Общая минерализация и химический состав воды как важные факторы для изучения экобиоморф и экологических ниш гидромакрофитов

Автор: Свириденко Б.Ф.
Журнал: Вестник Нижневартовского государственного университета @vestnik-nvsu
Рубрика: Экология растений
Статья в выпуске: 4 (72), 2025 года.
Бесплатный доступ
Цель работы заключается в обосновании подходов экобиоморф к гидромакрофитов дифференциации на основе количественных данных о галотолерантности. Изучение экобиоморф и экологических ниш гидромакрофитов является современной важной гидроботаники. задачей Большинство существующих систем жизненных форм водных макроскопических растений не содержит какуюлибо количественную информацию об отношении гидромакрофитов к абиотическим факторам водной среды. Общая минерализация (соленость) наряду с общей жесткостью и активной реакцией воды – важные факторы, оказывающие значительное влияние на водные растения. Количественные диапазоны значений этих факторов, установленные в ходе лабораторного анализа проб воды из природных экотопов, морфологическими наряду с характеристиками биологослужат основой для дифференциации экобиоморф и экологических ниш каждого вида. На основе исследований, выполненных в период с 1984 г. по 2023 г., установлены количественные диапазоны минерализации воды для 247 видов гидромакрофитов Западно-Сибирской равнины. В составе водной макрофитной флоры этого региона выделены 2 флористических комплекса: пресноводный и соляноводный. Естественной экологической границей между данными комплексами является диапазон минерализации 8–25 г/дм3, получивший в гидробиологии название критической солености. Виды пресноводного комплекса, ограниченные в своем распространении минерализацией до 0,2 г/дм3, считаются ультрапресноводными, до 1 г/дм3 – типично пресноводными, до 3 г/дм3 – условнопресноводными, до 5 г/дм3 – слабосолоноватопресноводными, до 8 г/дм3 – среднесолоноватопресноводными, до 25 г/дм3 – сильносолоноватопресноводными. Виды, обитающие в диапазоне минерализации от 8 до 80 г/дм3, образуют соляноводный комплекс. Указанные градации использованы при выделении экобиоморф гидромакрофитов наряду с другими экологическими параметрами (концентрация главных ионов и растворимых форм некоторых металлов, общая жесткость, рН воды). Эти границы разделяют экологические ниши видов и позволяют прогнозировать вероятный состав гидрофитоценозов на основе сведений о минерализации и составе воды в конкретных водных объектах.
Поверхностные воды, главные ионы, макроскопические гидрофиты, галотолерантность, экологические ниши
Короткий адрес: https://sciup.org/14134400
IDR: 14134400 | УДК: 58.04 | DOI: 10.36906/2311-4444/25-4/03
Total Salt Content and Chemical Composition of Water as Important Factors for the Study of Ecobiomorphes and Ecological Niches of Hydromacrophytes
The aim of the work is to substantiate approaches to the differentiation of ecobiomorphes of hydromacrophytes based on quantitative data on halotolerance. The study of ecobiomorphes and ecological niches of hydromacrophytes is an important problem of hydrobotany. Most of the existing systems of the life-forms of aquatic macrophytes do not contain any quantitative information on the relation of these plants to the abiotic factors of the aquatic environment. Total salt content along with total hardness and pH of water are important factors strongly affecting the aquatic plants. Quantitative ranges of these factors revealed upon laboratory tests of water samples from natural ecotopes along with their biological-morphological parameters form the basis for the differentiation of the ecobiomorphes and the ecological niches of each species. It was demonstrated that in continental water bodies, quantitative information on salt content and composition of water can only be obtained based on laboratory tests. Based on studies conducted in 19842023, quantitative ranges for water salt content for 247 species of hydromacrophytes in the West Siberian Plain were obtained. In the aquatic marcophyte flora of this region, 2 floristic systems were identified: a freshwater one and a saltwater one. The natural ecological threshold between these systems is the salt content range of 8-25 g/dm3, which is called critical salt content in hydrobiology. Species of a freshwater system limited within a salt content of up to 0.2 g/dm3 are called ultrafreshwater, up to 1 g/dm3 typical freshwater, up to 3 g/dm3 conditionally freshwater, up to 5 g/dm3 slightly saltish freshwater, up to 8 g/dm3 middle-saltish freshwater, up to 25 g/dm3 strongly saltish freshwater. Species living in a salt content range between 8 to 80 g/dm3 form a saltwater system. These grades are used to identify ecobiomorphes of hydromacrophytes in combination with other ecological parameters (concentration of main ions and soluble forms of certain metals, total hardness and pH of water). These borders divide the ecological niches of the species making it possible to predict potential composition of hydrophyte conenoses based on the information on water salt content and composition in specific water bodies.
