Обобщение фаунистических материалов о совкообразных чешуекрылых (Lepidoptera, Noctuoidea: Erebidae, Nolidae, Noctuidae) Прибайкалья и Западного Забайкалья (Бурятия)
 Прибайкалья и Западного Забайкалья (Бурятия)")
Автор: Гордеев С.Ю., Гордеева Т.В.
Журнал: Природа Внутренней Азии @nature-inner-asia
Рубрика: Биология
Статья в выпуске: 1 (34), 2026 года.
Бесплатный доступ
На территории Прибайкалья и Западного Забайкалья (Бурятия) достоверно установлено 477 видов бабочек из семейств Erebidae, Nolidae, Noctuidae. Максимальное видовое богатство отмечено в лесостепном (285 видов) и нижнетаежном (248 видов) поясах, в то время как наибольшая доля узкоспециализированных видов характерна для гольцового (20,5 %) и степного (16,9 % поясов. По широтной составляющей ареала ядро фауны (65,2 %) формируют суббореальные виды. По долготной структуре ареала преобладают транспалеарктические виды. Фауна имеет смешанный характер с выраженными западными (евросибирскими, центральнопалеарктическими) и восточными (восточнопалеарктическими, центрально-восточнопалеарктическими, южносибирско-палеархеарктическими) элементами. Выделено четыре основных экологических комплекса: лугово-степной, лесной, влажно-прибрежный и высокогорный. Значительна доля локальных (64 %) и редких (44 %) видов, что указывает на высокую специализацию и уязвимость фауны совок.
Чешуекрылые, совки, Noctuoidea, Прибайкалье, Западное Забайкалье, Бурятия, хорология, высотная поясность, биотопическое распределение
Короткий адрес: https://sciup.org/148333175
IDR: 148333175 | УДК: 595.786 | DOI: 10.18101/2542-0623-2026-1-6-36
Synthesis of Faunistic Data on Lepidoptera (Noctuoidea: Erebidae, Nolidae, Noctuidae) from Cisbaikalia and Western Transbaikalia (Buryatia)
477 species of Lepidoptera, Noctuoidea (Erebidae, Nolidae, Noctuidae) have been reliably recorded in Cisbaikalia and Western Transbaikalia (Buryatia). The maximum species richness is noted in forest-steppe (285 species) and lower taiga (248 species) belts, the largest proportion of highly specialized species is characteristic of goltsy (20.5 %) and steppe (16.9 %) belts. In terms of latitudinal distribution, the core of the fauna (65.2 %) is formed by subboreal species. Regarding the longitudinal structure of the ranges, transpalearctic species predominate. The fauna is of a mixed nature with pronounced western (Euro- Siberian, Central Palearctic) and eastern (East Palearctic, Central-East Palearctic, South Siberian-Palaeoarchearctic) elements. Four main ecological complexes have been identified: meadow-steppe, forest, wetland coastal, and high-mountain. The proportion of local (64 %) and rare (44 %) species is exceptionally high, indicating a high degree of specialization and vulnerability of the fauna.
Текст научной статьи Обобщение фаунистических материалов о совкообразных чешуекрылых (Lepidoptera, Noctuoidea: Erebidae, Nolidae, Noctuidae) Прибайкалья и Западного Забайкалья (Бурятия)
Настоящая работа — продолжение серии статей о совках (совкообразных) — ноктуоидных чешуекрылых Прибайкалья и Западного Забайкалья [Гордеев и др., 2019, 2022; Гордеев, Гордеева, 2024]. Основой послужил материал коллекционного фонда лаборатории систематики и экологии животных (ЛСиЭЖ) ИОЭБ СО РАН (г. Улан-Удэ), источники литературы [Кононенко, 1990; Рудых, Власова, 1997; Матов, Белова, 2016; Махов и др., 2018; Каталог чешуекрылых России, 2019; Махов, 2021; Gordeev, Gordeeva, 2021; Kononenko, 2010, 2016]. История исследований региона, методика сбора, хранения и идентификации материала описана ранее [Гордеев и др., 2019, 2022].
Основная цель: обобщение фаунистических данных о Noctuoidea семейств Erebidae, Nolidae, Noctuidae Прибайкалья и Западного Забайкалья, их хорологии, высотной поясности, биотопическом распределении.
Термины и определения, перечень сокращений и обозначений
Совкообразные чешуекрылые (Noctuoidea) — наиболее многочисленная группа высших ночных чешуекрылых. В настоящей работе рассмотрено три семейства Noctuoidea: Erebidae, Nolidae, Noctuidae [Каталог чешуекрылых России, 2019]. Остальные встречающиеся в Бурятии Noctuoidea: Notodontidae, Lymantriidae, Arctiidae — проанализированы ранее [Шодотова и др., 2007].
Фауна Lepidoptera рассмотрена традиционно в следующих аспектах: трехмерная структура ареала (широтно-высотный (или зонально-поясный), долготный) [Городков, 1984; Михайлов, 2018]; высотное распределение видов; биотопическая их приуроченность. Кроме того, проведена оценка встречаемости видов в регионе.
По широтной составляющей выделены следующие группы ареалов видов:
АА — Арктоальпийская: занимающие тундру, лесотундру в приполярных районах; горные тундры и альпийские луга — в бореальной зоне.
АБ — Арктобореальная: распространенные в бореальной (таежной) зоне и лесотундре; в горах — таежном поясе и подгольцовье.
Б — Бореальная: приуроченные к бореальному поясу (зона тайги — хвойных лесов), подпоясу лиственных лесов, лесостепи.
СБ — Суббореальная: распространенные в суббореальных широколиственных лесах; степи; лесостепи.
ТМ — Температная (умеренная): придерживающиеся бореального и суббореального поясов.
ПЗ — Полизональная: распространенные от лесотундры до степи.
По долготной составляющей ареала взяты за основу следующие типы:
К — Космополитическая: обитающие в северной и южной частях земного шара.
ГА — Голарктическая: распространенные в Палеарктике (Старый Свет) и Не-арктике (Новый Свет).
ТП — Транспалеарктическая: обитающие в Палеарктике от берегов Атлантического (порой включая Северную Африку) до Тихого океана.
ЕС — Евросибирская: распространенные от Западной/Северной Европы до Байкала / восточных границ Забайкалья и Центральной Якутии.
ЦП — Центральнопалеарктическая: распространенные в центральной части Палеарктики. Сюда отнесены виды с сибирскими, южносибирско-китайскими, сибирско-монгольскими, восточносибирско-монгольскими, южносибирско-мон-гольскими, южно-уральско-южносибирско-монгольскими, среднеазиатско-южносибирскими, забайкальско-монгольско-китайскими ареалами.
ЮП — Южносибирско-палеархеарктическая: распространенные в южной гумидной и частично в аридной части Восточной Палеарктики, от гор Южной Сибири до Приморья. Сюда включены виды забайкальско-палеархеарктического и южносибирско-палеархеарктического комплексов.
ЦВП — Центрально-восточнопалеарктическая: распространенные в восточной части Палеарктики — от Приморья до Уральских гор.
ВП — Восточнопалеарктическая: распространенные от Восточной Сибири до Тихого океана, в том числе его островов.
АП — Амфипалеарктическая: с разорванными ареалами, обитающие в при-океанических районах Евразии.
При характеристике высотного распределения видов выделены следующие высотно-растительные пояса:
СТ — степной пояс.
ЛС — лесостепной пояс.
НТ — нижнетаежный пояс, или пояс мелколиственных и смешанных лесов.
ВТ — верхнетаежный пояс, или пояс хвойных, главным образом лиственничных и кедровых, лесов оптимального развития.
ПГ — пояс подгольцовий, или хвойных редколесий угнетенного развития.
ГЦ — гольцовый пояс, или пояс высокогорных тундр.
При оценке встречаемости видов в Западном Забайкалье и Прибайкалье задано два параметра: Р — редкий, О — обычный. Для видов, населяющих небольшие по площади участки, приводится термин «локальный» (Л).
Обычный — вид, встречающийся каждый год в пределах свойственного ему в регионе местообитания с численностью не менее одного-трех экземпляров за учетное время: на маршрутах протяженностью 3–5 километров (для видов, активных днем); за 3–7 ловушко/часов (для ночных видов).
Редкий — встречающийся в отдельные годы с численностью один и менее экземпляров на суммарных маршрутах 10–30 км или за 20 и более суммарных ловушко/часов.
Для выявления оригинальности фаун высотных поясов применен коэффициент оригинальности (Ko) [Дубатолов и др., 2004].
n а1 1 N — число видов в выделе;
— ri 11-1 n — количество выделов в рассматриваемом регионе;
N ai — количество выделов, где встречается i-й вид.
Ниже перечислены основные биотопы (местообитания), занимаемые видами Erebidae, Nolidae, Noctuidae: луг (лг); высокогорный луг (вл); горные каменистые россыпи (гкр); болото (бт); тундра (тд); степь (ст); ильмовники (ил); все типы лесов (втл); влажные леса (влл); разреженные леса (рл); лес темнохвойный (лт); лес светлохвойный (лс); лес лиственный и/или смешанный (ллс); кустарниковые заросли (кз); аграрные, в том числе застроенные, распаханные (агр).
Все известные нам виды совок Прибайкалья и Западного Забайкалья рассмотрены по высотному распределению, широтной и долготной составляющим их ареалов, биотопической приуроченности, оценена степень их редкости (редкий, обычный, локальный) (табл. 7).
Результаты и обсуждение
Высотное распределение совок
Анализ высотного распределения видов Erebidae, Nolidae, Noctuidae достаточно четко отражает классический принцип смены фаун по высотным поясам — от богатого и разнородного видового комплекса в нижних, более благоприятных поясах (лесостепном и нижнетаежном) к специфичному, обедненному и узкоспециализированному комплексу в высокогорьях — подгольцовом и гольцовом поясах (табл. 1, рис. 1).
Степной пояс. Характеризуется наибольшей долей стенотопных видов, приуроченных к открытым аридным местообитаниям. Типичные представители этого пояса — виды родов Cucullia sp., Eublemma sp., Drasteria sp., Callistege sp. Здесь преобладают виды, чьи гусеницы связаны с травянистой растительностью.
Лесостепной пояс. Здесь отмечено максимальное видовое богатство, а также высокая доля эвритопных видов. Типичные представители — виды родов Hypena sp., Zanclognatha sp., Polypogon sp., Catocala sp. Это «коридор», где соприкасаются фауны степей и лесов. Высокая мозаичность местообитаний способствует здесь высокому видовому разнообразию.
Верхнетаежный пояс (темнохвойная тайга). Отличается преобладанием типично лесных, часто влаголюбивых видов, связанных с хвойными породами деревьев. Типичные представители: виды родов Syngrapha sp., Xestia sp., Polia sp., Panolis flammea . Здесь исчезают степные и многие лесостепные виды.
Подгольцовый пояс (редколесья). Отмечено резкое обеднение видового состава, появление видов, характерных для высокогорий и северных тундр. Типичные представители — Syngrapha parilis, Anarta melanopa, Xestia tecta, Lasionycta sp. Доминируют виды с голарктическими и арктоальпийскими ареалами, устойчивые к суровым условиям.
Гольцовый пояс (высокогорные тундры). Характеризуется наиболее оригинальной и обедненной фауной. Типичные представители — Sympistis zetterstedti, Polia richardsoni, Xestia quieta . Отмеченные здесь виды часто имеют островные ареалы в аналогичных биотопах других горных систем или в Арктике. Связаны они все с каменистыми россыпями, горными тундрами и альпийскими лугами.
Таблица 1
Виды совок, отмеченные в разных высотных поясах
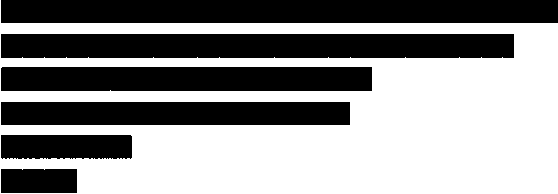
Рис. 1. Распределение видов совок по высотным поясам
|
Высотный пояс |
Количество видов |
Процент от общего числа |
|
СТ |
166 |
34,8 |
|
ЛС |
285 |
59,7 |
|
НТ |
248 |
52,0 |
|
ВТ |
151 |
31,7 |
|
ПГ |
67 |
14,0 |
|
ГЦ |
44 |
9,2 |
Лесостепной (285) Нижнетаёжный (248) Степной (166)
Верхнетаёжный (151) Подгольцовый (67) Гольцовый (44)
Анализ оригинальности фаун совок высотных поясов ожидаемо показал, что наиболее высок коэффициент оригинальности гольцового пояса (табл. 2). Каждый пятый вид, найденный здесь в тундрах, является уникальным и нигде больше не встречается. Достаточно высокая оригинальность у степного пояса. Около 17% степных видов не отмечено больше нигде, то есть степная фауна совок самобытна. Средние значения у подгольцового и верхнетаежного поясов. Здесь есть свои уникальные виды, адаптированные к местным условиям, вместе с тем много видов, проникающих сюда из соседних поясов. Минимальная оригинальность у нижнетаежного и лесостепного поясов.
