Оценка динамики численности и половозрастной структуры лося (A. alces L.) в России с использованием имитационной модели и данных многолетнего мониторинга
 в России с использованием имитационной модели и данных многолетнего мониторинга")
Автор: Петросян В.Г., Дергунова Н.Н., Бессонов С.А., Назарова К.А., Омельченко А.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Статья в выпуске: 1-7 т.11, 2009 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/148198521
IDR: 148198521 | УДК: 574.34:599.735.3
Evaluation of the number dynamics and sex-age structure of moose populations (Alces alces L.) in Russia
Текст статьи Оценка динамики численности и половозрастной структуры лося (A. alces L.) в России с использованием имитационной модели и данных многолетнего мониторинга
МЕТОДИКА ИССЛЕДОВАНИЙ
За последние 20 лет наблюдается значительное снижение численности лося как в отдельных регионах, так и в целом по России. Численность этого вида в России составляет 605 тысяч особей. По данным учета 2007 г. В скандинавских странах на меньшей в 11 раз территории обитает 550-600 тыс. лосей [1,2]. Ежегодная добыча вида в Феноскандии (Финляндия, Норвегия, Швеция) и России составляет 200-250 тыс. и 15-21 тыс. соответственно [2, 6-10]. Известно, что официальные данные по численности и добыче лося в странах Феноскандии, в частности в Финляндии, вполне надежные: случаев браконьерства в Финляндии практически нет, охотники, объединенные в общины, предоставляют объективные данные [6, 7]. С 1970 г. за состоянием популяции в Финляндии (особенно за плотностью и структурой населения) ведется постоянное наблюдение. Это обстоятельство позволяет утверждать, что модельное исследование финской популяции лося в сравнении с российской позволит выявить некоторые общие факторы, оказывающие влияния на динамику численности лося в России.
Цель работы - создание имитационной модели для выявления факторов, определяющих современную динамику численности и половозрастную структуру лося в России. В работе также подразумевается решение практической задачи - проведение сравнительного анализа динамики численности и параметров половозрастной структуры лося в России и Финляндии для определения оптимальных параметров рациональной эксплуатации ресурсов лося в России.
Разработана имитационная модель, которая предназначена для анализа динамики численности животных с учетом функциональных факторов (параметров плодовитости, жизнеспособности, смертности, официального отстрела, истребления хищниками и браконьерами, емкости угодий, доступности корма и др.), влияющих на те кущее состояние и динамику численности популяции. В модели рассматривается 18 возрастных групп особей двух полов. Каждая возрастная группа отличается от предыдущей группы на один год. Переменные, характеризующие чис ленность лосей в каждой из этих групп, форми руют вектор состояния системы (популяции):
X = ( X 1,1 , X 1,2 , X 2,1 , X 2,2
,....
, X 18,1 , X 18,2 ),
где X. 1 и X. 2 - численность самцов и самок соответственно в .-й возрастной группе. В началь ный момент времени моделирования (t = 0) по- ловозрастная структура определяется на основе литературных данных [11-23]. В модели предусмотрены возможности выбора параметров половозрастной структуры популяции по усмотрению пользователя (или на основе данных учетов при их наличии) в начальный момент t = 0. Далее определяется численность особей в последующий момент времени t + 1. В модели учитываются следующие параметры: репродуктивный статус особей в зависимости от возраста; параметры, указывающие доли стельных самок в популяции; количество эмбрионов на самку в зависимости от возраста; коэффициенты, характеризующие жизнеспособность особей в зависимости от их возраста; параметры лицензионного отстрела с учетом половой и возрастной структуры; влияние хищников (волка, медведя и др.) дифферен
цированно для различных возрастных групп; параметры, учитывающие неофициальный отстрел
животных и естественную смертность и др.