Текст научной статьи Общая минерализация и химический состав воды как важные факторы для изучения экобиоморф и экологических ниш гидромакрофитов
Свириденко Б.Ф. Общая минерализация и химический состав воды как важные факторы для изучения экобиоморф и экологических ниш гидромакрофитов // Вестник Нижневартовского государственного университета. 2025. № 4(72). С. 29-42.
Sviridenko, B.F. (2025). Total Salt Content and Chemical Composition of Water as Important Factors for the Study of Ecobiomorphes and Ecological Niches of Hydromacrophytes. Bulletin of Nizhnevartovsk State University, 4(72), 29-42. (in Russ.).
Западно-Сибирская равнина является регионом, в котором факторы водной среды широко варьируют, что позволяет получить на этой территории количественную информацию об экологической толерантности гидромакрофитов. Минерализация (соленость) воды является одним из главных абиотических факторов для гидрофильных растений [4; 9; 13; 14; 15; 16; 23; 42]. Цель работы заключается в обосновании подходов к дифференциации экобиоморф гидромакрофитов на основе количественных данных о галотолерантности.
Для обитания разных видов гидромакрофитов пригодна как ультрапресная вода с очень низкой концентрацией растворенных минеральных веществ (до 0,1 г/дм3), так и в разной степени насыщенная солями. Каждый вид находит условия для своего существования только в определенной, нередко относительно узкой части общего диапазона минерализации поверхностных вод. Способность организмов (видов) жить в конкретном диапазоне минерализации вод получила название солевыносливости или галотолерантности. Это определение основано на законе толерантности В.Э. Шелфорда [18; 22; 24]. Известно, что растворенные в воде соли влияют не только на осмотические процессы в растительных клетках, но могут оказывать также токсическое действие, в связи с чем они представляют собой лимитирующий фактор. Поэтому выявление диапазонов галотолерантности, ограничивающих возможность существования континентальноводных видов, имеет большое научное значение в плане изучения экобиоморф и экологических ниш этих растений.

Под минерализацией воды понимают количество растворенных в ней минеральных веществ. В водных объектах вода не встречается в химически чистом виде, так как в ней растворены соли и другие вещества. Общая минерализация представляет сумму компонентов, найденных в ходе гидрохимического анализа. В число учитываемых компонентов обычно относят катионы калия (K+), натрия (Na+), кальция (Ca2+), магния (Mg2+) и анионы хлора (Cl-), сульфат-анион (SO 4 2-), гидрокарбонат-анион (HCO 3 -), тогда как другие ионы в этот показатель не входят [2; 21; 43]. При определении концентрации Ca2+ используют титриметрический метод с комплексоном III в присутствии индикатора мурексид, концентрации Cl- – аргентометрический метод, SO 4 2- – весовой метод, HCO 3 - – метод обратного титрования в присутствии смешанного индикатора (метиловый красный – метиленовый голубой). Содержание катионов магния (Mg2+) и одновалентных катионов (K+ + Na+) определяют путем расчета [3; 25; 36].
В современный период исследование ионного состава растворенных солей в воде проводят методом высокоэффективной жидкостной хроматографии на ионном хроматографе «Стайер» с кондуктометрическим детектором. Для разделения ионов используют хроматографические колонки: при определении катионов – Shodex IC YS-50, при определении анионов – TRANSGENOMIC ICSep AN2 [20]. Для определения массовой концентрации карбонат- и гидрокарбонат-ионов используют значения свободной щелочности и общей щелочности, применяя соотношения и расчетные формулы [6; 7]. Содержание хлорид-иона определяют титриметрически по реакции с азотнокислым серебром [8]. Определение содержания нитрат-, фосфат-, сульфат-ионов проводят по методике [26]. В итоге общую минерализацию воды определяют как сумму ионов. Также выявляется общая жесткость воды, количественно выражаемая как сумма двухвалентных катионов (Ca2+ + Mg2+, мг-экв/дм3), и состав основных растворенных солей.
