Текст научной статьи Об участии гетеротрофных микроорганизмов в начальном почвообразовании на отходах агломерации железных руд
ганизмов, преимущественно местного происхождения, способных к детоксикации [Водолеев, Анд-роханов, Клековкин, 2007; Liste, 2009; Evaluation…, 2011]. Такие приёмы могут ускорить минерализационную и гумификационную активность в корнеобитаемом слое, улучшить минеральное питание растений, но доказательств о выживании и участии гетеротрофных микроорганизмов в этих процессах на песчаных токсичных железорудных отходах, недостаточно.
Любая почва возникает, функционирует и эволюционирует при участии и под частичным контролем биоты и в большой мере адаптирована к её нуждам [Макаров, 2007]. В зрелых почвах свыше 90% растительных остатков подвергается биодеградации с участием гетеротрофных микромицетов и бактерий [Кузнецов, Градова, 2006]. Они же участвуют в детоксикации металлов и гетеротрофном «гумусовом» цикле углерода. Показано, что микроорганизмы прижизненно метабилизируют органические остатки до низкомолекулярных соединений, а по мере гибели клеток внутри цитоплазмы происходят изменения, которые сопровождаются привлечением разрушающихся соединений в сборку первичных частиц гумусовых веществ (размером несколько нанометров) и затем – фрактальных кластеров или основных элементов организации гумусовой матрицы [Федотов, Лысак, Шалаев, 2013]. Источником гумусоподобных веществ могут быть меланины (прогуминовые или парагуминовые соединения) темноокрашенных микромицетов и азотобактерий, например род Azotobacter [Завгородняя, 2000; Попов, Зеленков, Теплякова, 2016]. Установлено, что азотобактер осуществляет продуцирование меланина лишь в присутствии бензойной кислоты [Gospodaryov, Lushchak, 2011] – продукта распада лигнина, который участвует в гумификации. Меланины грибного происхождения после деградации наиболее близки по элементному составу, молекулярным массам и оптическим свойствам с таковыми гуминовых веществ. Сообщается [Гесслер, Егорова, Белозерская, 2014], что в экстремальных условиях роста содержание меланинов у микромицетов может увеличиваться, обеспечивая им выживание в неблагоприятных условиях. Меланины выполняют адаптивную функцию, защищают микроорганизмы от повышенной радиации, УФ- и γ-излучения, тяжёлых металлов и активных форм кислорода, литического действия энзимов биоконкурентов за источники питания и энергии [Gospodaryov, Lushchak, 2011]. Меланизированные штаммы азотобактера выявлены ранее в антропогенно преобразованных почвах сибирского мегаполиса [Артамонова, 2002]. Информация о развитии неспорообразующих бактерий и несовершенных микро-мицетов в техногенных отходах железной руды на этапе инициирования в них почвообразования скудна. Имеются единичные сведения об адаптации гетеротрофных бактерий к условиям обитания на песчаных железорудных отходах [Артамонова и др., 2011].
Цель данной работы – изучить жизнедеятельность несовершенных микромицетов и неспорообразующих бактерий углеродной гетеротрофии в эмбриозёмах на песчаных отходах агломерации железной руды в условиях их самозарастания и лесной рекультивации.
Материалы и методы исследования
Объекты исследований представлены эмбри-озёмами инициальными и органо-аккумулятивными, формирующимися на отходах агломерации обогащения (обогащения) железосодержащей руды и её концентратов. Это хвосты магнитной сепарации рудных пород, которые представляют собой смесь измельчённой железосодержащей руды и технологических реагентов после обогатительного передела минерального сырья. Отходы гидротранспортом доставляются на полигоны – хвосто-хранилища, пребывают десятилетиями на открытом воздухе, представляя собой угрозу окружающей среде. В Кузбассе источником загрязнения окружающей среды являются отходы агломерации преимущественно магнетитовых руд (Fе 3 O 4 ), запасы которых составляют в регионе более 1.5 млрд т. Кроме железа они содержат сульфиды цинка, меди, а также кобальт, бор, золото. Они присутствуют в отвалах рудников и в отстойниках аглофабрик, одна из которых (Абагурская) находится в черте г. Новокузнецка. Её хвостохранилища являются причиной многолетнего загрязнения атмосферы города.