Рекуррентное соотношение, позволяющее определить количество особей для возрастных групп, имеет следующий вид:
X t + 1 = в • ( N 5 РХ 1 + N 2 ^ 2 р х 2 + N з 5 РХ з ) ;
X t + 1 = в • ( N 1 - (1 - p ) Х 1 + N . 5 . (1 - Р ) Х 2 + N з 5 з (1 - Р ) Х 3 )
X 2 + = X t ,1 « 1,1 - X t ,1 / 1,1 - X t А,;
X
t + 1
2,2
X 1,2 а 1,2 X 1,2 / 1,2 X 2,1 A 1,2 ’
t + 1 t t t t
X 3,1 X 2,1 a 2,1 X 2,1 / 2,1 X 2,1 ® 2,1 X 2,1 A 2,1 ;
X
t + 1
3,2
X 2,2 a 2,2 X 2,2 / 2,2
X 2,2 ® 2,2
2,1 2,2 ;
X 4Д = X 3 ,1 “ 3,1 - X 3,1 / 3,1 - X 3,1 ® 3,1 - X 3 JA3,1;
X
t + 1
4,2
X3,2 a 3,2 X 3,2 / 3,2 X3,2 ® 3,2 X 3,2 A 3,2 ’
t +1 t t t t л 18,1 ^17,1а17,1 ^-17,1 /17,1 ^17,1Ш17,1 ^17,1^17,1;
X 18,2 = X 17,2 a 17,2 - X 17,2 / 17,2 - X 17,2 ® 17,2 - X 17,2 A 18,2 ;
N 1 = min( S„S ... • a ); N . = min( S 2, S min • b ); N 3 = min( S 3 , S ... • c );
/ 18 18\
S min = min i s X t 1 , S X t 2 | ;
Vi=4 i=4
S 1 = S X t 2 ; S 2 = S X t 2 ;
i = 3
S a =-------1------;
S 1 + S 2 + S 3
i = 9
S 3 = S X '2 ; i = 13
b =---- S -----;
S 1 + S 2 + S 3
S c =------------;
S 1 + S 2 + S 3
где a .. - коэффициент выживаемости, т.е. перехода особей из одной возрастной группы в другую с учетом естественной смертности ( i = 1, 2...18; j = 1, 2). На основе литературных данных установлено, что значения a.. ( а. 1 = а. 2) могут быть представлены с помощью зависимостей a i = 1 . 10 - 5 i 3 - 0.0022 i 2 + 0.0262 i + 0.9171. Коэффициент в = в ( N , S , t ) — показатель плодовитости животных, который определяется в зависимости от N , S , t ; где N - общая численность животных в момент времени t , S - площадь обитания животных, га. В модели в определяется из следующе- 18 2
1000* SS X t j
го условия: i = 1 j = 1
S
<= n ; т. е. если в мо — р опт
мент времени t выполняется это неравенство, то величина в будет изменяться во времени согласно следующим значениям: вt+1 = 0,8; вt+2 = 0,5; вt+3 = 0,4; вt+4 = 0,8 (где ропт - допустимая плотность животных, экз. на 1000 га). Это условие обеспечивает ограничение плотности животных при значительном росте их численности на данной территории. Переменная у - коэффициент жизнеспособности особей в зависимости от плотности животных и возрастной группы (i = 1, 2.. .18; j = 1, 2), которая определяется с помощью соотношения у, = 7 •ю-6i4 - 0,0002i3 + 0,0031i2 - 0,0226i + 0,0598;
®.j - переменная, характеризующая неофициальное изъятие животных ( i = 3, 4^18; j = 1, 2);
A i , j - переменные, характеризующие изъятия животных хищниками ( i = 2,3,4^18; j = 1,2), p -половое соотношение новорожденных особей;
5 - случайная величина, определяющая количество эмбрионов на стельную самку ( i = 1^3); X i - доля стельных самок ( i = 1^3).
Численные эксперименты проводились с использованием созданной базы данных охотничье-промысловых видов России на основе многолетних данныхмониторинга (1981-2005 гг.) [8-10]. База данных создавалась сотрудниками ИПЭЭ РАН и Центрохозконтроля России в 2003-2009 гг. Данные учета численности Финской популяции лося брались из литературных источников [7].
Количество генераций, необходимых для получения значения показателей половозрастной структуры популяций лося, определялось помощью методов, представленных в работе [24].
АНАЛИЗ РЕЗУЛЬТАТОВ И ОБСУЖДЕНИЕ
Один из важных этапов использования разработанной модели - верификация параметров модели и проверка адекватности полученных результатов. В связи с тем, что многие параметры модели априори не известны, поэтому для подбора наилучшего варианта реализаций динамики численности значения параметров брались из наперед заданного допустимого интервала. Полученные случайные реализации сравнивались с данными мониторинга численности животных на основе метода наименьших квадратов (МНК), т.е. подбирались такие параметры модели, которые приводили к динамике численности лося, близкой к данным мониторинга. После анализа случайных генераций и оценки параметров адекватности была выбрана модельная реализация с точностью 90-92%. Результаты оценок динамики численности и половозрастной структуры лося на основе имитационной модели представлены на рис. 1 и 2.
Оценки, полученные с помощью имитационной модели показывают, что доля новорожденных особей меняется в диапазоне от 24 до 34% при среднем значении 27,8%, сеголеток -10-18% при среднем 12,42%, численность половозрелых особей в возрасте до 12 лет - от 3056,6% при среднем значении 52,4% (см. рис. 2).
