Общие закономерности синергических и антагонистических взаимодействий в радиобиологии

Автор: Евстратова Е.С., Гераськин С.А., Жураковская Г.П., Толкаева М.С., Петин В.Г.
Рубрика: Научные статьи
Статья в выпуске: 2 т.32, 2023 года.
Бесплатный доступ
Многие направления практической медицины используют комбинированное действие ионизирующих/неионизирующих излучений с гипертермией/химическими агентами, повышающими эффективность лучевой терапии. При этом, как показали многочисленные исследования, в результате таких воздействий могут регистрироваться как синергические, так и антагонистические эффекты. Целью работы является выявление общих закономерностей проявления синергизма и антагонизма при действии факторов разной природы на живые объекты. Представлены собственные экспериментальные данные, а также проанализированные результаты других авторов, свидетельствующие о разных типах взаимодействия одних и тех же воздействующих факторов. При этом эффект не зависит от объекта исследования и варьирует от синергизма до антагонизма в зависимости только от параметров агентов: мощности дозы, интенсивности, степени нагрева, концентрации химического препарата и др. Показано, что главные закономерности проявления синергических взаимодействий такие как: 1) синергизм максимален при одновременном действии агентов и зависит от соотношения повреждений, индуцированных каждым агентом; 2) существует оптимальное соотношение воздействующих факторов, при котором синергизм максимален; 3) понижение или повышение интенсивности одного из агентов требует пропорционального уменьшения или увеличения интенсивности второго агента для сохранения максимального синергического взаимодействия, актуальны и для антагонистического взаимодействия воздействующих факторов. Так, на бактериальных клетках в качестве примера антагонизма рассматривается взаимодействие гипертермии и глицерина. На этом же объекте представлены обширные данные по влиянию разных концентраций NaCl c ионизирующим излучением, ультразвуком и гипертермией, результат взаимодействия характеризовался устойчивым антагонизмом. Как глицерин, так и NaCl - вещества-осмолиты и их негативное действие может смягчаться при использовании средств, стабилизирующих систему осмотического гомеостаза. На дрожжевых клетках показано, как синергическое взаимодействие ультрафиолетового света и гипертермии плавно переходило в антагонистическое по мере снижения температурной нагрузки и отклонении параметров воздействующих факторов от оптимального. При этом эффекты антагонизма также характеризуются оптимальным соотношением доз применяемых агентов, при которых регистрируемый эффект имеет максимальное значение. Сделан вывод об общности основных закономерностей проявления эффектов синергизма и антагонизма при взаимодействии факторов разной природы независимо от биологического объекта, подвергающегося комбинированному воздействию.
Антагонизм, синергизм, общие закономерности, гипертермия, ионизирующее излучение, ультразвук, уф-свет, осмолиты, цисплатин, бактерии, дрожжи, клетки млекопитающих
Короткий адрес: https://sciup.org/170199708
IDR: 170199708 | УДК: 612.014.4:615.015.21:577.346 | DOI: 10.21870/0131-3878-2023-32-2-132-141
General patterns of synergistic and antagonistic interactions in radiobiology
Currently multimodal therapy is widely used for treatment of chronic diseases including cancer. The joint use of ionizing or on-ionizing radiation with hyperthermia or chemotherapeutic agents increases the effectiveness of the treatment. A response of living things to combination treatment may be synergistic, additive or antagonistic. The study is aimed to examine the common regularities of synergistic and antagonistic effects development following exposure of biological objects to various factors. The article presents analysis of experimental data received by the authors of the paper and the data published by other teams. It has been found that therapeutic effects of the same factors do not depend on the objects under study, it depends on the agents parameters: a dose rate, intensity, degree of heating (temperature), concentration of chemotherapeutic agents, etc. The common regularities of synergistic interactions are: 1) synergistic effect is maximum, if the agents act simultaneously, it depends on the ratio of injuries induced by each agent; 2) the optimal correlation between influencing factors provides the maximum synergistic effect; 3) in order to maintain maximum synergistic or antagonistic effects interaction, it is necessary to reduce or to increase the intensity of one of the agents and proportionally to increase or to reduce the intensity of the second agent. The combined effects of hyperthermia and glycerol on bacterial cells are considered as an example of the antagonistic effect. Exposure to different concentrations of sodium chloride, NaCl, in combination with ionizing radiation, or ultrasound, or hyperthermia forms a stable antagonistic effect. Both glycerol and NaCl are osmotic substances and their negative effects can be attenuated by means that stabilize the osmotic homeostasis system. Yeast cells shown the synergistic interaction of ultraviolet light and hyperthermia then it began smoothly passed into antagonistic interaction as the temperature load decreased and the parameters of the influencing factors deviated from the optimal. At the same time, the antagonistic effects are also characterized by the optimal ratio of doses of the agents used, at which the observed effect has the maximum value. The conclusion is made about the commonality of the main regularities of the manifestation of the effects of synergy and antagonism at the interaction of factors of different nature regardless of the biological object, which is subjected to combined influence.