Таблица 2
Коэффициент оригинальности фауны совок по высотным поясам
|
Высотный пояс |
Общее число |
Коэффициент оригинальности |
|
|
видов в поясе |
видов-уникумов |
||
|
СТ |
166 |
28 |
0,169 (16,9%) |
|
ЛС |
285 |
— |
— |
|
НТ |
248 |
2 |
0,008 (0,8%) |
|
ВТ |
151 |
8 |
0,053 (5,3%) |
|
ПГ |
67 |
8 |
0,119(11,9%) |
|
ГЦ |
44 |
9 |
0,205 (20,5%) |
Таким образом, лесостепной и нижнетаежный пояса — фаунистические «коридоры», или «зоны смешения». Наряду с наличием здесь максимального видового богатства (285 и 248 видов) их фауна почти полностью состоит из видов, проникающих сюда из соседних поясов (степей, темнохвойной тайги). Наибольшая уникальность присуща крайним поясам — гольцовому и степному. Это доказывает экологический принцип — максимальное число видов наблюдается в условиях средней жесткости и высокой мозаичности среды (лесостепной, нижнетаежный), в то время как экстремальные условия (высокогорья, степи) отсеивают видовой состав, оставляя лишь узкоспециализированные, уникальные виды.
Хорологический анализ по широтной составляющей ареала
Анализ по широтной составляющей ареала показал, что основной состав (65,2%) фауны представлен суббореальными видами (табл. 3, рис. 2).
Суббореальная группа (311 видов) составляет ядро фауны совок региона. Отличается широкой экологической амплитудой видов, многие из которых встречаются в трех-четырех вертикальных поясах. Здесь находится наибольшее число видов родов Xestia sp. (25 видов), Apamea sp. (15 видов), Euxoa sp. (14 видов). Кроме того, для группы характерны роды Mythimna sp., Orthosia sp., Lacanobia sp., Cucullia sp. На фоне других доминируют лесные виды (74% от общего состава) и отмеченные в агроценозах (45%). По трофическим связям здесь преобладают полифаги и олигофаги.
Таблица 3
Ареалогические группы совок по широтно-высотной составляющей
|
Широтно-высотная зона |
Количество видов |
Доля, % |
|
СБ |
311 |
65,2 |
|
ТМ |
72 |
15,1 |
|
Б |
43 |
9,0 |
|
АБ |
39 |
8,2 |
|
АА |
12 |
2,5 |
|
Всего |
477 |
100,0 |
Температная группа (72 вида). Здесь преобладают виды родов Agrotis sp., Autographa sp., Catocala sp. Другими отличительными особенностями являются высокая доля мигрирующих видов (25%); обилие ксерофильных видов; узкая поясность (65% видов встречаются только в одном-двух поясах); удлиненный период лета.
Бореальная группа (43 вида). Характерными представителями группы являются виды родов Xylena sp., Eupsilia sp., Brachylomia sp. В целом эта группа отличается высокой долей монотипических родов (18 %); стенотопностью видов (70% приурочены к одному-двум биотопам); адаптацией видов к условиям сомкнутых лесов; коротким периодом активности имаго; узкой трофической специализацией видов, преобладанием среди них моно- и олигофагов.
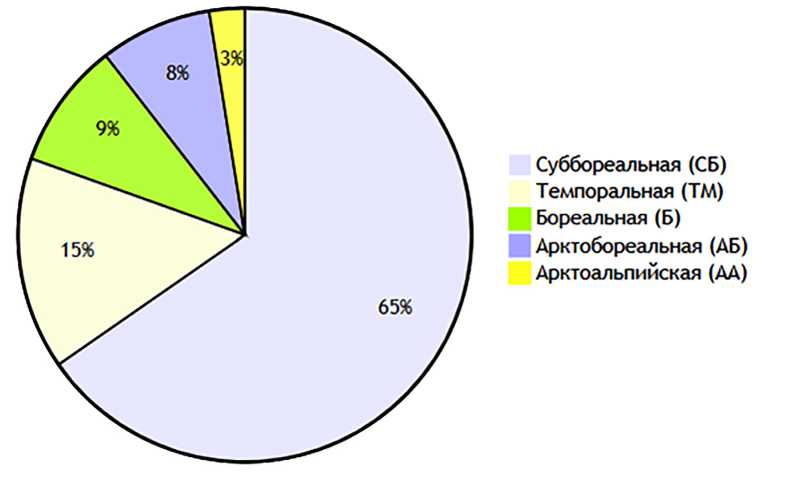
Рис. 2. Распределение видов совок по широтно-высотным зонам
Арктобореальная группа (39 видов). Характеризуется преобладанием родов Xestia sp., Polia sp., Anarta sp.; высоким уровнем оригинальности фауны (22 %); коротким жизненным циклом имаго; пищевой специализацией на стадии гусеницы на низких карликовых растениях.
Арктоальпийская группа (12 видов), высокогорно-арктический реликтовый комплекс. Уникальные виды родов Sympistis sp., Anarta sp; высокий процент эндемиков (42 %); строгая приуроченность к специфическим биотопам; островной характер распространения; монофагия (стадия гусеницы) на высокогорных растениях.
Таким образом, экологическое ядро фауны формирует суббореальная группа (311 видов) экологически пластичных видов, встречающихся в трех-четырех высотных поясах, с преобладанием лесных и связанных с агроценозами видов (74 и 45 % соответственно), доминированием полифагов и олигофагов — свидетельство высокой адаптивной способности.
Температная и бореальная группы (72 и 43 вида) характеризуются стенотоп-ностью видов (один-два высотных пояса). При этом температная группа отличается высокой долей мигрирующих видов, преобладанием ксерофилов и гусениц-полифагов, удлиненным периодом активности имаго; бореальная — узкой адаптивностью и обилием мезофильных видов, коротким периодом активности имаго, преобладанием моно- и олигофагов.
Арктобореальная и арктоальпийская группы (38 и 12 видов) отличаются высоким процентом эндемиков (38 и 42 %), коротким жизненным циклом имаго, высокой степенью биотопической приуроченности видов. Зональная дифференциация видов выражена четко — от пластичных суббореальных до стенотопных арктоальпийских. Стенотопные и оригинальные виды сконцентрированы в арктобореальной и арктоальпийской группах.
Хорологический анализ по долготной составляющей ареала
Анализ фаун Прибайкалья и Западного Забайкалья по долготной составляющей ареала показал, что их ядром являются виды с широким распространением: транспалеарктические — 37 % общего видового числа и голарктические — 11 % (табл. 4, рис. 3). Характерной чертой транспалеарктов является их эвритопность (занимают все типы биотопов — от степей до горных тундр). Виды этой группы формируют основу фауны, определяют ее общепалеарктический характер. Особенностью голарктических видов, подчеркивающих общность фауны Евразии и Северной Америки, является их связь с широко распространенными растительными ассоциациями.
Ареалогические группы совок по долготной составляющей
--ШИРОКОРАСПРОСТРАНЁННЫЕ (230 видов, 48,2%)
t Транспалеарктические (ТП) ------------------------ Голарктические (ГА) -----------------------------------
--ЗАПАДНЫЕ ПАЛЕАРКТИЧЕСКИЕ (133 вида, 27,9%)
Ь Евросибирские (ЕС) ----------------------------
Центральнопалеарктические (ЦП) ---------------------
--ВОСТОЧНЫЕ ПАЛЕАРКТИЧЕСКИЕ (88 видов, 18,4%)
--- Восточнопалеарктические (ВП) --------------------
-- Центрально-восточнопалеарктические (ЦВП) -----------
-- Южносибирско-палеархеарктические (ЮП)
__ СПЕЦИАЛИЗИРОВАННЫЕ (26 видов, 5,4%)
Ь Амфипалеарктические (АП) Космополитические (К) _____________________________
176 видов (36,9%)
51 вид (10,7%) '
56 видов (11,7%)
77 видов (16,1%)
57 видов (12,0%)
27 видов (5,7%)
4 вида (0,8%)
2 вида (0,4%)
2 вида (0,4%)
Таблица 4
|
Долготная группа |
Количество видов |
Процент от общего числа |
|
ТП |
176 |
36,9 |
|
ЦП |
77 |
16,1 |
|
ВП |
57 |
12,0 |
|
ЕС |
56 |
11,7 |
|
ГА |
51 |
10,7 |
|
ЦВП |
27 |
5,7 |
|
ЮП |
4 |
0,8 |
|
АП |
2 |
0,4 |
|
К |
2 |
0,4 |
|
Всего |
477 |
100 |
Рис. 3. Структурная схема долготных групп совок
К Западному компоненту относятся 28 % общего числа видов фауны. Сюда входят евросибирские виды (12 %), связывающие регион с европейской и западносибирской фауной. В этой группе видов, встречающихся чаще в лесных и лесостепных биотопах, преобладают центральнопалеарктические (16 %). Многие виды этой группы связаны с аридными и горными ландшафтами и отражают центральноазиатское влияние на фауну региона.
К Восточному компоненту (18%) отнесена группа видов с восточнопалеарктическим типом ареалов (12 %). Виды этой группы часто связаны с восточноазиатскими растительными группировками, такими как ильмовники ( Calyptra lata, Paragona cognata, P. multisignata, Catocala bella, C. deuteronympha и др.); влажные леса (аналоги маньчжурских широколиственных лесов) — Chrysorithrum flavomaculata, Parhylophila buddhae, Acronicta bellula и др., в том числе горнотаежные восточноазиатские формации ( Feralia sauberi, Brachyxanthia zelotypa ); восточноазиатские степи ( Cucullia fuchsiana, C. propinqua, Sympistis campicola и др.). К этому же компоненту отнесена группа видов с центрально-восточнопалеарктическим (ЦВП) и южносибирско-палеархеарктическим (ЮП) типами ареалов. Центрально-восточнопалеарктические виды представлены большей частью горностепными, а южносибирско-палеархеарктические виды ( Xylena czernilae, Xestia banghaasi, Eugraphe versuta ) являются редкими и локальными.
К особой, специализированной группе отнесены виды с амфипалеарктиче-скими ( Nola confusalis, N. karelica ) и космополитичными (К) ( Agrotis ipsilon, Xestia c-nigrum ) ареалами. Амфипалеарктические виды отмечены в регионе недавно, возможно, они появились здесь в последние десятилетия в связи с климатическими изменениями; виды-космополиты отмечены в европейской и дальневосточной части России как активные вредители, здесь они редкие и локальные.
Таким образом, региональная фауна имеет смешанный характер с преобладанием широкоареальных видов (около половины всего состава). Регион является переходной зоной между Западной и Восточной Палеарктикой. Западное влияние сильнее в полтора раза, но восточный компонент здесь хорошо выражен. Западные виды связаны с лесными и горными ландшафтами, восточные — приурочены к специфическим восточноазиатским растительным сообществам. Большинство видов представлено аборигенными элементами с устойчивыми ареалами. Редкие и специализированные группы требуют особого внимания, так как амфипалеаркти-ческие виды — возможные индикаторы климатических изменений. Космополиты демонстрируют экологическую пластичность в различных частях ареала. Высокое β-разнообразие — наличие видов с разной экологической приуроченностью — определяет устойчивость экосистем.
Биотопическая приуроченность
Экологическое ядро фауны совок региона составляют лесные и лугово-степные биотопы, главным образом лиственные/смешанные леса — 40 %, луга и степи — по 37% (табл. 5).
Лесной комплекс четко дифференцирован на светлохвойные (25 %) и влажные (20%) леса. Биотоп «все типы лесов» (10%) объединяет виды с высокой экологической пластичностью, способные существовать в различных лесных сообществах.
Аграрные ландшафты занимают седьмое место по видовому богатству (17 %), что свидетельствует об адаптивной способности многих видов совок к антропогенным преобразованиям среды.
К специализированным фаунистическим комплексам отнесен влажно-прибрежный комплекс, включающий биотопы, связанные с повышенным увлажнением. Это болота (5%), представляющие специализированные местообитания с уникальными фаунистическими комплексами. Уникальностью фауны отличаются также ильмовники (11%). Высокогорный комплекс (вл, тд, гкр) характеризуется относительно бедным, но высоким специфичным видовым составом. При этом высокогорные луга (8 %) являются переходным звеном между горными и равнинными экосистемами.