Хвостохранилище Абагурской обогатительной фабрики, с которого были отобраны пробы для почвенно-микробиологического анализа, действует с 1954 г., занимает к настоящему времени площадь около 35 га, где скопилось более 50 млн м3 отходов [Панова и др., 2017]. Хвосты – это мелкозернистые или пылеватые пески, зачастую очень токсичные и радиоактивные [Панова и др., 2017]. В их составе преобладают кислотные оксиды железа и остаточное магнитное железо, которое сосредоточено преимущественно в мелких фракциях. В процентном отношении лидируют SiO 2 , Fe 2 O 3 , CaO, Al 2 O 3 , FeO [Горбачёва, 2020]. Оксиды железа вместе с оксидами кремния и алюминия формируют кислые дренажи, загрязняющие почвы и водотоки прилегающих территорий. Из минералов присутствуют: Fe 3 O 4 ∙Fe 2 O 3 ∙FeO, каолинит, хлорид, вермикулит, полевые шпаты, биотит, кальцит, магнетит и карбонатные включения [Панова и др., 2017].
По истечении 30‒40 лет на поверхности мине- ральных отходов регистрируются локальные педо-подобные образования – эмбриозёмы. В них определялись: содержание углерода по Тюрину; актуальная кислотность традиционным в почвоведении методом; фитотоксичность – по угнетению растений, рекомендованных нормативными документами для тестирования токсичности. При проращивании семян овса, редьки и горчицы использовали 5-кратную повторность вместо 3-кратной, обеспечивающей снижение погрешности, обусловленной разбросом результатов. Микробиологические анализы – традиционные. Рост азотобактера и микро-мицетов учитывали на «голодном» агаре, для уточнения таксономической принадлежности мик-ромицетов использовали кислую среду Чапека. Антагонизм микроорганизмов выявлялся по регистрации литического действия, люминесценцию фиксировали с помощью инновационного продукта – ручки Invisible Ink en fnd Black Light. Данные статистически обрабатывали в программе Statistica.
Результаты и их обсуждение
Анализ содержания углерода в эмбриозёмах свидетельствует о дефиците органогена (таблица). Пересчёт углерода на гумус показал, что значения последнего соответствуют по общепринятой в почвоведении шкале Д.С. Орлова и Л.А. Гришиной очень низкому уровню (менее 2%).
|
Пул С (%) и кислотность в эмбриозёмах |
||||||
|
Проба |
Показатель |
M |
σ |
υ, % |
Доверительный интервал |
|
|
-95% |
+95% |
|||||
|
1 |
рН |
3.8 |
0.01 |
0 |
3.7 |
3.8 |
|
С, % |
0.3 |
0.01 |
4 |
0.3 |
0.3 |
|
|
2 |
рН |
3.8 |
0.02 |
1 |
3.7 |
3.8 |
|
С, % |
0.3 |
0.02 |
6 |
0.2 |
0.3 |
|
|
3 |
рН |
6.3 |
0.02 |
0 |
6.2 |
6.3 |
|
С, % |
1.0 |
0.02 |
2 |
1.0 |
1.0 |
|
|
4 |
рН |
5.7 |
0.02 |
0 |
5.7 |
5.8 |
|
С, % |
0.7 |
0.04 |
5 |
0.7 |
0.8 |
|
|
5 |
рН |
8.1 |
0.02 |
0 |
8.0 |
8.1 |
|
С, % |
1.2 |
0.11 |
9 |
0.9 |
1.5 |
|
|
6 |
рН |
8.0 |
0.03 |
0 |
8.0 |
8.1 |
|
С, % |
1.1 |
0.11 |
10 |
0.8 |
1.3 |
|
Примечание. 1, 2 – Э. инициальный; 3, 4 – Э. органо-акк. под злаками; 5, 6 – Э. органо-акк. под сосновыми насаждениями.