Текст научной статьи Общие закономерности синергических и антагонистических взаимодействий в радиобиологии
Многие методы лечения онкологических и других заболеваний используют комбинации факторов различной природы [1-7], которые могут взаимодействовать друг с другом, усиливая или ослабляя конечный биологический и терапевтический эффект. В первом случае говорят о синергическом взаимодействии [8-10], во втором – об антагонистическом эффекте [11, 12]. Оба эти эффекта описаны в работе с диплоидными дрожжевыми клетками Saccharomyces cerevisiae [13], посвящённой роли интенсивности УФ-излучения в проявлении характера взаимодействия УФ-света
Евстратова Е.С.* – зав. отд., к.б.н. ФГБУ «НМИЦ радиологии» Минздрава России. Гераськин С.А. – зав. лаб., д.б.н., проф. ФГБНУ ВНИИРАЭ. Жураковская Г.П. – вед. науч. сотр., д.б.н.; Толкаева М.С. – инженер (эколог); Петин В.Г. – гл. науч. сотр., д.б.н., проф. МРНЦ им. А.Ф. Цыба – филиал ФГБУ «НМИЦ радиологии» Минздрава России.
с гипертермией. В этих экспериментах эффективность антагонистического взаимодействия была меньше по сравнению с синергизмом. Неоднозначные радиобиологические эффекты наблюдались в результате последовательного облучения гаплоидных или диплоидных дрожжей Saccharomyces cerevisiae УФ-излучением и α-частицами [14, 15]. Независимо от последовательности применения этих агентов результирующая выживаемость несколько раз проходит через максимумы и минимумы, демонстрируя то синергические, то антагонистические эффекты. Характер взаимодействия этих агентов был близок к аддитивному как для гаплоидного ( rad 51), так и диплоидного ( rad 51/ rad 51) радиочувствительных мутантов. Несколько другие результаты получены для УФ-чувствительного мутанта ( rad 18/ rad 18) [15]. При последовательном действии одной и той же дозы α-частиц и больших доз УФ-излучения регистрировали синергизм. В области малых доз УФ-облучения антагонизм проявлялся в виде плеча на кривой выживаемости, которая превращалась из экспоненциальной в сигмоидную. Однако максимальные значения антагонизма регистрировали при использовании химических агентов [16], которые легче проникали в бактериальные (прокариоты), но не дрожжевые (эукариоты) клетки, для которых наиболее были выражены синергические взаимодействия [10, 17-19]. Целью данной работы является выявление общих закономерностей проявления синергических и антагонистических эффектов на про- и эукариотических клетках.
Материал и методы
В опытах использовали диплоидные дрожжевые клетки Saccharomyces cerevisiae дикого типа (штамм XS800) и бактериальные клетки Escherichia coli (резистентный и радиочувствительный штаммы B/r и B s-1 ). В качестве источника ионизирующего излучения использовали электроны от 25 МэВ линейного импульсного ускорителя. Мощности доз 10, 25 и 250 Гр/мин определяли ферросульфатным методом [19]. Источником УФ-света служила 254 нм ртутная лампа при интенсивностях 0,033; 0,25 и 1,5 Вт/м2, которые были измерены гермицидным измерителем фирмы General Electric [13]. Облучение обоими источниками происходило одновременно с гипертермией (45-57,5 ºС). Водный раствор глицерина разных концентраций применяли при его одновременном действии с повышенными температурами. Раствор NaCl применяли одновременно с гипертермией, ультразвуком или ионизирующим излучением. Для расчёта выживаемости клетки высевали на чашки Петри с твёрдой питательной средой так, чтобы 150-200 колоний образовывались после 24-часовой инкубации бактерий при 37 ºС и 3-5-суточной инкубации дрожжевых клеток при 30 ºС. Результаты представлены в виде среднего значения и его стандартной ошибки.