Таблица 5
Классификация видов совок по биотопической приуроченности
|
№ п/п |
Биотопы |
Сокращенное название биотопов |
Кол-во видов |
% от общего числа |
|
1 |
Лес лиственный и/или смешанный |
ллс |
181 |
37,9 |
|
2 |
Луг |
лг |
177 |
37,1 |
|
3 |
Степь |
ст |
175 |
36,7 |
|
4 |
Лес светлохвойный |
лс |
120 |
25,2 |
|
5 |
Влажные леса |
влл |
95 |
19,9 |
|
6 |
Кустарниковые заросли |
кз |
86 |
18,0 |
|
7 |
Аграрные ландшафты |
агр |
81 |
17,0 |
|
8 |
Ильмовники |
ил |
54 |
11,3 |
|
9 |
Все типы лесов |
втл |
48 |
10,1 |
|
10 |
Лес темнохвойный |
лт |
47 |
9,9 |
|
11 |
Высокогорные луга |
вл |
38 |
8,0 |
|
12 |
Болото |
бт |
24 |
5,0 |
|
13 |
Разреженные леса |
рл |
18 |
3,8 |
|
14 |
Тундра |
тд |
17 |
3,6 |
|
15 |
Горные каменистые россыпи |
гкр |
10 |
2,1 |
Четко прослеживается градиент от ксерофильных к гигрофильным условиям: степные сообщества (ст) → луговые (лг) → лесные (ллс, лс) → влажные леса (влл) → ильмовники (ил) → болота (бт). Каждая ступень этого градиента поддерживает специфический набор видов, при этом многие из них демонстрируют способность к освоению смежных по условиям увлажнения биотопов. Кустарниковые заросли (18%) имеют важную связь между лесными и открытыми местообитаниями, а разреженные леса (4%) - между сомкнутыми лесами и открытыми пространствами. Наиболее уязвимыми являются экстремальные биотопы с низким видовым богатством: горные каменистые россыпи (2 %), тундры (4 %), болота (5 %), которые требуют особого внимания при планировании охраняемых территорий. Высокое биоразнообразие в лиственных и смешанных лесах и лугах подчеркивает необходимость сохранения этих местообитаний как резервуаров фаунистического разнообразия.
Для анализа пространственной организации фауны совок на территории региона использован метод оценки сходства биотопических комплексов. В качестве меры сходства при этом применен коэффициент Сёренсена — Чекановского (рис. 4–5).
|
Биотоп |
лг |
ст |
ллс |
лс |
влл |
КЗ |
агр |
ил |
втл |
лт |
вл |
бт |
рл |
тд |
гкр |
|
лг |
1.0 |
0,7 |
0,5 |
0,4 |
0,3 |
0,6 |
0,5 |
0,2 |
0,3 |
0,2 |
0,3 |
0,2 |
0,4 |
0,1 |
0,1 |
|
ст |
0,7 |
1,0 |
0,4 |
0,3 |
0,2 |
0,5 |
0,6 |
0,2 |
0,2 |
0,1 |
0,2 |
0,1 |
0,3 |
0,1 |
0,1 |
|
ллс |
0,5 |
0,4 |
1,0 |
0,6 |
0,5 |
0,4 |
0,3 |
0,3 |
0,7 |
0,5 |
0,2 |
0,3 |
0,6 |
о,1 |
о,1 |
|
лс |
0,5 |
0,3 |
0,6 |
1,0 |
0,4 |
0,3 |
0,3 |
0.2 |
0,6 |
0,7 |
0,2 |
0,2 |
0,5 |
0,1 |
0,1 |
|
влл |
0,3 |
0,2 |
0,5 |
0,4 |
1,0 |
0,3 |
0,2 |
0.4 |
0,5 |
0,6 |
0,3 |
0,4 |
0,4 |
0,2 |
0,1 |
|
КЗ |
0,6 |
0,5 |
0,4 |
0,3 |
0,3 |
1,0 |
0,5 |
0,3 |
0,3 |
0,2 |
0,3 |
0,2 |
0,4 |
0,1 |
0,2 |
|
агр |
0,5 |
0,6 |
0,3 |
0,3 |
0,2 |
0,5 |
1,0 |
0.2 |
0,2 |
0,1 |
0,1 |
0,1 |
0,3 |
о,1 |
0,1 |
|
ил |
0,2 |
0,2 |
0,3 |
0,2 |
0,4 |
0,3 |
0,2 |
1.0 |
0,2 |
0,2 |
0,2 |
0,3 |
0,2 |
0,1 |
0,1 |
|
втл |
0,3 |
0,2 |
0,7 |
0,6 |
0,5 |
0,3 |
0,2 |
0.2 |
1,0 |
0,6 |
0,2 |
0,2 |
0,6 |
0,1 |
0,1 |
|
лт |
0,2 |
0,1 |
0,5 |
0,7 |
0,6 |
0,2 |
0,1 |
0.2 |
0,6 |
1,0 |
0,2 |
0,2 |
0,4 |
0,2 |
0,1 |
|
вл |
0,3 |
0,2 |
0,2 |
0,2 |
0,3 |
0,3 |
о,1 |
0.2 |
0,2 |
0,2 |
1,0 |
0,2 |
0,2 |
0,5 |
0,4 |
|
бт |
0,2 |
0,1 |
0,3 |
0,2 |
0,4 |
0,2 |
0,1 |
0.3 |
0,2 |
0,2 |
0,2 |
1,0 |
0,2 |
0,2 |
о,1 |
|
рл |
0,4 |
0,3 |
0,6 |
0,5 |
0,4 |
0,4 |
0,3 |
0.2 |
0,6 |
0,4 |
0,2 |
0,2 |
1,0 |
0,1 |
0,1 |
|
тд |
0,1 |
0,1 |
0,1 |
0,1 |
0,2 |
0,1 |
0,1 |
0.1 |
о,1 |
0,2 |
0,5 |
0,2 |
о,1 |
1,0 |
0,3 |
|
гкр |
0,1 |
0,1 |
о,1 |
0,1 |
0,1 |
0,2 |
0,1 |
0.1 |
0,1 |
0,1 |
0,4 |
0,1 |
0,1 |
0,3 |
1,0 |
Рис. 4. Матрица сходства биотопов (коэффициент Сёренсена — Чекановского)
Лугово-степной комплекс
Лесной комплекс
»слабое.сходство.
Влажно-прибрежный комплекс
Хвойные леса
Лиственные леса
Высокогорный комплекс
Все типы лесов втл
Болота бт
Влажные леса влл гныслабое сходство
Разреженные леса рл
Светлохвойные леса сл
Темнохвойные леса
Лиственные / смешанные ллс
Ильмовники ил
Биотопы Забайкалья
Рис. 5. Схема на основе матрицы сходства биотопов (коэффициент Сёренсена — Чекановского)
умеренное сходство
|
Луг |
Степь |
Агроландшафты |
Кустарники |
|
ЛГ |
СТ |
агр |
КЗ |
|
Высокогорные луга вл |
Тундры тд |
Горные россыпи ГКО |
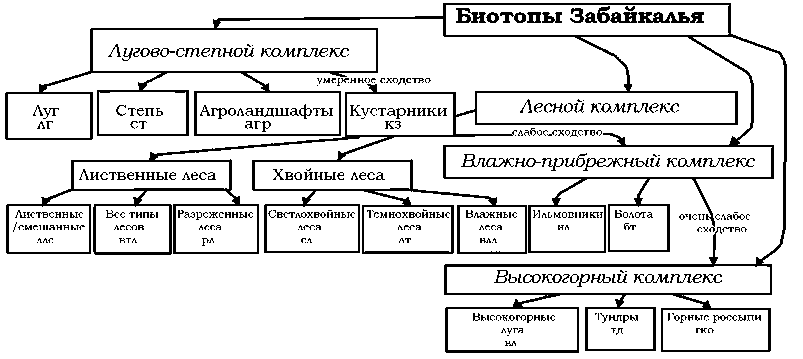
Высокое сходство лугово-степных биотопов, агроландшафтов и кустарников позволило свести их в единый лугово-степной комплекс открытых местообитаний.
Лесной комплекс четко разделяется на две подгруппы: лиственные (ллс, втл, рл) и хвойные леса (лс, лт, влл).
Влажно-прибрежный комплекс – специализированные биотопы, связанные с повышенным увлажнением. Занимают промежуточное положение между лесными и горными комплексами. Высокогорный комплекс – наиболее обособленная группа, демонстрирующая наименьшее сходство с другими биотопами.
Стрелки между кластерами показывают степень сходства — от умеренного (лугово-степной ↔ лесной) до очень слабого (влажно-прибрежный ↔ высокогорный).
Таким образом, фауна совок подразделяется на четыре четких экологических комплекса: лугово-степной, лесной, влажно-прибрежный, высокогорный. Ядро фауны формируют лесные (лиственные/смешанные — 40%) и лугово-степные (луга и степи — по 37%) биотопы.
Лесной комплекс дифференцирован на светлохвойные (25 %) и влажные (20 %) формации, при этом 10% видов демонстрируют высокую пластичность, населяя все типы лесов. Отмечается значительная синантропизация фауны – аграрные ландшафты занимают 7-е место по видовому богатству (17%).
Специализированные влажно-прибрежные (болота — 5 %), ильмовники (11 %) и высокогорные (высокогорные луга — 8%) комплексы обладают уникальной, хотя и менее богатой фауной. Четко выражен градиент увлажнения — от ксе-рофильных (степи) к гигрофильным (болота) условиям, в каждом из которых поддерживается специфический набор видов.
Экотонные биотопы (кустарники — 18%, разреженные леса — 4%) играют ключевую связующую роль. Природоохранный приоритет должны получить экстремальные биотопы с низким видовым богатством (горные россыпи — 2%, тундры — 4%, болота — 5%), а лиственные леса и луга требуют сохранения как основные резервуары биоразнообразия.
Встречаемость видов
Редкие локальные виды (41 %) в регионе — самая многочисленная группа совок (табл. 6). Суммарная доля локальных видов (О, Л + Р, Л) составляет 64 %. Это свидетельствует о том, что большинство видов совок в регионе являются стенотопными — приуроченными к специфическим местообитаниям, то есть фауна характеризуется высокой степенью экологической специализации. Суммарная доля редких видов (Р + Р, Л) составляет 44 %, то есть около половины. Это указывает на уязвимость фауны совок в регионе.
Классификация видов совок по встречаемости
Таблица 6
|
Категория |
Сокращение |
Количество видов |
Процент от общего числа |
|
Обычные нелокальные |
О |
157 |
32,9 |
|
Обычные локальные |
О, Л |
110 |
23,1 |
|
Редкие нелокальные |
Р |
14 |
2,9 |
|
Редкие локальные |
Р, Л |
196 |
41,1 |
|
Итого |
477 |
100 |
Редкие локальные виды (41 %) — наиболее уязвимая категория, требующая особых мер охраны, ориентированных на сохранение конкретных местообитаний. Они могут служить индикаторами ценных и малонарушенных экосистем. Обычные нелокальные виды (33%) широко распространены в различных биотопах, сравнительно устойчивы к антропогенному воздействию. Обычные локальные виды (23%) могут быть массовыми в специфических биотопах, указывают на наличие уникальных, но устойчивых экосистем, важных для поддержания биоразнообразия региона. Редкие нелокальные виды (3%) могут включать виды на границе ареала или виды с низкой плотностью популяций по всему региону. Такое распределение типично для многих групп насекомых в регионах с разнообразными ландшафтами и свидетельствует о высоком β-разнообразии.
Таким образом, фауна совок региона характеризуется преобладанием стенотопных и уязвимых форм. Большинство видов (64 %) являются локальными, что свидетельствует об их высокой экологической специализации и тесной связи с конкретными местообитаниями. Наиболее уязвимыми категориями являются редкие виды (44%), особенно локальные (41%), которые требуют целевых мер охраны их местообитаний.
Стабильный элемент фауны — обычные нелокальные виды (33%), относительно хорошо устойчивые к воздействиям внешних факторов. Поддерживающими уникальность экосистемы являются обычные локальные виды (23%). Подобное распределение, типичное для регионов с высоким ландшафтным разнообразием, подчеркивает важность сохранения мозаичности местообитаний для поддержания общего биоразнообразия.