Минимальные значения гумуса обнаружены в инициальных эмбриозёмах без растений, чуть выше они оказались в органо-аккумулятивных молодых почвах, под злаками, донником и под сосновыми насаждениями. Актуальная кислотность эм-бриозёмов инициальных (рН < 4.5) соответствовала сильной кислотности, в органо-аккумулятивных – значения рН водного раствора (5.5‒6.0) приблизились к нейтральной степени. Сильная кислот- ность способствовала подвижности тяжёлых металлов и высокой фитотоксичности (рис. 1).
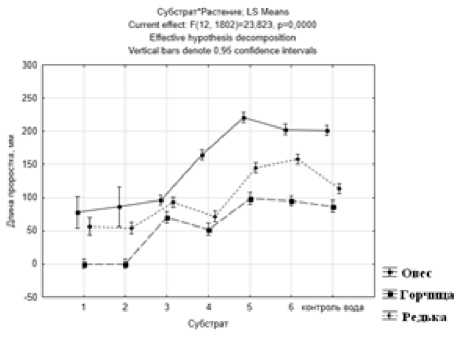
Рис. 1 . Длина проростков фитотестов:
1, 2 – Э. инициальный; 3, 4 – Э. органо-акк. под злаками; 5, 6 – Э. органо-акк. под сосновыми насаждениями
Наиболее устойчивым к таким условиям обитания оказался овёс. Для горчицы среда оказалась губительной. Следовательно, эмбриозёмы представляют собой олиготрофные среды обитания. Негативное влияние на накопление углерода мог оказывать песок в эмбриозёмах, поскольку в зрелых песчаных почвах гумус быстро минерализуется. Анализ гетеротрофных микробных ассоциаций показал, что в метаболически активном состоянии пребывают плесневые грибы р. Fusarium, Trichoderma, Penicillium и неспорообразующие грамотрицательные бактерии р. Azotobacter . Встречаемость азотобактерий (по степени обрастания мелкозёма) в инициальном эмбриозёме составляла 62–85%, в органо-акумулятивном – 100%, но меньшинство встречаемых бактерий в инициальном эмбриозёме оказались наиболее активными в процессе размножения. Их скорость роста в пересчёте на сутки на 30% была выше. В этом же типе эмбриозёма азотобактер продуцировал литические экзометаболиты против микро-мицетов (рис. 2). В органо-аккумулятивном эм-бриозёме они не наблюдались.
Мы полагаем, что углеродная гетеротрофия у азотобактера и плесневых грибов могла дополняться в условиях дефицита органической пищи древнейшим автотрофным путём усвоения углекислоты [Виноградский, 1952]. Хемосинтез органического вещества может проходить с участием углекислого газа, окиси углерода, муравьиной и уксусной кислот, метанола, карбонатов (в присутствии воды). Он обеспечивается за счёт энергии окисления неорганических соединений водорода, марганца, железа, серы, аммиака, а не энергии света. Среди хемосинтетиков присутствуют и азотобактерии, которые синтезируют энергию в виде молекулы АТФ. Такой метаболизм в эмбриозёмах исключать нельзя, но в нашем исследовании рост азотобактера обеспечивался на «голодной» питательной среде (Эшби), в рецептуре приготовления которой есть глюкоза (в небольших количествах); в её присутствии хемосинтез не происходит, поэтому корректнее говорить в нашем случае о гете-ротрофии.