Существенно сложнее определение абсолютной величины минерализации воды по ее электропроводимости в пересчете на NaCl, так как природные воды представляют собой сложную смесь электролитов. Хлоридные ионы, по данным М.Г. Валяшко [5], начинают преобладать только при минерализации более 10‰, однако в содовых и сульфатных гипергалинных озерах эти ионы также составляют лишь небольшую долю от общего состава. В то же время электропроводимость зависит в сильной мере от ионного состава, поэтому установление непосредственной связи между удельной электропроводимостью и концентрацией ионов в воде может быть сделано лишь со значительными погрешностями, а конечный результат чаще всего имеет неудовлетворительную точность [10]. Кроме того, метод не дает информации о составе растворенных солей и общей жесткости воды. Этот вывод наглядно демонстрируют материалы публикации [17], в которой для 27 водоемов приведены количественные данные о минерализации воды, полученные кондуктометрическим методом по электропроводимости (в пересчете на NaCl) и методами гидрохимического анализа. Большинство из этих водоемов (67%) имеют воду, не относящуюся по составу к хлоридно-натриевой согласно приведенным формулам М.Г. Курлова [17]. Полученные по электропроводимости значения минерализации воды составляют только 48–86% от реальных значений, полученных при гидрохимическом анализе. Таким образом, кондуктометрические измерения минерализации воды континентальных водных объектов существенно искажают данный показатель и не могут использоваться в гидроботанических исследованиях.
Во многих работах для оценки экологии морских видов растений принято приводить данные о солености воды в так называемых практических единицах солености (psu) или даже просто в единицах измерения электропроводимости воды (мкСм/см) без комментария об экологическом значении указанных величин. Но в разных частях акваторий внутренних морей состав основных ионов может сильно отличаться от океанического состава, при этом реальные значения солености сильно варьируют. Например, соленость вод Балтийского моря изменяется на разных участках от 2‰ до 20‰. Поэтому использование кондуктометрического метода оценки минерализации воды в подобных экосистемах также ведет к значительным ошибкам и неверным выводам относительно солевыносливости видов.
Материал и методы
В период 1984–2023 гг. на Западно-Сибирской равнине выполнялись работы по оценке параметров воды в экотопах водных макроскопических растений. Целью было выявление количественных диапазонов толерантности видов к ведущим параметрам водной среды. В 1984–1994 гг. сбор данных выполнялся в казахстанской части равнины. В экотопах изучалась концентрация основных ионов (Cl-, SO 4 2- , HCO 3 - , K+, Na+, Mg2+, Ca2+), общая минерализация, общая жесткость, активная реакция воды. Эти параметры водной среды выражают количественно на основе измерений, выполняемых с применением аналитических методик, что позволяет объективно сравнивать экологические требования видов растений и использовать их в экобиоморфологической классификации и определении границ экологических ниш. В начале работы исследования параметров воды выполнялись Т.В. Свириденко в Северо-Казахстанской комплексной гидробиологической лаборатории Казахского НИИ рыбного хозяйства. Минерализация воды определялась как сумма основных ионов, количественно определяемых в трехкратной повторности с точностью до 5% на основе общепринятых методик [2; 3; 21; 25; 36; 43].
В 1995-2023 гг. эти исследования были продолжены на российской территории в основном в пределах Омской, Курганской, Тюменской, Новосибирской областей и Ханты-Мансийского автономного округа – Югры. Анализ проб воды выполнен Ю.А. Мурашко в гидрохимической лаборатории НИИ экологии Севера и Центре коллективного пользования Сургутского государственного университета на основе современного аналитического оборудования по соответствующим методикам [6; 7; 8; 29; 26].