Результат синергического или антагонистического эффектов оценивали коэффициентом эффективности комбинированного воздействия k, который определялся отношением изоэффек-тивных доз на кривых выживаемости после аддитивного сложения эффектов отдельно действующих факторов, и реально наблюдающихся после комбинированного воздействия этих факторов. Для синергизма коэффициент k всегда больше 1, для аддитивности он равен 1 и для антагонизма коэффициент k меньше 1.
Результаты и обсуждение
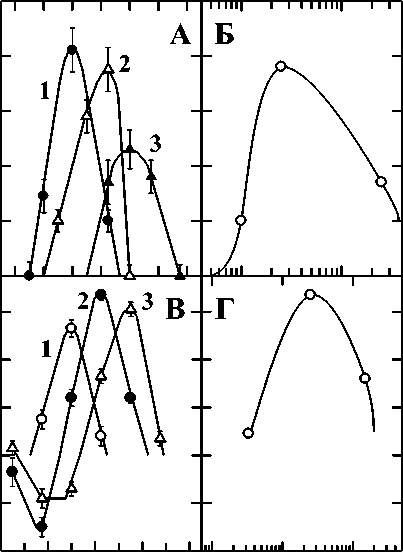
Для демонстрации общих закономерностей проявления эффектов синергизма, подробно описанных в работах [10, 19], на рис. 1 приведены результаты одновременного терморадиационного действия на диплоидные дрожжевые клетки Saccharomyces cerevisiae, штамм XS800. На панели А представлена зависимость коэффициента синергического усиления от температуры, при которой происходило облучение при разных мощностях доз ионизирующего излучения: кривые 1 – 10, 2 – 25, 3 – 250 Гр/мин. Видно, что для каждой мощности дозы имеется определённая температура, обеспечивающая максимальный синергический эффект, которая смещается в область более высоких температур с увеличением мощности дозы. На панели Б представлена зависимость коэффициента синергического усиления k от мощности дозы ионизирующего излучения при фиксированной действующей температуре 52,5 ºС. Эти результаты также указывают на существование оптимальной мощности туре 52,5 ºС.
2,0
0,6
Температура, оС 46 48 50 52 54 56 58
Мощность дозы, Гр/мин
10 100
1,8
0,8
46 50 54 58
Температура, оС
0,01 0,1 1
Интенсивность
УФ света, Вт/м2
2,0
1,8
1,6
1,4
1,2
1,0
1,6
1,4
1,2
1,0
0,8
0,6
Рис. 1. А, Б: Результаты одновременного терморадиационного действия на диплоидные дрожжевые клетки Saccharomyces cerevisiae , штамм XS800. А – зависимость коэффициента эффективности комбинированного воздействия k от температуры, при которой происходило облучение, и от мощности дозы ионизирующего излучения; кривые 1 – 10, 2 – 25, 3 – 250 Гр/мин; Б – зависимость коэффициента k от мощности дозы ионизирующего излучения при фиксированной действующей температуре (52,5 ºС). В, Г: Результаты одновременного действия ультрафиолетового излучения (254 нм) и гипертермии на диплоидные дрожжевые клетки Saccharomyces cerevisiae , штамм XS800. В – зависимость коэффициента k от температуры, при которой происходило облучение: 0,033 (кривая 1), 0,25 (кривая 2) и 1,5 Вт/м2 (кривая 3); Г – зависимость коэффициента k от интенсивности УФ-света при постоянной действующей температуре (52,5 ºС).
Аналогичные зависимости можно видеть на рис. 1В и 1Г для одновременного действия гипертермии и УФ света с разной интенсивностью: кривые 1 – 0,033, 2 – 0,25, 3 – 1,5 Вт/м2 [13, 17]. Представленные на рис. 1В данные интересны ещё и тем, что синергические эффекты, наблюдаемые при одних температурах, переходят в антагонистические при снижении температуры. При этом каждой из использованных в эксперименте интенсивностей УФ-света соответствует свой температурный диапазон, в котором наблюдается антагонизм и своя температура, максимизирующая этот эффект. Для мощности 0,033 Вт/м2 (рис. 1В, кривая 1) недостаточно данных для подтверждения антагонизма, однако есть теоретические расчёты, указывающие на возможность его существования. Т.е., как для синергизма, так и для антагонизма, при каждой интенсивности имеется определённая температура, обеспечивающая максимальный эффект. Эта температура смещается (увеличивается) в область более высоких температур для эффекта синергизма с увеличением УФ-интенсивности. Эти результаты указывают также на существование оптимальной интенсивности при облучении клеток при одной и той же температуре, например, 52,5 ºС или любой другой (рис. 1Г).