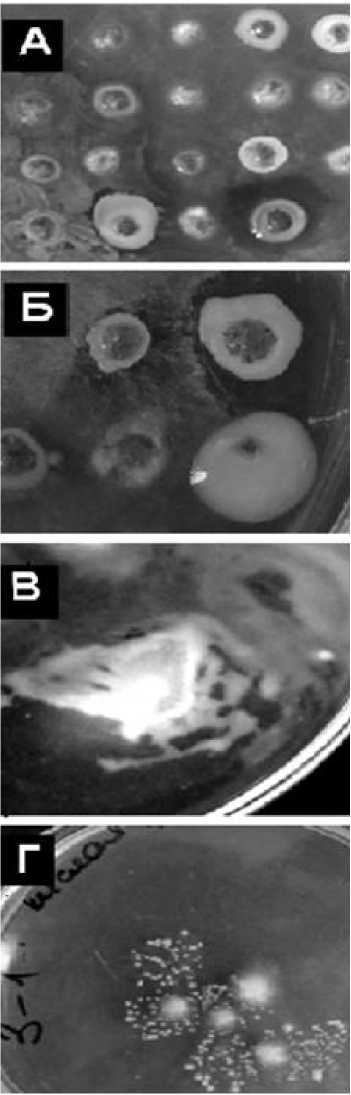
Рис. 2. Физиологические особенности микроорганизмов:
А – литические зоны азотобактера, Б – слизе-образование азотобактера, В – люминесценция азотобактера, Г -деморфизм микромицетов
Что касается проявления литической (антибиотической) активности Azotobacter chroococcum, то антифунгальный эффект был обнаружен ранее, в 60-е гг. прошлого века [Мишустин, Наумова, Хохлова, 1969]. Угнетать развитие микромицетов могли анисомицин группы азалидов [Придатчина и др., 1982], а также антибиотик фенольной группы – 5-алкил резорцин [Придатчина, 1984], способный подавлять рост Penicillium chrysogenum, Fusarium culmorum и простейших [Stasiuk, Kozubek, 2010]. Нужно отметить, что микромице-ты в такой обстановке проявляли диморфизм – наличие двух морфологических типов роста: дрожжевого и плесневого (гифального, мицелиального). Дрожжевой рост сопровождался почкованием, что наряду с фрагментами гиф, отдельных клеток, а также спор и конидий расширило возможности вегетативного размножения несовершенных грибов. Через плеоморфизм – фазовые изменения роста повышалась их численность бесполым путём. В органо-аккумулятивном эмбри-озёме такой адаптивной стратегии роста не наблюдалось.
Активной жизнедеятельности азотобактера способствовал не только синтез токсинов, но и слизеобразование. Слизью называют вещество, окружающее клетку, имеющее аморфный, бесструктурный вид и легко отделяющееся от поверхности клетки, по толщине зачастую превосходящее её диаметр. Поэтому основная масса находящихся в почвах органических полимеров микробного происхождения представлена внеклеточными полисахаридами и полиуронидами бактерий, или, иначе говоря, слизями и их остатками. Слизи азотобактера по химической природе являются полисахаридами, в составе которых доминирует ангидрид уроновой кислоты (до 75%).
Имеющиеся в составе полисахаридов карбоксильные и фенольные группы ответственны за распад кристаллических решеток минералов. Реагируя с определенными химическими элементами, они образуют комплексные связи, способствующие переходу элементов в раствор. Поэтому в эмбри-озёмах, где преобладают минеральные частицы, слизи – это важное биогенное средство извлечения элементов зольного питания. Одновременно слизи защищают клетки азотобактера от механических повреждений, иссушения, выедания простейшими, биодеградации другими микроорганизмами.
Слизи выступают также в роли резерва питательных веществ в ситуациях их дефицита, обезвреживания от экотоксикантов, в том числе путём хелатирования. В процессе жизнедеятельности азотобактер осуществляет сцепление песчаных частиц слизью, способствующих образованию педо-агрегатов. Следовательно, слизи – это мульти-функциональный адаптер сохранения жизнедеятельного состояния азотобактера в эмбриозёмах, биогенный фактор формирования водоудержива- ющей способности молодых почв на техногенных песчаных отходах. Проявлением другой адаптивной способности у азотобактера и плесневых грибов оказалась люминесценция их живых клеток голубым светом.