В итоге результаты были представлены в табличной форме, где для видов гидромакрофитов указаны выявленные диапазоны значений общей минерализации, общей жесткости и рН воды (минимальные и максимальные). Полученные данные позволяют оценивать режимы изменений этих факторов, в пределах которых возможно существование каждого вида. Предварительные материалы были опубликованы ранее [29; 31; 32; 33], однако эти исследования продолжаются, в связи с чем диапазоны значений факторов систематически уточняются. Количественная оценка толерантности видов уже послужила основой для разработки системы экобиоморф цветковых гидрофитов и харовых водорослей (Сharophyta) [27; 29; 34]. Кроме того, алгоритм количественной оценки параметров водной среды на основе информации о толерантности видов макроскопических водорослей и высших гидрофитов к минерализации, жесткости и активной реакции приведен в одной из работ [30].
Всего была получена информация разной степени полноты о диапазонах галотолерантности 247 видов гидромакрофитов Западно-Сибирской равнины. Для широко распространенных видов диапазоны галотолерантности выявлены весьма надежно, в связи с чем полученные в последние годы новые данные не приводят к изменению их граничных значений. Однако для редких видов, находки которых на территории Западно-Сибирской равнины весьма малочисленны или даже единичны, приведенные диапазоны (единичные значения) носят предварительный характер. При анализе этих данных отмечено, что отдельные пороговые значения минерализации являются лимитирующими для целых групп видов, в связи с чем для выражения их галотолерантности была принята следующая градация поверхностных вод (табл. 1).
Таблица 1
Подразделение поверхностных вод Западно-Сибирской равнины по минерализации
|
Группы вод |
Подгруппы вод |
Минерализация, г/дм3 |
|
Пресная |
ультрапресная |
до 0,2 |
|
типично пресная |
0,2–1,0 |
|
|
Условно-пресная |
условно-пресная |
1,1–3 |
|
Солоноватая |
слабосолоноватая |
3,1–5 |
|
среднесолоноватая |
5,1–8 |
|
|
сильносолоноватая |
8,1–25 |
|
|
Соляная |
соляная |
более 25 |
Соответственно, виды, отмеченные в диапазоне минерализации до 0,2 г/дм3, называем ультрапресноводными, до 1 г/дм3 – типично пресноводными, до 3 г/дм3 – условнопресноводными, до 5 г/дм3 – слабосолоновато-пресноводными, до 8 г/дм3 – среднесолоновато-пресноводными, до 25 г/дм3 – сильносолоновато-пресноводными, свыше 25 г/дм3 – соляноводными. Эти градации использованы при выделении экобиоморф гидромакрофитов наряду с другими экологическими параметрами (общая жесткость и рН воды).
Результаты и обсуждение
В составе водной макрофитной флоры Западно-Сибирской равнины выделено 2 совокупности видов, которые обозначены как пресноводный и соляноводный флористические комплексы [28; 29]. Естественной экологической границей между данными комплексами является узкий диапазон минерализации, получивший в гидробиологической литературе название критической солености [41]. Согласно концепции критической солености, в воде океанического состава соленость около 5–8 г/дм3 представляет собой универсальный барьер, разделяющий биологические процессы разной направленности или интенсивности. Особая роль барьера критической солености объясняется резкими изменениями в этом диапазоне физико-химических свойств среды. Критическая соленость является границей распространения организмов с различной осморегуляторной способностью. По обе стороны узкой зоны солености 5–8 г/дм3 по-разному протекают обменные процессы в организмах и тканях, а многие важнейшие макромолекулярные вещества имеют различную структуру [37; 41]. Барьерная роль солености 5–8 г/дм3 отчетливо проявляется только в водах с океаническим составом солей. В водах другого состава, например, в Аральском и Каспийском морях отмечено смещение критической солености в сторону более высоких концентраций и одновременное увеличение интервала между его крайними точками [1; 12]. Это явление имеет место и в поверхностных водах Западно-Сибирской равнины, отличающихся широким разнообразием по концентрации и составу солей. Поэтому обобщенная «зона стыка» видов пресноводного и соляноводного флористических комплексов в местных водоемах охватывает диапазон минерализации 8–25 г/дм3, то есть расположена в сильносолоноватых водах. Критическая соленость в пределах этой зоны сдвигается в ту или другую сторону в зависимости от состава воды в конкретном водоеме. Это явление связано с действием закона антагонизма ионов Ж. Леба, согласно которому токсическое влияние одновалентных катионов (K+ + Na+) существенно нивелируется при наличии в воде двухвалентных катионов (Mg2+ + Ca2+) [11; 44]. Как известно, в аридных и семиаридных районах Северной Евразии широко распространены среди минерализованных поверхностных вод не только хлоридно-натриевые, но также сульфатные и гидрокарбонатные воды с высоким содержанием двухвалентных катионов кальция и магния [38; 39; 40]. При минерализации меньше 8–25 г/дм3 отмечается максимум видов пресноводного комплекса, соответственно, при минерализации более 8–25 г/дм3 располагается максимум видов соляноводного комплекса. Виды из разных флористических комплексов редко формируют общие фитоценозы, поскольку критическая соленость (диапазон 8–25 г/дм3) является для них одинаково неблагоприятной. В водных объектах с такой минерализацией наблюдается низкопродуктивные популяции минимального числа видов.