Таблица 7
Фаунистический список совок Прибайкалья и Западного Забайкалья
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
1 |
Paracolax tristalis (Fabricius, 1794) |
ТМ |
ТП |
СТ, ЛС, НТ |
ил, лс, ллс, агр |
О, Л |
|
2 |
Simplicia rectalis (Eversmann, 1842) |
ТМ |
ТП |
ЛС, НТ |
ллс, агр |
О, Л |
|
3 |
Zanclognatha lunalis (Scopoli, 1763) |
СБ |
ТП |
ЛС, НТ |
ил, кз, ллс |
О |
|
4 |
Z. tarsipennalis (Treitschke, 1835) |
СБ |
ТП |
ЛС, НТ |
ллс, агр |
Р, Л |
|
5 |
Z. tenuialis Rebel, 1896 |
СБ |
ТП |
ЛС, НТ |
атл, агр |
Р, Л |
|
6 |
Z. tristriga W. Kozhantschikov, 1929 |
СБ |
ВП |
СТ, ЛС, НТ |
ллс, агр |
О |
|
7 |
Pechipogo strigilata (Linnaeus, 1758) |
ТМ |
ТП |
ЛС, НТ |
ллс, агр |
О |
|
8 |
Polypogon tentacularia (Linnaeus, 1758) |
ТМ |
ТП |
СТ, ЛС, НТ |
ллс, агр |
О |
|
9 |
Herminia grisealis ([Den.et Schiff.], 1775) |
СБ |
ТП |
ЛС, НТ |
ллс |
Р, Л |
|
10 |
Hypena crassalis (Fabricius, 1787) |
СБ |
ЦП |
НТ |
ллс |
Р, Л |
|
11 |
H. obesalis Treitschke, 1829 |
ТМ |
ЕС |
ЛС, НТ, ВТ |
ллс, кз |
О |
|
12 |
H. proboscidalis (Linnaeus, 1758) |
СБ |
ТП |
ЛС, НТ |
втл, агр |
Р, Л |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
13 |
H. rostralis (Linnaeus, 1758) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст, ллс, агр |
О |
|
14 |
H. tristalis Lederer, 1853 |
ТМ |
ТП |
СТ, ЛС, НТ |
втл, кз, агр |
Р, Л |
|
15 |
Rivula sericealis (Scopoli, 1763) |
ТМ |
ТП |
ЛС, НТ, ВТ |
лг, влл, кз |
Р, Л |
|
16 |
Scoliopteryx libatrix (Linnaeus, 1758) |
ТМ |
ГА |
ЛС, НТ |
влл, агр |
О |
|
17 |
Calyptra lata (Butler, 1881) |
СБ |
ВП |
СТ |
ил, ст |
Р, Л |
|
18 |
C. thalictri (Borkhausen, 1790) |
ТМ |
ТП |
СТ, ЛС, НТ |
влл, агр |
О |
|
19 |
Hypenodes humidalis Doubleday, 1850 |
ТМ |
ТП |
ЛС, НТ |
влл, бт |
Р, Л |
|
20 |
Parascotia fuliginaria (Linnaeus, 1761) |
СБ |
ТП |
ЛС, НТ |
влл |
Р, Л |
|
21 |
Paragona cognata (Staudinger, 1892) |
СБ |
ВП |
СТ, НТ, ВТ |
ил, влл, лт |
Р, Л |
|
22 |
P. multisignata (Christoph, 1881) |
СБ |
ВП |
СТ |
ил, ст |
Р, Л |
|
23 |
Laspeyria flexula ([Den.et Schiff.], 1775) |
ТМ |
ТП |
ЛС, НТ, ВТ |
влл, ллс, лт, лс |
Р, Л |
|
24 |
Enispa lutefascialis (Leech, 1889) |
СБ |
ВП |
ВТ |
ллс |
Р, Л |
|
25 |
Trisateles emortualis ([Den.et Schiff.], 1775) |
СБ |
ТП |
ЛС, НТ |
влл, ллс |
Р, Л |
|
26 |
Eublemma ostrina (Hübner, [1808]) |
СБ |
ЕС |
СТ |
ст, ил |
Р, Л |
|
27 |
E. rosea (Hübner, 1790) |
СБ |
ЕС |
СТ |
ст, ил |
Р, Л |
|
28 |
Phytometra viridaria (Clerck, 1759) |
ТМ |
ЕС |
СТ |
ст, ил |
Р, Л |
|
29 |
Catocala adultera Ménétriés, 1856 |
ТМ |
ТП |
СТ, ЛС, НТ |
ллс, ст, агр |
О |
|
30 |
C. bella Butler, 1877 |
СБ |
ВП |
СТ |
ст, ил |
Р, Л |
|
31 |
C. deuteronympha Staudinger, 1861 |
СБ |
ВП |
СТ |
ст, ил |
О, Л |
|
32 |
C. fraxini (Linnaeus, 1758) |
ТМ |
ТП |
СТ, ЛС, НТ |
ллс, ст, агр |
О |
|
33 |
C. fulminea (Scopoli, 1763) |
ТМ |
ТП |
СТ, ЛС, НТ |
ллс, агр |
О |
|
34 |
C. helena Eversmann, 1856 |
СБ |
ВП |
СТ |
ст, ил |
О, Л |
|
35 |
C. nupta (Linnaeus, 1767) |
ТМ |
ТП |
ЛС, НТ |
влл, ллс, кз |
О |
|
36 |
C. nymphaeoides Herrich-Schäffer, 1845 |
СБ |
ВП |
СТ |
ст, ил |
Р, Л |
|
37 |
C. pacta (Linnaeus, 1758) |
ТМ |
ТП |
ЛС, НТ, ВТ |
влл, ллс, агр |
О |
|
38 |
C. proxeneta Alphéraky, 1895 |
СБ |
ВП |
СТ |
ст, ил |
Р, Л |
|
39 |
Euclidia dentata Staudinger, 1871 |
СБ |
ЦВП |
СТ |
ст, ил, лг |
О |
|
40 |
E. glyphica (Linnaeus, 1758) |
СБ |
ТП |
СТ, ЛС, НТ |
ст, ил, лг |
О |
|
41 |
Callistege fortalitium (Tauscher, 1809) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
42 |
C. mi (Clerck, 1759) |
ТМ |
ТП |
СТ, ЛС |
лг, ллс |
О |
|
43 |
Drasteria rada (Boisduval, 1848) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
44 |
D. scolopax (Alphéraky, 1892) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
Продолжение табл. 7
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
76 |
P. ornata (Bremer, 1864) |
СБ |
ЦП |
ЛС, НТ, ВТ |
втл |
О, Л |
|
77 |
Lamprotes c-aureum (Knoch, 1781) |
СБ |
ТП |
ЛС, НТ, ВТ |
втл |
Р, Л |
|
78 |
Plusidia cheiranthi (Tauscher, 1809) |
СБ |
ТП |
ЛС, НТ, ВТ, ПГ |
лг, ллг |
Р, Л |
|
79 |
Autographa bractea ([Den.et Schiff.], 1775) |
Б |
ЕС |
ЛС, НТ |
рл |
Р, Л |
|
80 |
A. buraetica (Staudinger, 1892) |
ТМ |
ГА |
ЛС, НТ, ВТ |
ст, лг, втл, агр |
О |
|
81 |
A. excelsa (Kretschmar, 1862) |
Б |
ТП |
ЛС, НТ, ВТ |
лг, втл |
Р, Л |
|
82 |
A. gamma (Linnaeus, 1758) |
СБ |
ТП |
ЛС, НТ, ВТ |
лг, втл |
Р, Л |
|
83 |
A. macrogamma (Eversmann, 1842) |
Б |
ТП |
ЛС, НТ, ВТ |
лг, втл |
Р, Л |
|
84 |
A. mandarina (Freyer, 1845) |
Б |
ТП |
ЛС, НТ, ВТ |
лг, втл |
О, Л |
|
85 |
Syngrapha ain (Hochenwarth, 1785) |
Б |
ТП |
ЛС, НТ, ВТ |
лг, втл |
О, Л |
|
86 |
S. diasema (Boisduval, 1829) |
АБ |
ГА |
НТ, ВТ, ПГ |
вл, тд, лт |
Р, Л |
|
87 |
S. hochenwarthi (Hochenwarth, 1785) |
АБ |
ЕС |
ВТ, ПГ |
вл, тд, лт |
Р, Л |
|
88 |
S. interrogationis (Linnaeus, 1758) |
Б |
ГА |
НТ, ВТ, ПГ |
тд, бт, лт |
О |
|
89 |
S. microgamma (Hübner, 1823) |
Б |
ГА |
НТ, ВТ |
влл, лт |
Р, Л |
|
90 |
S. parilis (Hübner, 1809) |
АА |
ГА |
ПГ, ГЦ |
вл, тд |
Р, Л |
|
91 |
Plusia festucae (Linnaeus, 1758) |
Б |
ТП |
СТ, ЛС, НТ |
лг |
О |
|
92 |
P. putnami (Grote, 1873) |
СБ |
ГА |
СТ, ЛС, НТ |
лг |
О |
|
93 |
Protodeltote pygarga (Hufnagel, 1766) |
СБ |
ТП |
СТ, ЛС, НТ |
лг, рл, кз, ст |
О |
|
94 |
Deltote bankiana (Fabricius, 1775) |
СБ |
ТП |
СТ, ЛС, НТ |
лг, рл, кз, с |
О |
|
95 |
D. deceptoria (Scopoli, 1763) |
СБ |
ТП |
СТ, ЛС, НТ |
лг, рл, кз, ст |
О |
|
96 |
D. uncula (Clerck, 1759) |
СБ |
ТП |
СТ, ЛС, НТ |
лг, рл, кз, ст |
О |
|
97 |
Acontia martjanovi (Tschetverikov, 1904) |
СБ |
ЦП |
СТ, ЛС |
кз, ст |
Р, Л |
|
98 |
A. trabealis (Scopoli, 1763) |
СБ |
ТП |
СТ, ЛС |
О |
|
|
99 |
Panthea coenobita (Esper, 1785) |
Б |
ТП |
ЛС, НТ, ВТ |
лт, лс |
О, Л |
|
100 |
Thiacidas ludifica (Linnaeus, 1758) |
Б |
ТП |
ЛС, НТ, ВТ |
ллс, лт, лс |
О, Л |
|
101 |
Colocasia coryli (Linnaeus, 1758) |
Б |
ТП |
ЛС, НТ, ВТ |
ллс, лт, лс, кз |
О, Л |
|
102 |
Acronicta alni (Linnaeus, 1767) |
СБ |
ТП |
ЛС, НТ, ВТ |
ллс, лт, лс |
О |
|
103 |
A. auricoma ([Den.et Schiff.], 1775) |
Б |
ТП |
ЛС, НТ, ВТ |
ллс, влл, кз |
Р, Л |
|
104 |
A. bellula Alphéraky, 1895 |
СБ (Н) |
ВП |
ЛС, НТ, ВТ |
ллс, ил, ст |
Р, Л |
|
105 |
A. cinerea |
СБ |
ЕС |
СТ, ЛС |
ст |
Р, Л |
|
106 |
A. concerpta Draudt, 1937 |
Б |
ВП |
ЛС, НТ, ВТ |
ллс |
О |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
107 |
A. cuspis (Hübner, 1813) |
Б |
ТП |
ЛС, НТ, ВТ |
втл |
О, Л |
|
108 |
A. lutea Bremer et Grey, 1852 |
СБ (Н) |
ВП |
СТ, ЛС |
лг, рл, кз |
Р, Л |
|
109 |
A. menyanthidis (Esper, 1789) |
Б |
ТП |
ЛС, НТ |
ллс, влл, кз |
Р, Л |
|
110 |
A. psi (Linnaeus, 1758) |
Б |
ТП |
СТ, ЛС |
ст, влл, лс |
О |
|
111 |
A. rumicis (Linnaeus, 1758) |
Б |
ТП |
СТ, ЛС, НТ |
ст, влл, лс, кз, агр |
О |
|
112 |
A. strigosa ([Den.et Schiff.], 1775) |
СБ |
ТП |
СТ, ЛС |
ст, лс |
О, Л |
|
113 |
A. tridens ([Den.et Schiff.], 1775) |
Б |
ТП |
СТ, ЛС |
ст, влл, лс, кз |
О |
|
114 |
A. vulpina (Grote, 1883) |
Б |
ГА |
ЛС, НТ |
влл, лт, лс |
О |
|
115 |
Simyra albovenosa (Goeze, 1781) |
СБ |
ЕС |
СТ, ЛТ |
лг, влл |
Р, Л |
|
116 |
S. nervosa ([Den.et Schiff.], 1775) |
СБ |
ЕС |
СТ |
ст |
Р, Л |
|
117 |
S. splendida Staudinger, 1888 |
СБ |
ВП |
СТ, ЛС |
ст, влл, лс, кз |
Р, Л |
|
118 |
Mycteroplus cornuta (Püngeler, 1914) |
СБ |
ЦП |
СТ |
ст, лс |
О, Л |
|
119 |
Cucullia absinthii (Linnaeus, 1761) |
СБ |
ТП |
СТ |
ст, лс |
Р, Л |
|
120 |
C. amota Alphéraky, 1887 |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
121 |
C. argentea (Hufnagel, 1766) |
СБ |
ТП |
СТ |
ст |
О |
|
122 |
C. artemisiae (Hufnagel, 1766) |
СБ |
ТП |
СТ, ЛС |
лг, ст, лс |
О |
|
123 |
C. asteris ([Den.et Schiff.], 1775) |
СБ |
ТП |
СТ, ЛС |
лг, ст, лс |
Р, Л |
|
124 |
C. biornata Fischervon Waldheim, 1840 |
СБ |