Люминесцентный путь превращения химической энергии в световую в условиях аэробиоза у грамотрицательных бактерий обязан окислительному метаболизму. Выброс энергии – это результат неутилизированной энергии и нейтрализации ферментами активных форм кислорода, токсичных (но необходимых) для процесса окисления глюкозы. Интересно, что голубое свечение микроорганизмов обусловлено возбуждением флавина, в результате окисления альдегида и восстановления молекулы рибофлавинфосфата. Провоцируют свечение активные формы кислорода, прежде всего, его реактивные соединения: синглетный (высокореактивный) кислород – радикал супероксид– аниона, водородный пероксид, гидроксильный анион. В настоящее время считается, что образование свободных радикалов является одним из патогенетических механизмов повреждения митохондрий, липидов, ДНК клетки [Коровина, Захарова, Обыночная, 2003]. Процесс перекисного окисления липидов в мембранах осуществляется по свободно радикальному механизму, подобно тому, как по цепному механизму происходит деление ядер урана [Коровина, Захарова, Обыночная, 2003]. Разрушаются жирорастворимые соединения, такие как убихинон – коэнзим СоQ8 у грамотрица-тельных бактерий, СоQ6 – дрожжей, поскольку они более уязвимы для процессов перекисного окисления. Кроме убихинона у Azotobacter chroococcum , могут поражаться, и другие липиды – их у бактерии не менее 16 [Алексеева, 2005]. Окислительным атакам могут подвергаться ферредоксины – белковые железосерные кластеры и флаводоксины – белковые структуры, идентичные ферредоксинам. Они участвуют в азотфиксации и их поражение может негативно отразиться на фиксации молекулярного азота азотобактером [Готтшалк, 1982]. Тем не менее, у представителей рода Azotobacter выработался особый механизм защиты от негативного действия кислорода — так называемая дыхательная защита. Она осуществляется путём значительной интенсификации дыхания, снижающего концентрацию кислорода в клетках [Берцова, Демин, Богачев, 2005]. Также имеется особый белок Shethna, защищающий нитрогеназу от кислорода [Maier, Moshiri, 2000]. Вероятно, такая защита способствовала выживанию и размножению азотобактера в эмбриозёмах.
Свободные радикалы, ведущие цепь окисления (обычно это перекисные радикалы), могут вступать в реакцию взаимодействия (рекомбинация или диспропорционирование) и в реакцию с молекула- ми клеточной мембраны, превращая их в свободные радикалы (самоподдерживающаяся лавинообразная реакция). Инициаторами таких взаимодействий являются химические реакции, связанные с изменением валентности иона металла (Cu2+, Fe2+). Особенно опасно для клетки одновременное присутствие О2- и Н2О2, причиной которого является нарушение эволюционно сформированных механизмов защиты микроорганизмов против токсичных форм кислорода – ферментов, способны предохранять клетки от «самозагрязнения» потоками активного кислорода и перекиси водорода путём каталитического расщепления [Хочачка, Сомеро, 1988]. Они имеют определенную специализацию как по отношению к конкретным видам радикалов и перекисей, так и по локусам возникновения активных форм кислорода. Активное ферментативное звено включает супероксиддисму-тазу, пероксидазу и каталазу. В этих ферментах важную роль имеет геминный компонент – протеиды с геминовой группой. Не исключено, что в эм-бриозёмах возникают ситуации, когда в микробных клетках возрастает не только пул свободных радикалов, но и повреждаются железосодержащие белки, их белковые комплексы с железом и серой под воздействием реакционных соединений азота – окиси азота и пероксинитрата. Известно, что окись азота производится практически всеми типами живых организмов, в том числе бактериями и микро-мицетами [Röszer, 2012]. Окись азота является высокореактивным свободным радикалом со временем жизни порядка нескольких секунд, но при этом обладает высокой способностью к проникновению сквозь биологические мембраны. Эндогенный оксид азота проявляет своё действие на клетки путём S-нитрозилирования тиоловых соединений (включая тиоловые группы серосодержащих аминокислот, таких, как цистеин) и нитрозилирования ионов переходных металлов, которых в эмбриозё-мах достаточно. Установлено [Park, 2003; Imlay, Korshunov, Imlay, 2015], что цистеин вследствие его высокой реакционной способности может восстанавливать внутриклеточное железо, способствуя протеканию реакции окисления пероксида водорода – H2O2 с ионами железа (реакция Фентона), которая сопровождается образованием токсичных гидроксильных радикалов и индукцией хемолюминесценции. Интенсивность свечения, возникающего при рекомбинации свободных радикалов, пропорциональна квадрату их концентрации в системе. В экспериментах с биологическими субстратами и органическими веществами показатель светосуммы люминесценции, индуцированной реакцией Фентона, стабилен при концентрации компонентов [Fe2+] = 10-3 моль/л, [H2O2] = 10-3 моль/л, pH = 2 [Иванова, Трофимова, Пискарёв, 2014]. Такие условия реальны в инициальных эм- бриозёмах. Не исключено также связывание окиси азота не только с Fe, но и с Cu, Zn, Cr, Co, Mn, ионами переходных металлов в составе простети-ческих групп и активных каталитических центров металлоферментов, дефицита которых в эмбри-озёмах тоже нет, они наследуются от отходов железной руды. Интересно другое – выявленное нами свечение регистрировалось у микроорганизмов, обитающих преимущественно под злаками и сосновыми посадками. Эти растения являются основными продуцентами лигнина в растительном мире. Продукт его разложения – бензойная кислота – является важнейшим компонентом гумификации растительного опада хвойных и лиственных растительных сообществ [Анохина и др., 2018]. Окисление остатков лигнина и его связь с белками микроорганизмов в слабощелочной среде обычно сопровождаются образованием гуминовых кислот. В окислении лигнина участвуют лигнолитические микромицеты, благодаря присутствию ферментов – оксидаз, таких как марганецпероксидаза и лиг-нинпероксидаза. В качестве кофакторов окисления органических остатков у них выступают гемы Fe, Cu, Mn, обеспечивающие разрушение перекиси водорода до молекулярного кислорода и воды [Дармов, Горшунова, Тарасова, 2017]. Присутствие металлов в таких местообитаниях не оспаривается, поскольку стратегия растений поглощать тяжёлые металлы связана с выделением корнями производных мугеиновых кислот, действующих как внешние фитосидерофоры, мобилизующие нерастворимые ионы металлов [Murakami et al., 1989; Takagi, 1993]. Следовательно, молекулярный кислород – продукт каталитического расщепления геминными ферментами быстро вовлекается в окисление железа, и процесс образования свободных радикалов в микробных клетках продолжается. Свечение способствует освобождению невостребованной энергии микробной клеткой и частичной нейтрализации радикалов, тем самым предохраняет микроорганизмы от развития митохондриальной дисфункции, пероксидации липидов, нарушения ДНК и нитрогеназы. Жизнедеятельные клетки являются источником микробной биомассы и участником гумификации, поскольку выброс энергии, синтез оксидаз в присутствии лигнина способствует образованию гуминовых кислот.
Заключение
Эмбриозёмы, формирующиеся на отходах обогащения железной руды, жизнепригодны для гетеротрофных неспорообразующих бактерий и несовершенных микромицетов. На инициальном этапе педогенеза среда обитания микроорганизмов характеризуется сильной кислотностью, олиготроф-ностью, токсичностью, присутствием оксидного и магнитного железа. Жизнедеятельные микроорга- низмы обнаружили к ним различные способы адаптации: азотобактерии выживают, благодаря токсино- и слизеобразованию, микромицеты – благодаря диморфизму – дрожжевому и мицелиальному росту, что расширяет возможности их вегетативного размножения и сохранения популяции. В нейтральной среде обитания – эмбриозёмах органо-аккумулятивных разнообразие типов роста у микромицетов отсутствовало. Специфическим способом адаптации микромицетов и азотобактера к нарушению окислительных реакций, обусловленных присутствием железа и других металлов, оказалась люминисценция. Освобождение клетками неиспользованной энергии в форме света, предохраняло их от гиперкоагуляции белков, перекисного окисления липидов, гипергликемии, которые вызываются перекисными радикалами и активными формами кислорода. Обычно освобождение химической энергии, не вовлекаемой в процессы метаболизма, наблюдается у микромицетов и бактерий при термодинамически полном окислении, но за пределы клетки энергия освобождается в форме тепла, но не в форме света. В таком случае бактерии теряют около половины продуцируемой энергии, плесневые грибы – значительно больше, поскольку используют для синтетических целей лишь около 10% энергии. В патологическом состоянии – при интенсивном окислении кислорода в присутствии избытка свободных радикалов, энергия выбрасывается фотонами с максимумом излучения бактериями 470‒500 нм, что достаточно для возбуждения свечения в видимой части спектра. Испускаемая энергия – 70 ккал/1 эйнштейн значительно превышает энергию большинства биохимических реакций, например, при распаде высокоэнергетической молекулы АТФ (7 ккал) [Биолюминесценция, 2020]. Такой энергетический приток в олиготрофную среду способствует, на наш взгляд, протеканию химических и автокаталитических процессов окислительного характера вокруг клетки для её же блага. Освобождённая энергия, оксидазы погибших микроорганизмов и присутствие лигнина, очевидно, способствуют гумификации в олиготрофной песчаной молодой почве, что при необходимости микроорганизмы могут использовать в качестве углеродного ресурса. Полученная информация о жизнедеятельных гетеротрофных микроорганизмах в эмбриозёмах, формирующихся на песчаных полигонах железорудных отходов, расширяет знания об экологии микроорганизмов и их участии в современном педогенезе.