Установлено, что на Западно-Сибирской равнине пресноводный флористический комплекс экологически неоднороден. Он подразделяется на группы: ультрапресноводную (23% видов), типично пресноводную (34%), условно-пресноводную (24%), слабосолоновато-пресноводную (6%), среднесолоновато-пресноводную (5%) и сильносолоновато-пресноводную (5%). Эти группы условны и между ними нет четких границ, поскольку существует экологический континуум. Соляноводный флористический комплекс (2,7%) представляют виды, не проникающие в пресные, условно-пресные и слабосолоноватые воды: Lamprothamnium papulosum (Wallr.) J. Groves, Percursaria percursa (Ag.) Bory, Ulothrix flacca (Dillwyn) Thuret, Ulothrix implexa (Kützing) Kützing, Ruppia maritima L. , Ruppia drepanensis Tineo и Althenia filiformis F. Petit subsp. orientalis Tzvel. (рис. 1). Особо выделяется эвригалинный вид Cladophora glomerata (L.) Kützing, отмеченный в очень широком диапазоне минерализации воды (0,10–79,70 г/дм3).
Согласно определению И.Г. Серебрякова [35], понятие «жизненная форма» содержит 3 аспекта: 1 – жизненная форма как основная таксономическая единица; 2 – жизненная форма как своеобразный общий облик (габитус) дефинитивных особей вида, включая надземные и подземные органы; 3 – жизненная форма как выражение приспособленности растений к определенным условиям абиотической и биотической среды.
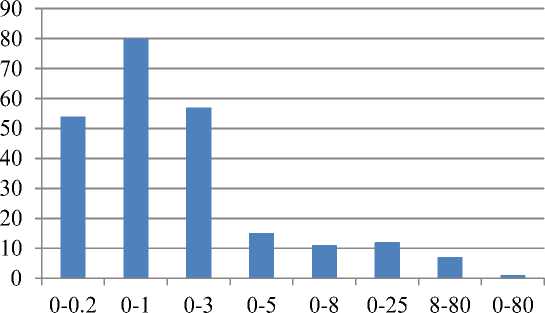
Рис. 1. Количество видов гидромакрофитов из разных галотолерантных групп.
По оси абсцисс – диапазоны галотолерантности (г/дм3), по оси ординат – количество видов
Жизненные формы (биоморфы) растений характеризуют на основе важнейших биологических и морфологических параметров видов, позволяющих кратко, но достаточно информативно описать их габитус. К таким признакам у цветковых гидромакрофитов принадлежат продолжительность большого жизненного цикла (многолетние, двулетние, однолетние), степень связи видов с грунтом как со средой обитания (укореняющиеся, свободноплавающие), типы и расположение зимующих почек в сочетании с особенностями структуры надземных побегов (длиннопобеговые корневищные, розеточные корневищные, длиннопобеговые клубневые, розеточные клубневые, длиннопобеговые столонные, длиннопобеговые турионовые укореняющиеся, розеточные турионовые, длиннопобеговые турионовые свободноплавающие, листецовые турионовые, длиннопобеговые без зимующих почек – гидротерофиты).