ЕС |
СТ |
ст |
О |
|
125 |
C. biradiata W. Kozhantschikov, 1925 |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
126 |
C. distinguenda Staudinger, 1892 |
СБ |
ЦП |
СТ |
ст |
О |
|
127 |
C. elongata Butler, 1880 |
СБ |
ЦВП |
СТ |
ст |
Р, Л |
|
128 |
C. fraudatrix Eversmann, 1837 |
СБ |
ТП |
СТ, ЛС |
лг, ст |
О |
|
129 |
C. fuchsiana Eversmann, 1842 |
СБ |
ВП |
СТ, ЛС |
лг, ст, лс |
Р, Л |
|
130 |
C. humilis Boursin, 1941 |
СБ |
ЦВП |
СТ |
ст |
Р, Л |
|
131 |
C. kurilullia Bryk, 1942 |
СБ |
ЦВП |
СТ |
лг, ст, лс |
Р, Л |
|
132 |
C. lactucae ([Den.et Schiff.], 1775) |
СБ |
ЕС |
СТ |
лг, ст, лс |
Р, Л |
|
133 |
C. lindei lindei Heyne, 1899 |
СБ |
ЕС |
СТ, ЛС |
лг, ст, лс |
Р, Л |
|
134 |
C. lucifuga ([Den.et Schiff.], 1775) |
СБ |
ТП |
СТ, ЛС, НТ |
лг, ст, ллс |
О, Л |
|
135 |
C. magnifica Freyer, 1839 |
СБ |
ЕС |
СТ |
ст |
Р, Л |
|
136 |
C. papoka G. Ronkay et L. Ronkay, 1986 |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
137 |
C. praecana Eversmann, 1843 |
СБ |
ЕС |
СТ, ЛС, НТ |
лг, ст, лс, ллс |
Р, Л |
|
138 |
C. propinqua Eversmann, 1842 |
СБ |
ВП |
СТ, ЛС, НТ |
лг, ст, лс, ллс |
Р, Л |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
139 |
C. scopariae Dorfmeister, 1853 |
СБ |
ЕС |
СТ |
ст |
О, Л |
|
140 |
C. splendida (Cramer, 1777) |
СБ |
ТП |
СТ |
ст |
О |
|
141 |
C. tristis Boursin, 1934 |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
142 |
C. umbratica (Linnaeus, 1758) |
СБ |
ГА |
СТ, ЛС, НТ |
влл, лт, лс |
О |
|
143 |
C. virgaureae Boisduval, 1840 |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
144 |
Calophasia lunula (Hufnagel, 1766) |
ТМ |
ГА |
СТ, ЛС, НТ |
ст, лс, агр |
О |
|
145 |
Sympistis campicola Lederer, 1853 |
СБ |
ВП |
СТ |
ст |
О |
|
146 |
S. heliophila (Paykull, 1793) |
АБ |
ГА |
НТ, ВТ, ПГ |
тд, лс |
Р, Л |
|
147 |
S. kaszabi L. Ronkay, 1988 |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
148 |
S. senica senica (Eversmann, 1856) |
ТМ |
ЦП |
ЛС, НТ, ВТ |
ст, лс |
О, Л |
|
149 |
S. zetterstedti (Staudinger, 1857) |
АА |
ГА |
ПГ, ГЦ |
тд, вл, гкр |
Р, Л |
|
150 |
Stilbina nictitans (Bremer et Grey, 1853) |
СБ |
ВП |
СТ |
ст, лс |
Р, Л |
|
151 |
Phidrimana amurensis (Staudinger, 1892) |
СБ |
ТП |
СТ, ЛС |
ст, ил |
Р, Л |
|
152 |
Amphipyra livida ([Den.et Schiff.], 1775) |
СБ |
ТП |
ЛС, НТ |
ллс |
Р, Л |
|
153 |
A. perflua (Fabricius, 1787) |
СБ |
ТП |
ЛС, НТ, ВТ |
ллс, лс, лт |
О |
|
154 |
A. sergei (Staudinger, 1888) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
155 |
A. tetra (Fabricius, 1787) |
СБ |
ЕС |
СТ, ЛС |
ст, лс, кз |
Р, Л |
|
156 |
A. tragopoginis (Clerck, 1759) |
СБ |
ЕС |
СТ, ЛС |
втл, агр |
Р, Л |
|
157 |
Brachionycha nubeculosa (Esper, 1785) |
ТМ |
ТП |
ЛС, НТ, ВТ |
втл, ллс |
О |
|
158 |
B. sajana Draudt, 1934 |
СБ |
ЦВП |
СТ, ЛС, НТ |
ллс |
Р, Л |
|
159 |
Meganephria retinea Gyulai et L. Ronkay, 1999 |
СБ |
ЦП |
СТ |
ст, ил |
Р, Л |
|
160 |
Feralia sauberi (Graeser, 1892) |
БМ |
ВП |
НТ, ВТ |
лт, влл |
О, Л |
|
161 |
Periphanes cora (Eversmann, 1837) |
СБ |
ТП |
СТ |
ст |
Р, Л |
|
162 |
P. scutata (Staudinger, 1895) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
163 |
Pyrrhia exprimens (Walker, 1857) |
СБ |
ГА |
СТ, ЛС, НТ |
лт, влл |
Р, Л |
|
164 |
P. umbra (Hufnagel, 1766) |
СБ |
ТП |
СТ |
ст, лг |
О |
|
165 |
Schinia bieneri (Rebel, 1926) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
166 |
Protoschinia scutosa ([Den.et Schiff.], 1775) |
СБ |
ТП |
СТ |
ст |
О |
|
167 |
Heliothis adaucta Butler, 1878 |
СБ |
ТП |
СТ |
ст, лг |
О |
|
168 |
H. ononis ([Den.et Schiff.], 1775) |
СБ |
ГА |
СТ, ЛС |
лг, ст |
О |
|
169 |
Helicoverpa armigera (Hübner, 1808) |
СБ |
ТП |
СТ |
ст, лг |
О |
|
170 |
Callopistria albolineola (Graeser, 1889) |
СБ (Н) |
ВП |
ЛС, НТ |
ллс, ил, ст |
Р, Л |
|
171 |
Cryphia fraudatricula (Hübner, 1803) |
СБ |
ТП |
СТ |
ллс, ил, ст |
О, Л |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
172 |
Bryophila granitalis (Butler, 1881) |
СБ (Н) |
ВП |
ЛС, НТ |
ллс, ил, ст |
Р, Л |
|
173 |
B. orthogramma Boursin, 1954 |
СБ |
ТП |
СТ, ЛС, НТ, ВТ |
влл, лт, ллс, ил, ст |
Р, Л |
|
174 |
Victrix fabiani Varga et L. Ronkay, 1989 |
СБ |
ЦП |
СТ, ВТ, ПГ |
ллс, ил, ст, лт, тд |
Р, Л |
|
175 |
V. umovii (Eversmann, 1846) |
Б |
ЕС |
НТ, ВТ |
лс, лт |
Р, Л |
|
176 |
Athaumasta expressa (Lederer, 1855) |
СБ |
ЦП |
СТ, ЛС, НТ |
ллс, ил, ст, лс |
Р, Л |
|
177 |
A. siderigera (Christoph, 1893) |
СБ |
ЦП |
СТ, ЛС, НТ, ВТ |
ллс, ил, ст, лс, тл |
Р, Л |
|
178 |
A. splendida O. Bang-Haas, 1927 |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
179 |
Pseudeustrotia candidula ([Den.et Schiff.], 1775) |
ТМ |
ТП |
СТ, ЛС, НТ |
ллс, лс, ст |
О |
|
180 |
Caradrina albina Eversmann, 1848 |
СБ |
ЕС |
СТ, ЛС |
ст, лс |
Р, Л |
|
181 |
C. montana Bremer, 1861 |
СБ |
ГА |
СТ, ЛС, НТ, ВТ |
ллс, ил, ст, лс, тл |
О |
|
182 |
C. morosa Lederer, 1853 |
СБ |
ЦВП |
СТ, ЛС |
ст, лс |
Р, Л |
|
183 |
C. morpheus (Hufnagel, 1766) |
СБ |
ГА |
СТ, ЛС |
ст, лс |
О, Л |
|
184 |
C. petraea Tengström, 1869 |
СБ |
ЕС |
СТ, ЛС |
ст, лс |
Р, Л |
|
185 |
Hoplodrina blanda ([Den.et Schiff.], 1775) |
СБ |
ЕС |
СТ, ЛС |
ст, лс |
О |
|
186 |
H. octogenaria (Goeze, 1781) |
СБ |
ТП |
СТ, ЛС |
ст, лс, кз |
О |
|
187 |
Stygiodrina maurella (Staudinger, 1888) |
СБ |
ВП |
СТ, ЛС |
кз, ст, лс |
О, Л |
|
188 |
Chilodes distracta (Eversmann, 1848) |
СБ |
ЕС |
ЛС, НТ |
лг, влл, ллс |
Р, Л |
|
189 |
Rusina ferruginea (Esper, 1787) |
СБ |
ЕС |
СТ, ЛС, НТ |
лг, лс, кз |
Р, Л |
|
190 |
Athetis correpta (Püngeler, 1906) |
СБ |
ВП |
СТ, ЛС |
ил, ст, лг |
Р, Л |
|
191 |
A. funesta (Staudinger, 1888) |
СБ |
ВП |
СТ, ЛС |
ст, лс |
О, Л |
|
192 |
A. furvula (Hübner, 1808) |
СБ |
ТП |
СТ, ЛС |
ст, лс |
О, Л |
|
193 |
A. gluteosa (Treitschke, 1835) |
СБ |
ТП |
СТ, ЛС |
ст, лс |
Р, Л |
|
194 |
A. lepigone (Möschler, 1860) |
СБ |
ТП |
СТ, ЛС |
ст, лс |
Р, Л |
|
195 |
A. pallustris (Hübner, 1808) |
СБ |
ЕС |
СТ, ЛС, НТ |
бт, лг |
О |
|
196 |
Euplexia lucipara (Linnaeus, 1758) |
ТМ |
ТП |
ЛС, НТ, ВТ |
лс, лт, ллс |
Р, Л |
|
197 |
Auchmis mongolica (Staudinger, 1896) |
СБ |
ЦП |
СТ, ЛС, НТ, ВТ |
ст, гкр, лс, лт |
Р, Л |
|
198 |
Oxytripia orbiculosa (Esper, 1799) |
СБ |
ТП |
СТ |
ст |
Р, Л |
|
199 |
Sidemia bremeri (Erschoff, 1870) |
СБ |
ВП |
СТ, ЛС |
ст, лс |
Р, Л |
|
200 |
S. spilogramma (Rambur, 1871) |
СБ |
ВП |
СТ, ЛС |
ст, лс |
О, Л |
|
201 |
Hampsonicola deccerti (Hampson, 1908) |
СБ |
ЦП |
СТ, ЛС, НТ |
ст, лс |
О |
|
202 |
Calamia decipiens (Alphéraky, 1895) |
СБ |
ЦП |
СТ, ЛС |
ст, лс |
Р, Л |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
203 |
C. tridens (Hufnagel, 1766) |
СБ |
ЕС |
СТ |
ст |
Р, Л |
|
204 |
Staurophora celsia (Linnaeus, 1758) |
СБ |
ТП |
СТ |
ст, лс |
О |
|
205 |
Celaena haworthii (Curtis, 1829) |
Б |
ТП |
НТ, ВТ |
бт, влл |
О, Л |
|
206 |
Helotropha leucostigma (Hübner, 1808) |
СБ |
ТП |
НТ, ВТ |
бт, ллс, влл |
О, Л |
|
207 |
Hydraecia micacea (Esper, 1789) |
Б |
ГА |
СТ, ЛС, НТ, ВТ |
бт, ллс, влл, агр |
Р, Л |
|
208 |
H. mongoliensis Urbahn, 1967 |
СБ |
ВП |
СТ, ЛС |
бт, ллс, влл, ст, лг |
О |
|
209 |
H. petasitis Doubleday, 1847 |
СБ |
ТП |
СТ, ЛС |
бт, ст, лг |
Р, Л |
|
210 |
Amphipoea asiatica (Burrows, 1911) |
СБ |
ЦВП |
СТ, ЛС |
ст, влл, лс |
Р, Л |
|
211 |
A. fucosa (Freyer, 1830) |
СБ |
ТП |
СТ, ЛС, НТ |
бт, влл, ст, лг |
О |
|
212 |
A. lucens (Freyer, 1845) |
СБ |
ТП |
СТ, ЛС, НТ |
бт, ст, лг |
Р, Л |
|
213 |
Rhizedra lutosa (Hübner, 1803) |
СБ |
ГА |
СТ, ЛС, НТ |
ллс, ст, лг, агр |
О |
|
214 |
Nonagria typhae (Thunberg, 1784) |
СБ |
ТП |
СТ |
бт, ст, лг |
Р, Л |
|
215 |
Longalatedes elymi (Treitschke, 1825) |
СБ |
ТП |
СТ |
ст, лг |
О, Л |
|
216 |
Hypocoena stigmatica (Eversmann, 1855) |
ТМ |
ГА |
СТ, ЛС |
ст, лг |
О, Л |
|
217 |