Из оценок следует, что одна из важных причин низкой численности лосей в России - незначительный годовой прирост животных. Для понимания роли этого показателя в динамике численности лося мы использовали имитационную модель для оценки динамики численности лося
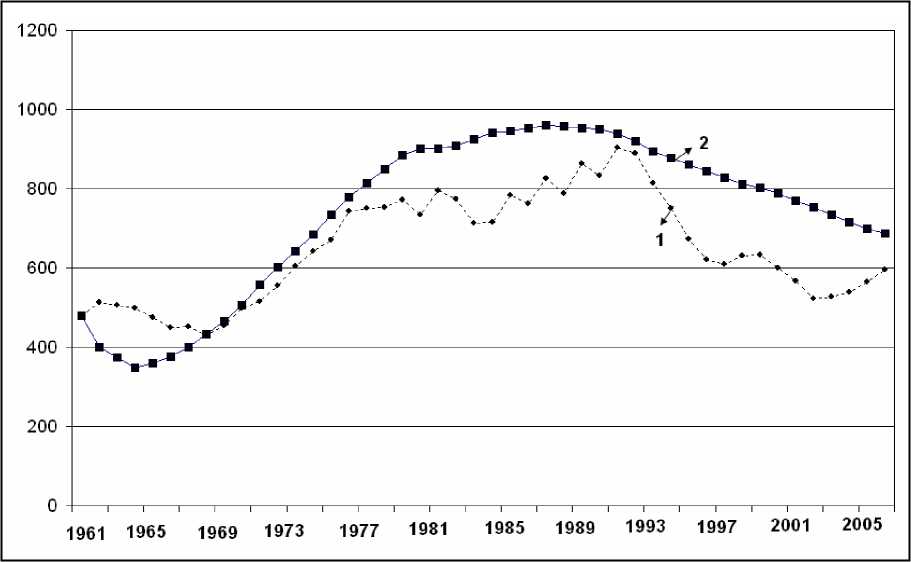
Рис. 1. Динамика численности лося в России: (1 - данные мониторинга, 2 - модельные оценки; по оси X - годы мониторинга, по оси Y - численность лосей в России, тысяч особей)
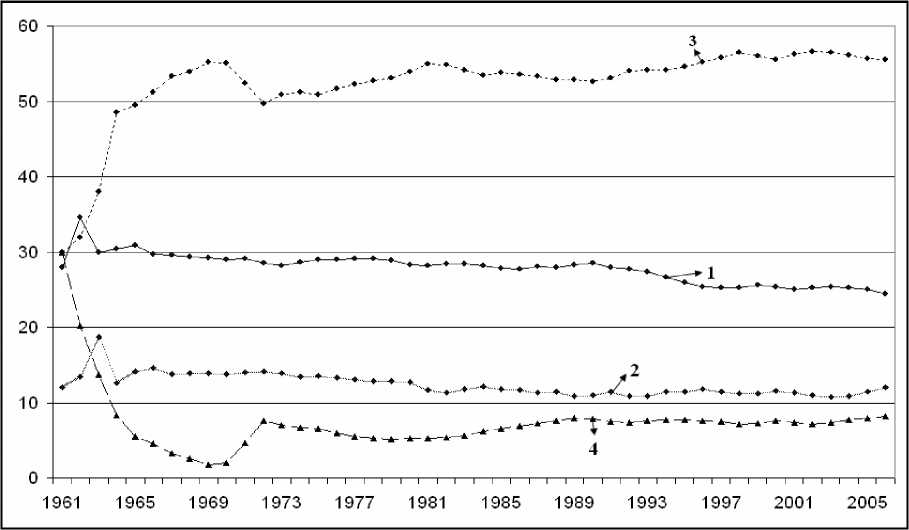
Рис. 2. Модельная оценка возрастной структуры лося в 1961-2005 гг.: (1 - новорожденные, 2 -сеголетки, 3 - взрослые до 12 лет, 4 - взрослые возраст у которых больше 12 лет; по оси X - годы, по оси Y - численность лосей для возрастных групп, тыс. особей)
в Финляндии на основе данных мониторинга в 1975-2005 гг. Полученные нами оценки показывают, что воспроизводство телят в финской популяции лося в 1975-2005 гг. изменялось в диапазоне 44-58%. Благодаря разумной методике по квотированию изъятия животных возрастная структура популяции поддерживалась на уровне, способствующем наибольшей плодовитости самок. Данные мониторинга указывают, что в популяции преобладали самки с высоким потенциалом плодовитости. Годичный прирост составлял около 60 телят на 100 взрослых особей [7].