Также учитывается положение ассимиляционной поверхности относительно границы, разделяющей водную и воздушную среды (гелофиты, плейстофиты, гидатофиты), линейные размеры дефинитивных особей (высокие, средневысокие, низкие) и некоторые специфические черты биологии и морфологии, например, листорасположение (для длиннопобеговых), форма листовых пластинок и их развитость, наличие листовых ловчих камер, форма листецов, особенности цветения (надводно- и подводноцветущие) [27].
В определении понятия «жизненная форма» И.Г. Серебрякова [35] третий аспект акцентирует внимание на выявлении факторов, обеспечивших эволюционное формирование биоморфы конкретного вида, что в итоге позволяет установить специфику его экологической ниши. Это направление было дополнено в работах Е.М. Лавренко и В.М. Свешниковой [19], сформулировавших представление об экобиоморфах. Именно эта задача количественной оценки экологической индивидуальности каждого вида и его экологической ниши является основной при экобиоморфологических исследованиях. Экобиоморфологическая концепция имеет особенно решающее значение при изучении жизненных форм не только высших гидромакрофитов, но и многих континентальноводных макроскопических низших растений, у которых габитуальные признаки малоинформативны, в связи с чем системы жизненных форм еще не разработаны. Биоморфы близкородственных видов из некоторых родов высших гидромакрофитов также сложно дифференцировать на основании только биолого-морфологических признаков, поэтому количественные данные о галотолерантности имеют решающие значение для выявления их экобиоморф и экологических ниш.
Например, биоморфы видов из рода Myriophyllum – M. spicatum L. и M. verticillatum L. весьма сходны габитуально (рис. 2, а ), однако на основе выявленных значений галотолерантности их экобиоморфы различаются.
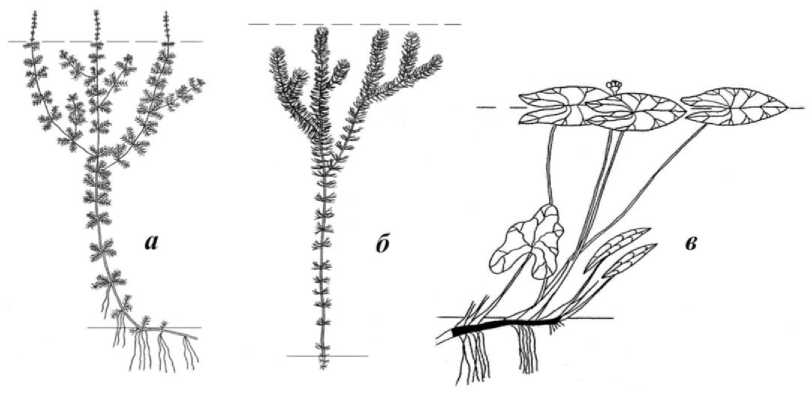
Рис. 2. Схемы биоморф видов: а – Myriophyllum spicatum и M. verticillatum; б – Ceratophyllum demersum и C. submersum; в – Nuphar lutea и N. pumila . Пунктиром показан уровень воды, сплошной линией – уровень грунта
В экобиоморфологической классификации вид M. spicatum – многолетний поликарпический длиннопобеговый турионовый укореняющийся высокий надводноцветущий слабосолоновато-пресноводный (диапазон минерализации 0,20– 4,10 г/дм3) гидатофит, вид M. verticillatum – многолетний поликарпический длиннопобеговый турионовый укореняющийся высокий надводноцветущий условнопресноводный (диапазон минерализации 0,02–2,70 г/дм3) гидатофит.
Другой пример относится к двум сходным по биоморфам видам из рода Ceratophyllum : С. demersum L. и C. submersum L . (рис. 2, б ), экобиоморфы которых различаются по галотолерантности. Вид Ceratophyllum demersum – многолетний длиннопобеговый турионовый свободноплавающий (бескорневой) высокий подводноцветущий условно-пресноводный (диапазон галотолерантности 0,03–1,60 г/дм3) гидатофит, тогда как C. submersum – многолетний длиннопобеговый турионовый свободноплавающий (бескорневой) высокий подводноцветущий слабосолоноватопресноводный (диапазон галотолерантности 0,23–5,10 г/дм3) гидатофит.