Photedes fluxa (Hübner, 1809) |
СБ |
ТП |
СТ, ЛС, НТ, ВТ, ПГ |
ст, лг, бт, влл |
О, Л |
|
218 |
Globia sparganii (Esper, 1790) |
СБ |
ТП |
СТ |
ст, лг, бт |
Р, Л |
|
219 |
Eremobina pabulatricula (Brahm, 1791) |
СБ |
ТП |
СТ, ЛС, НТ |
лг, влл, лс, лт |
О, Л |
|
220 |
Apamea altijuga (W. Kozhantschikov, 1925) |
СБ |
ЦП |
ВТ, ПГ |
вл, бт, тд, лс, лт |
О, Л |
|
221 |
A. anceps ([Den.et Schiff.], 1775) |
СБ |
ТП |
СТ |
ст |
Р, Л |
|
222 |
A. crenata (Hufnagel, 1766) |
СБ |
ТП |
СТ, ЛС |
ст, лс |
О, Л |
|
223 |
A. exstincta (Staudinger, 1889) |
СБ |
ЦП |
ВТ, ПГ |
лс, лт, ст |
Р, Л |
|
224 |
A. furva ([Den.et Schiff.], 1775) |
СБ |
ЕС |
СТ |
ст |
Р, Л |
|
225 |
A. ingloria (A. Bang-Haas, 1912) |
АБ |
ГА |
ПГ, ГЦ |
вл, гкр, тд |
О, Л |
|
226 |
A. kaszabi Varga, 1982 |
СБ |
ЦП |
СТ, ПГ |
ст |
Р, Л |
|
227 |
A. lateritia (Hufnagel, 1766) |
СБ |
ТП |
СТ, ЛС, НТ |
лг, лс, ллс |
О |
|
228 |
A. leucodon (Eversmann, 1837) |
СБ |
ВП |
СТ, ЛС, ПГ |
ст |
О |
|
229 |
A. monoglypha (Hufnagel, 1766) |
СБ |
ТП |
СТ, ЛС, НТ |
лт, лс |
О, Л |
|
230 |
A. oblonga (Haworth, 1809) |
СБ |
ТП |
СТ, ЛС, ПГ |
ст, лс |
Р, Л |
|
231 |
A. remissa (Hübner, 1809) |
СБ |
ГА |
СТ |
ст |
Р, Л |
|
232 |
A. rubrirena (Treitschke, 1825) |
СБ |
ГА |
НТ, ВТ |
лс, лт, ллс |
Л, Р |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
233 |
A. sordens (Hufnagel, 1766) |
СБ |
ТП |
СТ, ЛС, ПГ |
ст, лс |
О |
|
234 |
Loscopia scolopacina (Esper, 1788) |
СБ |
ТП |
ЛС, НТ |
лг, лс, ллс |
Р, Л |
|
235 |
Oligia latruncula ([Den.et Schiff.], 1775) |
СБ |
ЕС |
СТ, ЛС |
лг |
Р, Л |
|
236 |
Ogilia leuconephra Hampson, 1908 |
СБ |
ЦВП |
СТ, ЛС |
ст, лс |
Р, Л |
|
237 |
Mesoligia furuncula ([Den. et Schiff.], 1775) |
СБ |
ТП |
СТ, ЛС |
лг, лс, ст |
Р, Л |
|
238 |
Litoligia literosa (Haworth, 1809) |
СБ |
ЕС |
СТ, ЛС |
лг, ст |
Р, Л |
|
239 |
Mesapamea secalis (Linnaeus, 1758) |
СБ |
ЕС |
СТ, ЛС |
лг, лс, ллс, кз |
Р, Л |
|
240 |
Resapamea vulpecula (Eversmann, 1852) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст, лг, лс, ллс, кз |
О |
|
241 |
Xylomoia graminea (Graeser, 1889) |
СБ |
ВП |
СТ, ЛС |
ст, лг |
О |
|
242 |
Brachylomia viminalis (Fabricius, 1777) |
Б |
ТП |
СТ, ЛС, НТ |
ст, лг, лс, лт, ллс |
О, Л |
|
243 |
Hyppa rectilinea (Esper, 1788) |
Б |
ТП |
НТ, ВТ |
лт |
О, Л |
|
244 |
Parastichtis suspecta (Hübner, 1817) |
ТМ |
ГА |
СТ, ЛС, НТ |
ст, лг, лс, ллс |
О |
|
245 |
Parvispinia parvispina (Tschetverikov, 1904) |
СБ |
ЦП |
СТ, ЛС |
ст |
О, Л |
|
246 |
Xanthia togata (Esper, 1788) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ |
ст, лг, лс, лт, ллс, агр |
О |
|
247 |
Cirrhia icteritia (Hufnagel, 1766) |
ТМ |
ТП |
СТ, ЛС, НТ |
ст, лг, лс, ллс, агр |
О |
|
248 |
C. ocellaris (Borkhausen, 1792) |
ТМ |
ЕС |
СТ, ЛС |
ст, лс |
Р |
|
249 |
C. tunicata (Graeser, 1890) |
СБ |
ЦВП |
СТ, ЛС |
ст, лс |
О |
|
250 |
Vulpechola vulpecula (Lederer, 1853) |
СБ (Н) |
ЦВП |
СТ, ЛС, НТ |
ст, лс, ллс |
О |
|
251 |
Conistra vaccinii (Linnaeus, 1761) |
СБ |
ТП |
НТ, ВТ |
лт, влл |
О |
|
252 |
Hillia iris (Zetterstedt, 1839) |
Б |
ГА |
НТ, ВТ |
лт, влл |
О, Л |
|
254 |
L. consocia (Borkhausen, 1792) |
СБ |
ТП |
НТ, ВТ |
лт, влл |
Р, Л |
|
253 |
Lithophane lamda (Fabricius, 1787) |
СБ |
ТП |
НТ, ВТ |
лт, влл |
О, Л |
|
255 |
L. socia (Hufnagel, 1766) |
СБ |
ТП |
СТ, ЛС, НТ, ВТ |
влл |
О |
|
256 |
Xylena czernilae Volynkin, 2012 |
СБ |
ЮП |
СТ, ЛС |
влл |
Р, Л |
|
257 |
X. exsoleta (Linnaeus, 1758) |
СБ |
ТП |
СТ, ЛС |
ст, лг, лс |
Р, Л |
|
258 |
X. solidaginis (Hübner, 1803) |
СБ |
ТП |
СТ, ЛС |
ст, лс |
О |
|
259 |
X. vetusta (Hübner, 1813) |
СБ |
ТП |
СТ, ЛС |
ст, лс |
О |
|
260 |
Orbona fragariae (Vieweg, 1790) |
СБ |
ТП |
СТ, ЛС |
ст, влл, лс |
Р, Л |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
261 |
Eupsilia transversa (Hufnagel, 1766) |
СБ |
ТП |
НТ, ВТ |
влл, лт, лс |
О |
|
262 |
Enargia paleacea (Esper, 1788) |
СБ |
ТП |
СТ, ЛС, НТ, ВТ |
лг, лс, лт, ллс, агр |
О |
|
263 |
Ipimorpha contusa (Freyer, 1849) |
СБ |
ТП |
НТ, ВТ |
лт, влл, ллс, агр |
Р, Л |
|
264 |
I. retusa (Linnaeus, 1761) |
СБ |
ТП |
СТ, ЛС, НТ |
лг, лс, ллс |
Р, Л |
|
265 |
I. subtusa ([Den.et Schiff.], 1775) |
СБ |
ТП |
НТ, ВТ |
ст, лс, влл, ллс, агр |
О |
|
266 |
Brachyxanthia zelotypa (Lederer, 1853) |
СБ |
ВП |
СТ, ЛС |
влл, лг |
Р, Л |
|
267 |
Cosmia pyralina ([Den.et Schiff.], 1775) |
СБ |
ТП |
СТ, ЛС |
ст, ил, ллс, агр |
О, Л |
|
268 |
C. trapezina (Linnaeus, 1758) |
ТМ |
ТП |
СТ, ЛС |
ст, ил, ллс, агр |
О, Л |
|
269 |
C. trapezinula (Filipjev, 1927) |
СБ (Н) |
ВП |
СТ, ЛС |
ст, ил, агр |
О, Л |
|
270 |
Eremohadena pugnax (Alphéraky, 1892) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
271 |
Antitype chi (Linnaeus, 1758) |
СБ |
ТП |
СТ, ЛС, НТ |
ст, ллс, агр |
Р, Л |
|
272 |
Cteipolia amissa Gordeev, Gordeeva, Ronkay et Ronkay, 2022 |
СБ |
ЦП |
ЛС, НТ |
ллс |
Р, Л |
|
273 |
Dasypolia fani Staudinger, 1892 |
СБ |
ВП |
ЛС, НТ |
лг, ллс |
Р, Л |
|
274 |
D. lama Staudinger, 1896 |
СБ |
ЦП |
ЛС, НТ |
ст, лг |
Р, Л |
|
275 |
D. templi (Thunberg, 1792) |
СБ |
ТП |
СТ, ЛС, ПГ |
ст, лг |
Р, Л |
|
276 |
Blepharita amica (Treitschke, 1825) |
Б |
ЕС |
НТ, ВТ |
ллс, лс, лт |
О |
|
277 |
Mniotype adusta (Esper, 1790) |
СБ |
ТП |
ЛС, НТ, ВТ |
ллс, лс, лт |
Р, Л |
|
278 |
M. bathensis (Lutzau, 1901) |
Б |
ТП |
СТ, ЛС, НТ |
ллс, лс, лт |
О |
|
279 |
M. satura ([Den. et Schiff.],1775) |
Б |
ТП |
СТ, ЛС, НТ |
ст, ллс, лс, лт |
Р |
|
280 |
Panolis flammea ([Den. et Schiff.], 1775) |
ТМ |
ТП |
ЛС, НТ, ВТ |
лс, лт |
О, Л |
|
281 |
Orthosia ariuna Hreblay, 1991 |
СБ |
ВП |
СТ |
ст |
О, Л |
|
282 |
O. askoldensis (Staudinger, 1892) |
СБ |
ВП |
СТ |
ст |
Р, Л |
|
283 |
O. ella (Butler, 1878) |
СБ |
ЦВП |
СТ, ЛС, НТ |
ст, ллс, лс, лт, агр |
Р, Л |
|
284 |
O. gothica (Linnaeus, 1758) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ |
ст, ллс, лс, лт, агр |
О |
|
285 |
O. gracilis ([Den. et Schiff.], 1775) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст, ллс, лс, лт, агр |
Р, Л |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
286 |
O. incerta (Hufnagel, 1766) |
СБ |
ТП |
СТ, ЛС, НТ, ВТ |
ст, ллс, лс, лт, агр |
О |
|
287 |
Anorthoa munda ([Den. et Schiff.], 1775) |
СБ |
ТП |
НТ, ВТ |
лт |
Р, Л |
|
288 |
Perigrapha circumducta (Lederer, 1855) |
СБ |
ТП |
ЛС, НТ, ВТ |
лс, лт, ллс, ил |
О |
|
289 |
Tholera cespitis ([Den.et Schiff.], 1775) |
СБ |
ЕС |
СТ, ЛС |
ст |
Р, Л |
|
290 |
Th. decimalis (Poda, 1761) |
СБ |
ЕС |
СТ, ЛС |
ст |
О, Л |
|
291 |
Cerapteryx graminis (Linnaeus, 1758) |
ТМ |
ГА |
НТ |
лг, лс, ллс |
О, Л |
|
292 |
Anarta farnhami (Grote, 1873) |
СБ |
ГА |
СТ, ЛС, ПГ |
ст |
О, Л |
|
293 |
A. melanopa (Thunberg, 1791) |
АА |
ГА |
ПГ, ГЦ |
вл, тд |
Р, Л |
|
294 |
A. odontites (Boisduval, 1829) |
СБ |
ЕС |
СТ, ПГ |
ст |
Р, Л |
|
295 |
A. sabulorum (Alphéraky, 1882) |
СБ |
ЕС |
СТ |
ст |
Р, Л |
|
296 |
A. trifolii (Hufnagel, 1766) |
ТМ |
ГА |
СТ, ЛС, НТ, ВТ |
ст, лс |
О |
|
297 |
Coranarta carbonaria (Christoph, 1893) |
ТМ |
ВП |
НТ, ВТ, ПГ |
лт, бт, тд, вл, влл |
О |
|
298 |
Sajania devagor (W. Kozhantschikov, 1923) |
АБ |
ЦП |
ВТ, ПГ |
лт, гкр |
Р, Л |
|
299 |
Polia bombycina (Hufnagel, 1766) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ |
лс, лт, ллс |
О |
|
300 |
P. hepatica (Clerck, 1759) |
АБ |
ТП |
НТ, ВТ, ПГ |
лт, ллс, тд |
О, Л |
|
301 |
P. lama (Staudinger, 1896) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
302 |
P. lamuta (Herz, 1903) |
АБ |
ТП |
ВТ, ПГ, ГЦ |
тд |
Р, Л |
|
303 |
P. malchani (Draudt, 1934) |
АБ |
ЦВП |
НТ, ВТ |
лт, лс |
О, Л |
|
304 |
P. nebulosa (Hufnagel, 1766) |
ТМ |
ТП |
ЛС, НТ, ВТ |
лс, лг, кз, лт, ллс |
О |
|
305 |
P. richardsoni (Curtis, 1834) |
АА |
ГА |
ПГ |
вл, тд |
Р, Л |
|
306 |
P. subcontigua (Eversmann, 1852) |
СБ |
ЦП |
СТ, ЛС, ПГ, ГЦ |
ст, лс, вл |