Работа выполнена в рамках государственного задания ИПА СО РАН по бюджетному финансированию Министерства науки и образования Российской Федерации.
Список литературы Об участии гетеротрофных микроорганизмов в начальном почвообразовании на отходах агломерации железных руд
- Алексеева А.Е. Физиолого-биохимическая активность и разнообразие штаммов Azotobacter chroococcum, выделенных из почв Нижегородской области: дис. ... канд. биол. наук. Н. Новгород, 2005. 141 с.
- Анохина Н.А. и др. Динамика содержания ароматических кислот в биогеоценозах стационарных почвенных лизиметров // Вестник Московского университета. Сер. 17. Почвоведение. 2018. №4. С. 3-10.
- Артамонова В.С. Микробиологические особенности антропогенно преобразованных почв Западной Сибири. Новосибирск: Изд-во СО РАН, 2002. 225 с.
- Артамонова В.С. и др. Эколого- микробиологическое разнообразие микробных сообществ в техногенно-нарушенных ландшафтах Сибири // Сибирский экологический журнал. 2011. Вып. 5. С. 735-746.
- Берцова Ю.В., Демин О.В., Богачев А.В. Дыхательная защита нитрогеназного комплекса у Azotobacter vinelandii // Успехи биологической химии. 2005. Т. 45. С. 205-234.
- Биолюминесценция. 2020. [Электронный ресурс]. URL: https://www.bibliofond.ru/ view.aspx?id= 56452 (дата обращения: 19.12.2020)
- Виноградский С.Н. Микробиология почвы. Проблемы и методы. М.: Изд-во АН СССР, 1952. 792 с.
- Водолеев А.С., Андроханов В.А., Клековкин С.Ю. Почвоулучшители: рекультивационный аспект. Новосибирск: Наука, 2007. 148 с.
- Гесслер Н.Н., Егорова А.С., Белозерская Т.А. Экстремальных условиях существования (обзор) // Прикладная биохимия и микробиология. 2014. Т. 50, № 2. С. 125-134.
- Горбачёва К. Стратегия переработки отходов обогащения железных руд Мундыбашской обогатительной и Абагурской агломерационно-обогатительных фабрик. [Электронный ресурс]. URL: https://pandia.ru/text/77/194/28780/php (дата обращения: 26.10.2020).
- Готтшалк Г. Метаболизм бактерий. М.: Мир, 1982. 310 с.
- Дармов И.В., Горшунова Е.И., Тарасова Т.С. Исследование природных изолятов микромицетов Fusarium spp. - продуцентов лигнолитических ферментов // Учёные записки Казанского университета. Сер. Естественные науки. 2017. Т. 159, кн. 1. С. 72-84.
- Завгородняя Ю.А. Сравнительная характеристика гуминовых кислот и грибных меланинов: дис. ... канд. биол. наук. М., 2000. 109 с.
- Иванова Л.П., Трофимова С.В., Пискарёв И.М. Хемилюминесценция, индуцированная реакцией Фентона, - математическое моделирование процесса; особенности, параметры и условия применения для биомедицинских исследований // Современные технологии в медицине. 2014. Т. 6, № 4. С. 14-25.
- Коровина Н.А., Захарова И.Н., Обыночная Е.Г. Применение антиоксидантов в педиатрической практике // Сonsilium-medicum. 2003. Т. 5, № 9. С. 47-52.