Существенно различаются по экобиоморфам сходные виды Nuphar lutea (L.) Smith и N. pumila (Timm) DC. (рис. 2, в ). Вид Nuphar lutea многолетний поликарпический корневищный розеточный высокий надводноцветуший условно-пресноводный (диапазон минерализации 0,03–1,69 г/дм3) плейстофит, тогда как N. pumila – многолетний поликарпический корневищный розеточный высокий надводноцветуший типично пресноводный (диапазон минерализации 0,01–0,90 г/дм3) плейстофит.
На этих примерах показано, что морфологически сходные виды занимают различные экологические ниши. В итоге выявленные количественные диапазоны минерализации для видов гидромакрофитов наряду с другими экологическими параметрами (состав ионов, общая жесткость, концентрация растворимых форм некоторых металлов, рН воды) обеспечивают объективное описание их экобиоморф. Установленные границы в континууме значений минерализации поверхностных вод Западно-Сибирской равнины разделяют экологические ниши видов растений и позволяют прогнозировать вероятный состав гидрофитоценозов на основе сведений о минерализации и составе воды в конкретных водных объектах. При исследовании экологических ниш видов представляют интерес не только верхние значения диапазонов, но также и нижние. Ультрапресноводные виды, отмеченные в очень узком диапазоне минерализации (0–0,2 г/дм3), в условиях Западно-Сибирской равнины чаще всего связаны с дистрофными водными объектами (внутриболотными озерами). В таких водах, относимых О.А. Алекиным [2] к четвертому типу, для которого характерно крайне низкое содержание гидрокарбонатного аниона (НСО 3 - = 0) активная реакция кислая. Кроме того, в этих водах отмечено общее низкое содержание других растворенных солей, в том числе источников биогенных элементов. Эти гидроэкотопы заселяют ацидотолерантные олиготрофные виды, так как они не пригодны для представителей типично пресноводных, условно-пресноводных и солоноватопресноводных групп.
Выводы
Количественные значения общей минерализации, установленные в ходе лабораторного анализа проб воды из экотопов видов, позволяют определить диапазон их солевыносливости. Наряду с другими факторами среды, значения которых оцениваются количественно (концентрация главных ионов и растворимых форм некоторых металлов, общая жесткость, активная реакция), установленные диапазоны галотолерантности видов служат научной основой для дифференциации их экобиоморф. Максимальные значения общей минерализации, выявленные для разных видов, представляют собой существенные границы, позволяющие определять их экологические ниши в отношении данного абиотического фактора.
Диапазон минерализации поверхностных вод Западно-Сибирской равнины, в пределах которого могут существовать гидромакрофиты, составляет от 0 до 80 г/дм3. Установлено, что этот экологический континуум занят двумя комплексами видов – пресноводным и соляноводным, которые разделены диапазоном минерализации 8–25 г/дм3. В пресноводном комплексе выделены группы ультрапресноводных, типично пресноводных, условно-пресноводных, слабосолоновато-пресноводных, среднесолоновато-пресноводных и сильносолоновато-пресноводных видов, для которых обозначены максимальные количественные границы галотолерантности. В соляноводном комплексе выделена группа видов, обитающих в диапазоне минерализации от сильносолоноватых до соляных вод. Особо выделяется эвригалинный вид Cladophora glomerata, отмеченный в очень широком диапазоне минерализации воды.
В пределах установленных групп диапазоны солевыносливости каждого вида также весьма индивидуальны, что позволяет на их основе дифференцировать экобиоморфы даже морфологических очень близких видов. При исследовании экологических ниш видов представляют интерес не только максимальные значения диапазонов галотолерантности, но также и минимальные. Виды соляноводного комплекса не проникают в пресные воды. Также отмечено, что ультрапресные кислые воды с низким содержанием гидрокарбонатного иона не пригодны для представителей типично пресноводных, условнопресноводных и солоновато-пресноводных групп видов.