О, Л |
|
307 |
P. tiefi Püngeler, 1914 |
АБ |
ЦВП |
ВТ |
лт, лс, вл |
О, Л |
|
308 |
P. vespertilio (Draudt, 1934) |
АБ |
ЦВП |
ВТ |
лт, лс |
О, Л |
|
309 |
P. vesperugo (Eversmann, 1856) |
АБ |
ТП |
ВТ |
лт, лс |
О, Л |
|
310 |
Pachetra sagittigera (Hufnagel, 1766) |
СБ |
ЕС |
СТ |
ст |
О, Л |
|
311 |
Lasianobia lauta (Püngeler, 1900) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
312 |
Lacanobia aliena (Hübner, 1808) |
СБ |
ТП |
СТ, ЛС |
ст, лг, лс |
О |
|
313 |
L. contigua ([Den.et Schiff.], 1775) |
ТМ |
ТП |
ЛС, НТ, ВТ |
лг, лс, лт, ллс |
О |
|
314 |
L. contrastata (Bryk, 1942) |
СБ |
ВП |
ЛС, НТ |
лс |
Р, Л |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
315 |
L. mongolica Behounek, 1992 |
СБ |
ЦП |
СТ |
ст |
О |
|
316 |
L. oleracea (Linnaeus, 1758) |
ТМ |
ТП |
ЛС, НТ |
лг, лс, ллс |
Р |
|
317 |
L. splendens (Hübner, 1808) |
ТМ |
ТП |
ЛС, НТ |
лг, лс, ллс |
Р |
|
318 |
L. suasa ([Den. et Schiff.],1775) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ |
лг, ст, втл |
О |
|
319 |
L. thalassina (Hufnagel, 1766) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст, лс, ллс |
О |
|
320 |
L. w-latinum (Hufnagel, 1766) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст, лс |
Р, Л |
|
321 |
Melanchra persicariae (Linnaeus, 1761) |
ТМ |
ТП |
НТ |
лг, лс, лт |
О, Л |
|
322 |
Ceramica pisi (Linnaeus, 1758) |
ТМ |
ТП |
НТ |
лг, лс, лт |
О, Л |
|
323 |
Papestra biren (Goeze, 1781) |
АБ |
ГА |
СТ, ЛС, НТ, ВТ, ПГ |
ст, лг, лс, вл, тд |
О, Л |
|
324 |
Hada plebeja (Linnaeus, 1761) |
Б |
ТП |
СТ, ЛС, НТ, ПГ |
ст, лс |
О |
|
325 |
Hyssia cavernosa (Eversmann, 1842) |
СБ |
ТП |
СТ |
ст |
О |
|
326 |
Mamestra brassicae (Linnaeus, 1758) |
ТМ |
ТП |
СТ, ЛС, НТ |
лг, лс |
Р, Л |
|
327 |
Cornutifera simplex (Staudinger, 1889) |
СБ |
ЦП |
СТ, ПГ |
ст |
Р, Л |
|
328 |
Sideridis egena (Lederer, 1853) |
СБ |
ЕС |
СТ, ПГ, ГЦ |
ст |
О, Л |
|
329 |
S. lampra (Schawerda, 1913) |
СБ |
ЕС |
ЛС |
ст |
Р |
|
330 |
S. rivularis (Fabricius, 1775) |
СБ |
ТП |
СТ, ЛС, НТ |
ст, лс |
О |
|
331 |
S. turbida (Esper, 1790) |
СБ |
ТП |
СТ, ЛС, НТ |
ст, лс |
О |
|
332 |
Heliophobus kitti (Schawerda, 1914 |
СБ |
ТП |
СТ |
ст |
О |
|
333 |
H. reticulata (Goeze, 1781) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст, лг, лс |
О |
|
334 |
Saragossa incerta (Staudinger, 1896) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
335 |
Conisania arida (Lederer, 1855) |
СБ |
ЦП |
СТ |
ст |
О |
|
336 |
C. suavis (Staudinger, 1892) |
СБ |
ВП |
СТ, ЛС |
ст, лс |
Р, Л |
|
337 |
Hecatera bicolorata (Hufnagel, 1766) |
ТМ |
ТП |
СТ |
ст |
Р, Л |
|
338 |
Hadena aberrans (Eversmann, 1856) |
СБ |
ВП |
СТ, ЛС |
ст, лс |
О, Л |
|
339 |
H. capsincola ([Den. et Schiff.],1775) |
СБ |
ЕС |
ЛС, НТ |
лг, лс |
Р, Л |
|
340 |
H. compta ([Den. et Schiff.],1775) |
ТМ |
ТП |
СТ, ЛС, НТ |
ст, лг, лс |
Р, Л |
|
341 |
H. confusa (Hufnagel, 1766) |
СБ |
ТП |
СТ, ЛС, НТ |
ст, лг, лс |
О, Л |
|
342 |
H. corrupta (Herz, 1898) |
СБ |
ВП |
СТ, ЛС, НТ |
ст, лс |
О, Л |
|
343 |
H. perplexa ([Den. et Schiff.], 1775) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст, лг, лс |
Р, Л |
|
344 |
H. variolata (Smith, 1888) |
СБ |
ГА |
СТ, ЛС, НТ, ВТ, ПГ |
ст, лг, лс |
О |
|
345 |
Mythimna albiradiosa (Eversmann, 1852) |
СБ |
ТП |
СТ, ЛС, НТ |
лг, ст, лс, ллс |
Р, Л |
|
346 |
M. atrata Remm et Viidalepp, 1979 |
СБ |
ЦП |
СТ |
ст, лг |
Р |
|
347 |
M. conigera ([Den. et Schiff.], 1775) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ, ПГ, ГЦ |
ст, лг, бт, втл |
О |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
348 |
M. deserticola (Bartel, 1903) |
СБ |
ЦП |
СТ |
ст |
Р |
|
349 |
M. impura (Hübner, 1808) |
ТМ |
ТП |
ЛС, НТ |
лг, лс |
О, Л |
|
350 |
M. opaca (Staudinger, 1900) |
СБ |
ВП |
СТ |
ст |
О |
|
351 |
M. pallens (Linnaeus, 1758) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ |
ст, лг, бт, втл |
О |
|
352 |
M. pudorina ([Den. et Schiff.], 1775) |
СБ |
ТП |
ЛС, НТ |
лг, влл |
О, Л |
|
353 |
M. rufipennis Butler, 1878 |
СБ |
ВП |
НТ |
лг, влл |
Р, Л |
|
354 |
M. turca (Linnaeus, 1761) |
СБ |
ТП |
НТ |
лг, влл |
Р, Л |
|
355 |
M. velutina (Eversmann, 1846) |
СБ |
ВП |
СТ |
ст |
О, Л |
|
356 |
Leucania comma (Linnaeus, 1761) |
СБ |
ТП |
СТ, ЛС, НТ |
ст, лг |
О |
|
357 |
Lasionycta alpicola Lafontaine et Kononenko, 1988 |
АБ |
ЦП |
ВТ, ПГ, ГЦ |
лс, тд, вл |
Р, Л |
|
358 |
L. buraetica Kononenko, 1988 |
АА |
ЦП |
ПГ, ГЦ |
вл, тд |
Р, Л |
|
359 |
L. hospita A. Bang-Haas, 1912 |
АБ |
ЦВП |
СТ, ЛС, НТ, ВТ |
кз, лс |
О |
|
360 |
L. leucocycla (Staudinger, 1857) |
АБ |
ТП |
СТ, ЛС, НТ, ВТ |
ст, кз, вл, тд лс |
О, Л |
|
361 |
L. secedens (Walker, 1858) |
АБ |
ГА |
СТ, НТ, ВТ, ПГ, ГЦ |
ст, кз, вл, тд лс |
О, Л |
|
362 |
L. skraelingia (Herrich-Schäffer, 1852) |
АБ |
ГА |
ВТ, ПГ, ГЦ |
лс, бт, вл |
Р, Л |
|
363 |
L. staudingeri (Aurivillius, 1891) |
АА |
ГА |
ПГ, ГЦ |
вл, тд |
Р, Л |
|
364 |
Eriopygodes imbecilla (Fabricius, 1794) |
ТМ |
ЕС |
ЛС, НТ |
лг,кз, лс |
Р, Л |
|
365 |
Lasionhada proxima (Hübner, 1809) |
ТМ |
ТП |
СТ, ПГ |
ст, лг |
О |
|
366 |
Actebia confinis (Staudinger, 1881) |
СБ |
ЦП |
СТ |
ст |
Р |
|
367 |
A. confusa (Alphéraky, 1882) |
СБ |
ЦП |
СТ |
ст |
Р |
|
368 |
A. difficilis (Erschoff, 1887) |
СБ |
ЦВП |
СТ |
ст |
О |
|
369 |
A. fennica (Tauscher, 1837) |
Б |
ГА |
СТ, ЛС, НТ |
ст, лг |
О |
|
370 |
A. praecox (Linnaeus, 1758) |
СБ |
ТП |
ЛС, НТ |
ллс |
О |
|
371 |
A. praecurrens (Staudinger, 1888) |
СБ |
ЦВП |
ЛС, НТ |
ллс |
О |
|
372 |
A. squalida (Guenée, 1852) |
СБ |
ТП |
СТ, ЛС, НТ, ВТ, ПГ, ГЦ |
лг, ст, втл |
О |
|
373 |
Dichagyris ignara (Staudinger, 1896) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
374 |
D. musiva (Hübner, 1803) |
СБ |
ЕС |
СТ |
ст, кз |
О |
|
375 |
D. plumbea (Alphéraky, 1887) |
СБ |
ЦП |
СТ |
ст |
О, Л |
|
376 |
D. spissilinea (Staudinger, 1897) |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
377 |
D. stentzi (Lederer, 1853) |
АБ |
ВП |
СТ, ЛС, НТ, ВТ |
ст, лс |
Р, Л |
|
378 |
Euxoa adumbrata (Eversmann, 1842) |
ТМ |
ГА |
СТ, ЛС, НТ, ВТ, ПГ, ГЦ |
ст, лг, лс |
О |
|
379 |
E. aquilina ([Den. et Schiff.], 1775) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст, лг |
Р, Л |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
380 |
E. cursoria (Hufnagel, 1766) |
СБ |
ГА |
СТ |
ст |
О |
|
381 |
E. eruta (Hübner, 1827) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст, лг |
Р |
|
382 |
E. intolerabilis (Püngeler, 1902) |
СБ |
ЦВП |
СТ, ЛС |
ст |
Р, Л |
|
383 |
E. nigricans (Linnaeus, 1761) |
ТМ |
ТП |
СТ, ЛС, НТ |
ст, лг |
О |
|
384 |
E. novoobscurior Bryk, 1948 |
СБ |
ВП |
СТ, ЛС |
ст |
Р, Л |
|
385 |
E. ochrogaster (Guenée, 1852) |
ТМ |
ГА |
СТ, ЛС, НТ, ВТ |
ст, лг, втл |
О |
|
386 |
E. phantoma I. Kozhantshikov, 1928 |
СБ |
ЦВП |
СТ |
ст |
О |
|
387 |
E. recussa (Hübner, 1817) |
СБ |
ВП |
СТ, ЛС, НТ |
ст, лг, лс |
Р, Л |
|
388 |
E. sibirica (Boisduval, 1832) |
СБ |
ВП |
СТ, ЛС, НТ, ВТ, ПГ, ГЦ |
ст, лг, втл |
Р, Л |
|
389 |
E. steideli Kononenko, 2018 |
СБ |
ЦП |
СТ, ЛС, ПГ |
ст |
Р, Л |
|
390 |
E. tristis (Staudinger, 1897) |
СБ |
ЕС |
СТ |
ст |
Р, Л |
|
391 |
E. tritici (Linnaeus, 1761) |
СБ |
ЕС |
СТ |
ст |
Р, Л |
|
392 |
Feltia honesta (Staudinger, 1892) |
АБ |
ЦП |
СТ, ПГ, ГЦ |
ст, тд |
О, Л |
|
393 |
F. nigrita (Graeser, 1892) |
АБ |
ГА |
СТ, ПГ, ГЦ |
ст, тд |
О, Л |
|
394 |
Agrotis clavis (Hufnagel, 1766) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ, ПГ, ГЦ |
ст, лг, втл, агр |
О |
|
395 |
A. desertorum Boisduval, 1840 |
СБ |
ЕС |
СТ |
ст |
О |
|
396 |
A. exclamationis (Linnaeus, 1758) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ |
ст, лг, втл, агр |
О |
|
397 |
A. fatidica (Hübner, 1824) |
АБ |
ТП |
СТ |
ст |
Р, Л |
|
398 |
A. humigena Püngeler, 1899 |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