Причем в 1981-2005 гг. финны изымали 56-75% поголовья предыдущей осенней послепромысло-вой численности (42-74 тыс. особей), за исключением периода 1994-1997 гг. В эти годы они изымали 22-32 тыс. особей. Одна из важных особенностей управления популяцией лося в Финляндии состоит в высоком уровня добычи сеголеток в каждый год изучаемого периода, после которой численность восстанавливалась в течение одного сезона. В этот период доля сеголеток составляла 39-54%. Данные мониторинга показывают, что стабильный рост численности лося достигался в период увеличения доли сеголеток в добыче при общем ее объеме, не превышающем уровень воспроизводства вида. Следует отметить, что такой высокий уровень добычи сеголеток практикуется также в Швеции и Норвегии [2, 6, 7]. Из проведенного сравнительного анализа следует, что, по-видимому, охотники и браконьеры в России ежегодно отстреливают преимущественно взрослых особей, при этом не только снижают репродуктивный потенциал вида, но и создают благоприятное условия для давления хищников на сеголеток-сирот. Кроме этого, в Финляндии охотники отстреливают преимущественно самцов, сохраняя соотношение полов 1,6:1 (т.е. 1,6 коров на одного быка) при плотности животных не ниже 1-2 особей на 1000 га. Для российской популяции лося аналогичные параметры значительно ниже и составляют 0,4-0,6 особей на 1000 га.
Хотя проведенный анализ динамики численности и половозрастной структуры лося в России и Финляндии позволяет сделать предварительный вывод относительно некоторых факторов, оказывающих влияние на сокращение численности диких копытных животных в России, тем не менее, потребуется дополнительный сравнительный анализ обеспеченности животных доступными кормовыми ресурсами в двух странах.
ЗАКЛЮЧЕНИЕ
Усредненные показатели воспроизводства телят в России 157 тысяч особей в год (модельная оценка) и легального отстрела 15-21 тыс. особей в год (официальные данные) свидетельствуют в пользу того, что современная динамика численности в значительной степени связана с влиянием естественных (увеличение численности хищников - волка) и антропогенных (браконьерство) факторов. Этот вывод дополнительно подтверждается данными мониторинга 19612005 гг. Известно, что в периоды максимальной численности лося в России (1991-1992 гг.) численность волка составляла порядка 22-24 тыс. особей. В последние годы с уменьшением чис ленности лося с 904 (1991 г.) до 565 (2006 г.) тыс. особей численность волка увеличилась примерно два раза и составляет 42-45 тыс. особей (данные 2006-2007 гг.). Для восстановления численности лося в России необходимо вводить дифференцированные лицензии на отстрел сеголетков и взрослых особей, преимущественно самцов. Причем, исходя из Финского опыта управления популяциями лося, рекомендуется соотношение полов 1,6:1 (т. е. 1,6 коров на одного быка) при плотности животных не ниже 1-2 особей на 1000 га. Это будет способствовать нормализации половозрастной структуры популяции и упорядочит отстрел. Нормирование добычи копытных должно выполняться с учетом параметров воспроизводства вида, плотности и половозрастной структуры животных, численности хищников и технологии изъятия в регионе. В целях сохранения основного воспроизводственного ядра копытных рекомендуется увеличить площади особо охраняемых природных территорий путем создания новых заповедников, заказников и других участков для воспроизводства лося.
В целом проведенный анализ показывает, что популяция лося в России находится в критическом состоянии. Если при сильной антропогенной и высокой техногенной нагрузке, при существующем уровне воспроизводства лося ситуация с динамикой численности будет развиваться в нынешнем направлении, то в предстоящие годы может произойти резкий спад численности животных.
Разработанная имитационная модель показала высокую эффективность для оценки демографических параметров популяции (выживаемости, смертности, плодовитости, прироста численности и др.) на основе данных мониторинга. Она может быть использована для определения демографических параметров других парнокопытных животных (косули, олени, кабана) с целью анализа их динамики численности. Имитационная модель и созданное программное обеспечение, реализующего модель, могут быть использованы для различных субъектов России с целью анализа динамики численности, на территории которых предполагается предоставление в пользование участков лесного фонда для организации и ведения охотничьих хозяйств различного направления.
Работа выполнена при финансовой поддержки программы фундаментальных исследований Отделения биологических наук РАН «Биологические ресурсы России: Фундаментальные основы рационального использования» (проект № II. 11) и частично гранта РФФИ (№ 09-04-91331_а).