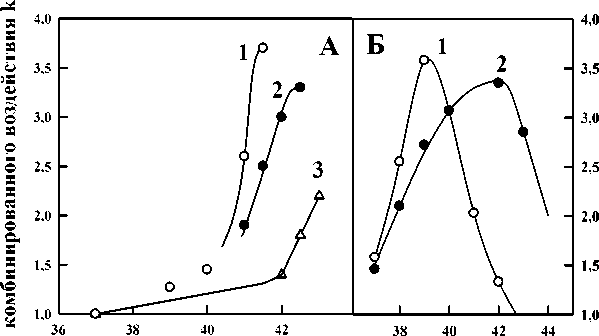
Чтобы подтвердить общность выявленных закономерностей, мы воспользовались данными других авторов [20-22], полученными для культивируемых клеток млекопитающих, которые сами авторы не применяли для расчёта эффективности синергического взаимодействия. На рис. 2А изображена рассчитанная нами зависимость коэффициента эффективности комбинированного воздействия k от температуры, при которой происходило облучение культивируемых клеток китайского хомячка CHO, и от мощности дозы ионизирующего излучения: кривые 1 – 0,033, 2 – 0,12, 3 – 3,6 Гр/мин. Хорошо видна описанная на предыдущем рисунке закономерность для дрожжевых клеток: с увеличением мощности дозы ионизирующего излучения зависимость синергизма от действующей температуры смещается в область более высоких температур. Рис. 2Б показывает зависимость коэффициента k от концентрации цисплатина: кривые 1 – 3, 2 – 6 мкМ, и от температуры, при которой происходило одновременное действие цисплатина и гипертермии на клетки китайского хомячка СНО. Видно, что и в этом случае сохраняются выявленные для дрожжевых клеток (рис. 1) закономерности – наличие действующей температуры, при которой эффект синергизма имеет наибольшее значение, и смещение куполообразной зависимости синергизма от температуры в область более высоких значений с увеличением концентрации цисплатина. Здесь заметим, что продолжительность действия химического вещества фиксированной концентрации можно рассматривать некоторым аналогом дозы, в то время как изменение концентрации отражает изменение интенсивности действия химических соединений. Аналогично, продолжительность действия фиксированной температуры можно также рассматривать в качестве аналога дозы, в то время как изменение величины температуры отражает изменение интенсивности нагрева.
Температура, оС
Температура, оС
Рис. 2. А: зави ры, при которой про дозы ионизирующег k от концентрации цисплатина; кривые 1 – 3, 2 – 6 мкМ, и температуры, при которой происходило одновременное действие цисплатина и гипертермии на клетки китайского хомячка СНО. По данным [20, 21].
Антагонистические взаимодействия хорошо изучены при защите термического поражения бактериальных клеток различными растворами солей. Посмотрим, выявляются ли при этом закономерности, аналогичные синергическим эффектам. На рис. 3А приведены экспериментальные данные о влияние водных растворов NaCl разной концентрации на выживаемость E. Coli B/r при воздействии гипертермии 52 ºС (продолжительность 15 мин). Данные на рис. 3Б – влияние водных растворов NaCl разной концентрации на выживаемость радиочувствительного мутанта бактерий E. coli B s-1 при воздействии гипертермии 52 ºС (продолжительность 15 мин). Видно, что в обоих случаях наблюдается куполообразная зависимость выживаемости клеток от концентрации хлорида натрия при его одновременном применении с фиксированной температурой. Это означает, что имеется оптимальное значение концентрации NaCl (5%), при которой оба штамма защищаются наиболее эффективно, т.е. проявляется максимальное антагонистическое взаимодействие гипертермии и раствора NaCl [16]. Эти данные показывают качественное сходство законом й й й
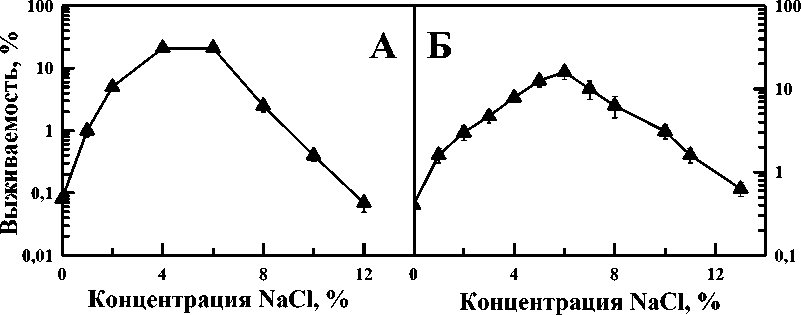
Рис.