399 |
A. ipsilon (Hufnagel, 1766) |
ТМ |
К |
СТ, ЛС, НТ, ВТ |
ст, лг, втл, агр |
Р, Л |
|
400 |
A. robusta Eversmann, 1856 ( characteristica ) |
СБ |
ВП |
СТ |
ст |
Р, Л |
|
401 |
A. ruta (Eversmann, 1851) |
АА |
ГА |
ПГ, ГЦ |
ст, вл, тд |
О, Л |
|
402 |
A. segetum ([Den.et Schiff.], 1775) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ, ПГ, ГЦ |
ст, лг, втл, агр |
О |
|
403 |
A. submolesta Püngeler, 1899 |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
404 |
A. trifurca Eversmann, 1837 |
СБ |
ТП |
СТ, ЛС, НТ |
ст, лг, ллс |
О, Л |
|
405 |
A. vestigialis (Hufnagel, 1766) |
СБ |
ЕС |
СТ |
ст |
Р, Л |
|
406 |
Ochropleura plecta (Linnaeus, 1761) |
ТМ |
ГА |
ЛС, НТ, ВТ |
лг, лс |
О |
|
407 |
Diarsia brunnea ([Den. et Schiff.], 1775) |
Б |
ТП |
НТ, ВТ |
втл |
О |
|
408 |
D. canescens (Butler, 1878) |
Б |
ВП |
ЛС, НТ, ВТ |
лг, ллс |
Р, Л |
|
409 |
D. dahlii (Hübner, 1813) |
Б |
ТП |
СТ, ЛС, НТ, ВТ |
ст, лг, втл, агр |
О |
|
410 |
D. mendica (Fabricius, 1775) |
Б |
ТП |
СТ, ЛС, НТ, ВТ |
ст, лг, втл |
О, Л |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
411 |
D. rubi (Vieweg, 1790) |
Б |
ЕС |
НТ, ВТ |
втл |
О, Л |
|
412 |
Cerastis leucographa ([Den. et Schiff.], 1775) |
СБ |
ТП |
СТ, ЛС, НТ |
кз, лс |
О, Л |
|
413 |
C. orientalis Boursin, 1948 |
СБ |
ТП |
НТ |
ллс |
Р, Л |
|
414 |
C. rubricosa ([Den.et Schiff.], 1775) |
СБ |
ТП |
СТ, ЛС, НТ |
ст, кз, лс |
О, Л |
|
415 |
Paradiarsia coturnicola (Graeser, 1892) |
СБ |
ЦП |
СТ, ПГ, ГЦ |
ст, тд, вл |
О, Л |
|
416 |
P. punicea (Hübner, 1803) |
ТМ |
ТП |
НТ |
ллс |
О, Л |
|
417 |
Netrocerocora quadrangula (Eversmann, 1844) |
СБ |
ЕС |
СТ |
ст |
Р, Л |
|
418 |
Lycophotia cissigma (Ménétriés, 1859) |
СБ |
ВП |
НТ, ВТ |
втл |
Р, Л |
|
419 |
Rhyacia caradrinoides (Staudinger, 1897) |
СБ |
ЦП |
СТ, ПГ, ГЦ |
ст, тд |
Р, Л |
|
420 |
Rh. junonia (Staudinger, 1881) |
СБ |
ЦП |
СТ, ПГ, ГЦ |
ст, вл |
Р, Л |
|
421 |
Rh. ledereri (Erschoff, 1870) |
СБ |
ЦВП |
СТ, ПГ, ГЦ |
ст, тд |
О, Л |
|
422 |
Rh. schistochroa Varga, 1973 |
СБ |
ЦП |
СТ, ПГ, ГЦ |
ст, тд |
Р, Л |
|
423 |
Rh. simulans (Hufnagel, 1766) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст |
Р, Л |
|
424 |
Chersotis andereggii (Boisduval, 1832) |
СБ |
ТП |
СТ |
ст |
Р, Л |
|
425 |
Ch. deplanata (Eversmann, 1843) |
СБ |
ВП |
СТ, ЛС, НТ |
ст, кз, лс |
Р, Л |
|
426 |
Ch. transiens (Staudinger, 1897) |
СБ |
ЦП |
СТ, ПГ, ГЦ |
ст, тд |
О, Л |
|
427 |
Noctua interposita (Hübner, 1790) |
СБ |
ЕС |
СТ, ЛС, НТ |
ст, кз |
Р, Л |
|
428 |
Cryptocala chardinyi (Boisduval, 1829) |
Б |
ТП |
СТ, ЛС, НТ |
лг |
О |
|
429 |
Spaelotis ravida ([Den.et Schiff.], 1775) |
Б |
ТП |
СТ, ЛС |
ст, лг |
Р, Л |
|
430 |
S. sennina Boursin, 1955 |
СБ |
ЦП |
СТ |
ст |
Р, Л |
|
431 |
S. suecica (Aurivillius, 1889) |
АБ |
ТП |
СТ |
ст |
О, Л |
|
432 |
Eurois occulta (Linnaeus, 1758) |
Б |
ГА |
СТ, ЛС, НТ, ВТ |
ст, лг, кз, втл, агр |
О |
|
433 |
Anaplectoides prasina ([Den.et Schiff.], 1775) |
Б |
ГА |
СТ, ЛС, НТ, ВТ |
ст, лг, кз, втл, агр |
О |
|
434 |
Xestia albonigra (Kononenko, 1981) |
СБ |
ВП |
ВТ |
лт |
Р, Л |
|
435 |
X. albuncula (Eversmann, 1851) |
АБ |
ГА |
НТ, ВТ, ПГ |
вл, лт, лс |
О |
|
436 |
X. ashworthii (Doubleday, 1855) |
СБ |
ЕС |
СТ |
ст |
Р, Л |
|
437 |
X. atrata (Morrison, 1874) |
АБ |
ГА |
НТ, ВТ |
влл, лт |
О |
|
438 |
X. baja ([Den.et Schiff.], 1775) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ |
ст, лг, кз, втл, агр |
О |
|
439 |
X. banghaasi (Corti et Draudt, 1933) |
АБ |
ЮП |
ВТ, ПГ, ГЦ |
лс, вл |
Р, Л |
|
440 |
X. borealis (Nordström, 1933) |
АБ |
ЕС |
НТ, ВТ |
лс, лт |
Р, Л |
|
441 |
X. brunneopicta (Matsumura, 1925) |
АБ |
ТП |
НТ, ВТ, ПГ |
лс, вл, бт |
О, Л |
|
442 |
X. c-nigrum (Linnaeus, 1758) |
ТМ |
К |
СТ, ЛС, НТ, ВТ |
ст, лг, кз, втл, агр |
О |
|
443 |
X. collina (Boisduval, 1840) |
АБ |
ТП |
СТ, ЛС, НТ |
ст, лг, кз,лс |
О |
Продолжение табл. 7
|
№ п/п |
Вид |
Ареал |
Пояс |
Биотоп |
Статус |
|
|
шир. |
долг. |
|||||
|
444 |
X. descripta (Bremer, 1861) |
АБ |
ЦВП |
НТ, ВТ |
лг, лс |
О, Л |
|
445 |
X. ditrapezium ([Den.et Schiff.], 1775) |
ТМ |
ТП |
СТ, ЛС, НТ, ВТ |
ст, лг, кз, агр |
О |
|
446 |
X. gelida (Sparre-Schneider, 1883) |
АБ |
ТП |
НТ, ВТ |
лт, лс |
О, Л |
|
447 |
X. herrichschaefferi (Alphéraky, 1895) |
АА |
ЦП |
СТ, ПГ, ГЦ |
ст, тд |
Р, Л |
|
448 |
X. kollari (Lederer, 1853) |
СБ |
ЦВП |
СТ |
ст |
О |
|
449 |
X. kolymae kolymae (Herz, 1903) |
Б |
ГА |
ВТ |
ст, лс |
О, Л |
|
450 |
X. laetabilis (Zetterstedt, 1839) |
АБ |
ГА |
НТ, ВТ |
лт, лс |
О, Л |
|
451 |
X. lorezi (Staudinger, 1891) |
АА |
ГА |
ВТ, ПГ, ГЦ |
лс, вл, тд |
О, Л |
|
452 |
X. penthima (Erschoff, 1870) |
Б |
ВП |
НТ, ВТ |
гкр |
О, Л |
|
453 |
X. quieta (Hübner, 1813) |
АА |
ГА |
ПГ, ГЦ |
гкр, тд |
Р, Л |
|
454 |
X. rhaetica (Staudinger, 1871) |
АБ |
ТП |
НТ, ВТ |
лт, лс |
О, Л |
|
455 |
X. rodionovi Mikkola, 1996 |
АА |
ЦП |
ПГ, ГЦ |
тд |
Р, Л |
|
456 |
X. senescens (Staudinger, 1881) |
СБ |
ЦП |
СТ, ВТ, ПГ, ГЦ |
ст, лс, тд |
О, Л |
|
457 |
X. sincera (Herrich-Schäffer, 1851) |
АБ |
ТП |
НТ, ВТ |
лт, лс |
О, Л |
|
458 |
X. speciosa (Hübner, 1813) |
АБ |
ГА |
НТ, ВТ |
лт, лс |
О |
|
459 |
X. subgrisea (Staudinger, 1897) |
АБ |
ЦВП |
НТ, ВТ |
ллс, лс |
О, Л |
|
460 |
X. tecta (Hübner, 1808) |
АА |
ГА |
ПГ, ГЦ |
ст, вл, тд |
О, Л |
|
461 |
X. triangulum (Hufnagel, 1766) |
СБ |
ТП |
СТ, НТ, ВТ |
ст, лг |
Р, Л |
|
462 |
X. ursae (McDunnough, 1940) |
Б |
ГА |
ВТ, ПГ |
ст, лс |
Р, Л |
|
463 |
X. wockei (Möschler, 1862) |
АБ |
ГА |
НТ, ВТ, ПГ, ГЦ |
вл, тд, лс |
О |
|
464 |
Protolampra sobrina (Duponchel, 1843) |
Б |
ТП |
СТ, ЛС, НТ, ВТ |
ст, лг, кз, ллс |
О, Л |
|
465 |
Sineugraphe exusta (Butler, 1878) |
ТМ |
ВП |
ЛС, НТ |
лг, ллс |
О, Л |
|
466 |
Prognorisma albifurca (Erschoff, 1877) |
СБ |
ЦП |
СТ, ПГ, ГЦ |
ст, тд |
О, Л |
|
467 |
Opigena polygona ([Den.et Schiff.], 1775) |
СБ |
ТП |
СТ |
ст, кз |
Р, Л |
|
468 |
Pseudohermonassa melancholica (Lederer, 1853) |
СБ |
ЦВП |
СТ, НТ, ВТ |
ст, лг |
О |
|
469 |
P. ononensis (Bremer, 1861) |
АБ |
ЦВП |
СТ, ПГ, ГЦ |
ст, тд |
О |
|
470 |
Coenophila subrosea (Stephens, 1829) |
Б |
ТП |
СТ, ЛС, НТ, ВТ |
лг, влл |
О, Л |
|
471 |
Eugraphe sigma ([Den.et Schiff.], 1775) |
ТМ |
ТП |
НТ, ВТ |
лг, втл |
О |
|
472 |
E. versuta (Püngeler, 1908) |
СБ |
ЮП |
СТ |
ст |
Р, Л |
|
473 |
Graphiphora augur (Fabricius, 1775) |
Б |
ТП |
СТ, ЛС, НТ, ВТ |
лг, втл |
О |
|
474 |
Naenia contaminata (Walker, 1865) |
СБ |
ВП |
СТ, ЛС |
ст, ллс |
О, Л |
|
475 |
Nyssocnemis eversmanni (Lederer, 1853) |
СБ |
ЦВП |
СТ, ЛС |
ст, кз |
О |
|
476 |
Isochlora grumi Alphéraky, 1892 |
СБ |
ЦП |
ЛС, НТ, ВТ |
ст |
Р, Л |
|
477 |
I. viridis Staudinger, 1882 |
СБ |
ЦП |
ПГ, ГЦ |
ст, тд |
Р, Л |
Выводы
На территории Прибайкалья и Западного Забайкалья к настоящему времени достоверно установлено присутствие 477 видов совок.
Максимальное видовое богатство сосредоточено в переходных зонах (лесостепь, нижняя тайга), тогда как узкоспециализированные виды преобладают в экстремальных условиях (степи, высокогорья).
В хорологическом аспекте фауна совок Прибайкалья и Забайкалья имеет смешанный характер. Регион является переходной зоной между Западной и Восточной Палеарктикой. Западное влияние сильнее в полтора раза, но и восточный компонент здесь хорошо выражен.
Региональная фауна совок подразделяется на четыре четких экологических комплекса: лугово-степной; лесной; влажно-прибрежный; высокогорный. Ядро фауны формируют лесные и лугово-степные биотопы. Ключевую связующую роль играют экотонные биотопы (кустарники, разреженные леса).
Преобладание локальных (64 %) и редких (44 %) видов указывает на высокую уязвимость фауны и необходимость сохранения мозаичности местообитаний для поддержания биоразнообразия.