- Кузнецов А.Е., Градова Н.Б. Научные основы эко-биотехнологии. М.: Мир, 2006. 504 с.
- Макаров И.Б. Плодородие и продуктивность почв: соотношение понятий // Плодородие. 2007. № 3. С. 33-35.
- Мишустин Е.Н., Наумова А.Н., Хохлова Ю.М. Ан-тифунгальный антибиотик из культуры Azotobacter chroococcum // Микробиология. 1969. Т. 39, вып. 1. С. 87-90.
- Панова В.Ф. и др. Переработка отходов обогащения железной руды // Вестник Сибирского государственного индустриального университета. 2017. № 3 (21). С. 56-62.
- Попов А.И., Зеленков В.Н., Теплякова Т.В. Биологическая активность и биохимия гуминовых веществ. Ч. 1. Биохимический аспект (обзор литературы) // Вестник Российской Академии естественных наук. 2016. № 1. С. 11-18.
- Придатчина Н.Н. Биологически активные вещества из клеточных липидов азотфиксирующей бактерии Azotobacter chroococcum: дис. ... канд. биол. наук. М., 1984. 144 с.
- Придатчина Н.Н. и др. Azotobacter chroococcum -продуцент нового противогрибкового антибиотика // Антибиотики. 1982. № 1. С. 3-5.
- Федотов Г.Н., Лысак Л.В., Шалаев В.С. Микроорганизмы и образование гумусовых веществ в почвах // Лесной вестник. 2013. № 7. С. 111-115.
- Хочачка П., Сомеро Дж. Биохимическая адаптация. М.: Мир. 1988. 568 с.
- Benabdellah K. et al. GintPDXl encodes a protein in volved in vitamin B6 biosynthesis that is up-regulated by oxidative stress in the arbuscular my-corrhizal fungus Glomus intraradices // New Phy-tol. 2009. Vol. 184. P. 682-693.
- Evaluation of Urban Soils: Suitability for green infrastructure or urban infrastructure. 2011. EPA Publication. 20 p. [Электронный ресурс]. URL: https://www. epa. gov/sites/production/files/2015-10/documents/evaluation-of-urban-soils.pdf (дата обращения: 18.12.2020).
- Gospodaryov D., Lushchak V. Some properties of melanin produced by Azotobacter chroococcum and its possible application in biotechnology // Бютехнолопя. 2011. Т. 4, № 2. C. 61-69.
- Imlay K.R.C., Korshunov S., Imlay J.A. Physiological roles and adverse effects of the two cystine importers of Escherichia coli // J. Bacteriol. 2015. Vol. 197 (23). P. 3629-3644.
- Liste H.-H. Auswahl und Konditionierung alternativer Pflansubstrate zur Rekultivverung von Deponien und Altablagerungen // Обеспечение безопасности закрытых полигонов твёрдых бытовых отходов экологическими методами: материалы междунар. семинара (7-13 сент. 2009 г. Пермь, ПГУ). Пермь; Берлин; М., 2009. С.69-78.
- Maier R.J., Moshiri F. Role of the Azotobacter vine-landii Nitrogenase-Protective Shethna Protein in Preventing Oxygen-Mediated Cell Death // J. Bac-teriol. 2000. Vol. 182, № 13. Р. 3854-3857.
- Murakami T. et al. Stabilities of metal complexes of mugeinic acids and their specific affinities for iron (III) // Chem. Lett. 1989. P. 2137-2140.
- Park S. High levels of intracellular cysteine promote oxidative DNA damage by driving the Fenton reaction // J. Bacteriol. 2003. Vol. 185(6). P. 19421950.
- Roszer T. Nitric Oxide is a Bioproduct in Prokaryotes // The Biology of Subcellular Nitric Oxide. Springer Science+Business Media. 2012. Vol 10, № 2. P. 19-46.
- StasiukM., KozubekA. Biological activity of phenolic lipids // Cell. Mol. Life Sci. 2010. Vol. 67. P. 841-860.
- Takagi S. Production of phytosiderophores // Iron Chelation in Plants and Soil Microorganisms. New York: Academic Press, 1993. P. 111-131.