гипертермии 52 С (продолжительность 15 мин). Б: влияние водных растворов NaCl различной концентрации на выживаемость бактерий E. coli B s-1 при воздействии гипертермии 52 ºС (продолжительность 15 мин) [16].
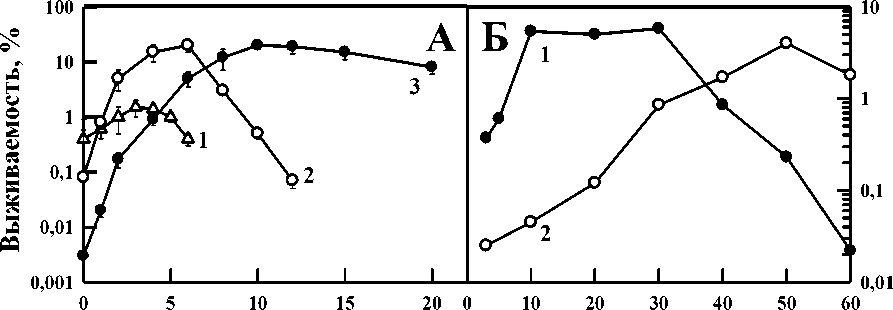
Следующая важная закономерность синергических взаимодействий заключается в зависимости синергизма от интенсивности применяемых агентов: при увеличении интенсивности действия ионизирующей радиации или УФ-света (рис. 1А и 1В) куполообразная зависимость синергизма от действующей температуры смещается в область более высоких температур. Для проверки этой гипотезы при антагонистических эффектах изучена зависимость выживаемости бактерий от действующей температуры и концентрации хлорида натрия или глицерина [16]. На рис. 4А изображена зависимость выживаемости бактерий Escherichia coli штамм B/r от концентрации NaCl в суспензионных средах в процессе термического воздействия при действии 50 (кривая 1), 52 (кривая 2) и 60 ºС (кривая 3). Видно, что независимо от действующей температуры в определённом диапазоне концентраций хлорида натрия наблюдается антагонистическое взаимодействие этих агентов, т.е. снижается повреждающее действие гипертермии. Характер антагонистического взаимодействия в процессе нагрева разной интенсивности значительно различается как по величине, так и по диапазону концентраций NaCl, в пределах которого оно регистрировалось. Видно также, что концентрационные максимумы антагонизма и куполообразная зависимость выживаемости от действующей температуры и концентрации хлорида натрия смещается в область больших концентраций этого соединения с ростом интенсивности теплового воздействия. Для подтверждения этой закономерности мы использовали также глицерин и СВЧ нагрев вместо традиционного нагрева. На рис. 4Б демонстрируется антагонистическое взаимодействие разных концентраций глицерина при анализе жизнеспособности бактерий Escherichia coli штамм B/r, прогретых СВЧ при 50 ºС в течение 3 ч (кривая 1) или при 60 ºС в течение 60 с (кривая 2). Видно смещение максимумов антагонизма в область больших концентраций глицерина при более интенсивном СВЧ прогреве. Именно такие
, не означает, что механизмы проявления синергизма и антагонизма аналогичны.
Концентрация NaCl, %
Концентрация глицерина, %
Рис. 4. А: зависимость выживаемости бактерий Е. coli В/гот концентрации NaCI в суспензионных средах в процессе термического воздействия при 50 (кривая 1), 52 (кривая 2) и 60 °C (кривая 3). Б: защитное влияние различных концентраций глицерина на жизнеспособность бактерий Е. coli В/r, прогретых СВЧ при 50 °C в течение 3 ч (кривая 1) или при 60 °C в течение 60 с (кривая 2) [16].
-
-
-й
-е у
- о соответствует экспериментальным данным (рис. 1 и 2).
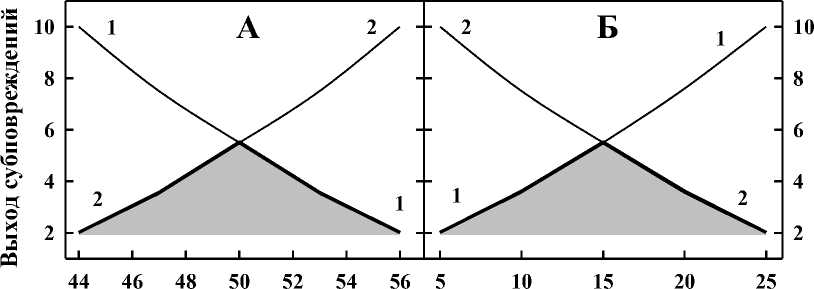
Температура, °С Мощность дозы, Гр/мин
-и о действия к от действующей температуры (А) и от мощности дозы ионизирующего излучения (Б).
Несколько другая ситуация наблюдается при действии фиксированной температуры и изменяющейся мощности дозы ионизирующего излучения (рис. 5Б) – число радиационных субповреждений возрастает с увеличением мощности дозы (кривая 1), а число термических субповреждений уменьшается (кривая 2). И в этом случае затемнённая область отражает зависимость коэффициента k от действующей температуры. Отметим, что в обоих случаях изменения выхода субповреждений оценивали для одной и той же выживаемости клеток.
Проанализируем возможную интерпретацию антагонистических взаимодействий. Рис. 3 демонстрирует такое взаимодействие гипертермии и хлорида натрия в зависимости от концентрации последнего для обоих штаммов Escherichia coli (B/r и B s-1 ). Видно, что эффект характеризуется наличием оптимальных концентраций, при которых отмечается максимальное антагонистическое взаимодействие. Было показано, что такие взаимодействия могут быть обусловлены изменением внутриклеточного осмотического давления [16]. Тогда с увеличением концентрации хлорида натрия (осмолита), повышающего осмотическое давление, происходит более эффективное антагонистическое взаимодействие гипертермии и хлорида натрия. Это возрастание имеет место до некоторого максимального (оптимального) значения концентрации хлорида натрия, наблюдающегося при изотонии – равенстве осмотического давления в суспензионной среде и клетках. Затем эффективность антагонизма снова уменьшается, что и наблюдалось в эксперименте – с увеличением концентрации осмолита в клеточной суспензии этот эффект исчезает, и, в конечном счёте, наблюдается термосенсибилизация клеток.
Антагонистические взаимодействия в процессе нагрева разной интенсивности резко отличаются от такого взаимодействия после действия одной фиксированной температуры. Из данных, представленных на рис. 4, следует, что величина антагонистического взаимодействия хлорида натрия или глицерина значительно увеличивается с ростом действующей температуры, а максимумы антагонизма сдвигаются в область больших концентраций этих соединений с ростом интенсивности действующей температуры. Мы уже проанализировали возможный механизм куполообразной зависимости антагонизма от концентрации осмолитов. Следует рассмотреть ещё возможную причину перемещения такой зависимости в область больших концентраций осмолитов NaCl и глицерина при повышении действующих температур. Для проявления антагонистического взаимодействия необходим определённый уровень насыщения водой клеток, ниже и выше которого они претерпевают воздействие гипертонического либо гипотонического стресса [16], о чём и свидетельствует куполообразная зависимость антагонизма от концентрации хлорида натрия или глицерина в суспензионной среде. При повышении действующей температуры эти стрессы ещё более усиливаются, и для их компенсации необходимо увеличивать концентрацию осмолитов, что и наблюдалось в эксперименте. Становится ясно, что система осмотического гомеостаза [23], наряду с ДНК, может рассматриваться в качестве чувствительной мишени при действии различных физических и химических агентов.
Таким образом, результаты данной работы убедительно показывают наличие общих закономерностей проявления синергических и антагонистических взаимодействий – оба эти эффекта имеют максимальное значение при определённом соотношении воздействующих агентов, которое смещается в область более высоких температур, концентраций химических агентов, мощностей доз или интенсивностей физических факторов для обеспечения максимального синергического или антагонистического взаимодействия.
Список литературы Общие закономерности синергических и антагонистических взаимодействий в радиобиологии
- Терапевтическая радиология: национальное руководство /под ред. А.Д. Каприна, Ю.С. Мардынского. М.: ГЭОТАР-Медиа, 2019. 704 с.
- Basic clinical radiobiology. Eds: M. Joiner, A. van der Kogel. London: CRC Press, 2009. 375 p.
- Chou T.-C. Drug combination studies and their synergy quantification using the Chou-Talalay method //Cancer Res. 2010. V. 70, N 2. P. 440-446.
- Chemoradiation in cancer therapy. Ed.: H. Choy. Totowa, NJ, USA: Humana Press, 2003. 420 p.
- Hahn G.M. Hyperthermia and cancer. New-York: Plenum Press, 1982. 285 p.
- Hall E.J., Giaccia A.J. Radiobiology for the radiologist. Philadelphia: Wolters Kluwer, 2018. 576 p.
- Streffer C., Vauper P., Hahn G. Biological basis of oncologic thermotherapy. Berlin, Heidelberg, New York, London, Paris, Tokyo, Hong Kong: Springer-Verlag, 1990. 169 p.
- Петин В.Г., Жураковская Г.П., Комарова Л.Н. Радиобиологические основы синергического взаимодействия в биосфере. М.: ГЕОС, 2012. 219 с.
- Petin V.G., Kim J.K. Synergistic interaction and cell responses to environmental factors. New York: Nova Sciences Publisher, 2016. 337 p.
- Evstratova E.S., Petin V.G., Zhurakovskaya G.P. Synergistic effects and their potential significance for the influence of natural intensities of environmental factors on cell growth //Synergy. 2018. V. 6, N 1. P. 1-8.
- Mitchel R.E.J., Morrison D.P. Heat-shock induction of ultraviolet light resistance in Saccharomyces cere-visiae //Radiat. Res. 1983. V. 96, N 1. P. 95-99.
- Barbe M.F., Tytell M., Gower D.J., Welch WJ. Hyperthermia protects against light damage in the rat retina //Science. 1988. V. 241, N 4874. P. 1817-1820.
- Petin V.G., Zhurakovskaya G.P., Komarova L.N. Fluence rate as a determinant of synergistic interaction of simultaneous action of UV-light and mild heat in Saccharomyces cerevisiae //J. Photochem. Photobiol. B: Biol. 1997. V. 38, N 2-3. P. 123-128.
- Calkins J., Ballard R., Gillespie M. Ultraviolet light-induced reactivation of alpha-irradiated yeast cells //Radiat. Res. 1978. V. 73, N 3. P. 440-451.
- Петин В.Г. Генетический контроль модификаций радиочувствительности клеток. М.: Энергоатомиздат, 1987. 208 с.
- Петин В.Г., Морозов И.И. Синергетика факторов окружающей среды. М.: ГЕОС, 2015. 249 с.
- Петин В.Г., Жураковская Г.П. Влияние интенсивности действующих агентов на проявление синергического взаимодействия //Радиационная Биология. Радиоэкология. 2015. Т. 55, № 6. С. 598-606.
- Петин В.Г., Белкина С.В., Жураковская Г.П. Математические модели и реакции клеток на облучение ионизирующими излучениями разного качества. М.: ГЕОС. 2020. 263 с.
- Petin V.G., Zhurakovskaya G.P. The peculiarities of the interaction of radiation and hyperthermia in Saccha-romyces cerevisiae irradiated with various dose rates //Yeast. 1995. V. 11, N 6. P. 549-554.
- Ben-Hur E. Mechanisms of the synergistic interaction between hyperthermia and radiation in cultured mammalian cells //J. Radiat. Res. 1976. V. 17, N 2. P. 92-98.
- Ben-Hur E., Elkind M.M. DNA damage and repair in hyperthermic mammalian cells: relation to enhanced cell killing //Radiation Research: Biomedical, Chemical and Physical Perspectives. New York: Academ Press, 1975. P. 703-717.
- Urano M., Kahn J., Majima H., Gerweck L.E. The cytotoxic effect of cis-diamminedichloro-platinum (II) on cultured Chinese hamster ovary cells at elevated temperatures: arrhenius plot analysis //Int. J. Hyperthermia. 1990. V. 6, N 3. P. 581-590.
- Панин Л.Е. Системные представления о гомеостазе //Бюллетень СО РАМН. 2007. Т. 127, № 5. С. 10-16.
