Оценка динамики растительности южной части Волго-Ахтубинской поймы

Автор: Бармин Александр Николаевич, Иолин Михаил Михайлович, Герасимова Ксения Анатольевна, Чувашов Андрей Викторович, Голуб Валентин Борисович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 5-2 т.18, 2016 года.
Бесплатный доступ
Проведен анализ повторных наблюдений (1961, 1973, 2010 гг.) на трансекте, и ключевых участках в южной части Волго-Ахтубинской поймы. Установлено, что наибольшая ксерофитизация растительного покрова и его пастбищная дигрессия в этом районе поймы были в 1973 г.
Волго-ахтубинская пойма, экологические шкалы л.г. раменского, dca-ординация, индикация
Короткий адрес: https://sciup.org/148204903
IDR: 148204903 | УДК: 633.2.03
Estimate of vegetation dynamics of the Volga-Akhtuba flood-plain (southern part)
The analysis of repeated observations (1961, 1973, 2010) in the transect and index sites in the southern part of the Volga-Akhtuba flood-plain has been carried out. Most vegetation xerophytization and pasture digression in this area of the flood-plain were identified in 1973.
Текст научной статьи Оценка динамики растительности южной части Волго-Ахтубинской поймы
В результате гидростроительства к началу 70-х годов прошлого века в бассейне р. Волги сохранился лишь один крупный регион с естественной пойменной растительностью. Это Волго-Ахтубинская пойма и дельта р. Волги. Существование здесь в зоне пустыни азональных сообществ с луговой, болотной и лесной растительностью обусловлено регулярными специальными попусками воды в нижний бьеф Волгоградского гидроузла. Искусственные попуски заменили естественные половодья, которые отличались большей продолжительностью и более высоким подъемом воды. Двумя другими важнейшими факторами, определяющими характер растительного покрова долины Нижней Волги, являются сенокошение и выпас скота.
Значительный интерес представляет периодическая оценка изменений растительности Волго-Ахтубинской поймы под воздействием антропогенных и природных факторов с целью научного обоснования мероприятий, обеспечивающих сохранение уникальной системы Нижней Волги [16].
МЕТОДИКА ИССЛЕДОВАНИЙ
В 1958-1961 гг. геоботаническая партия Всесоюзного аэрогеологического треста (ВАГТ) проводила картирование растительности долины Нижней Волги. В этот период в южной ее части были заложены 3 ключевых участка и трансекта. Участки располагаются у ерика Хора (85 га), близ ильменя Круглый Чураков (49 га) и в урочище Пастухова тоня (64 га). Трансекта пересекает пойму, начинаясь в районе станции Досанг и заканчиваясь вблизи села Петропавловка (рис. 1). Ее длина 10,3 км. Трансекта и ключевые участки были заложены до постройки Куйбышевского гидроузла, работа которого в наибольшей степени влияет на гидрологический режим низовий р. Волги [1].
На участках была проведена топографическая съемка, по результатам которой составлены карты масштаба 1:2000. Положение трансекты отражено на аэрофотоснимках. На ее линии было проведено нивелирование и вычерчен профиль с горизонтальным масштабом 1:5000. На всех топографических материалах были отмечены места расположения пробных площадок, на которых делали геоботанические описания.
Сохранились полевые дневники Л.С. Родман и И.Н. Горяиновой с геоботаническими описаниями пробных площадок на трансекте и ключевых участках. В дневниках имеется подробная характеристика мест геоботанических описаний и сами геоботанические описания, включающие список видов растений с указанием их обилия.
В середине 70-х годов прошлого века часть площади, пересекаемой трансектой, была обвалована и превращена в орошаемую пашню. Длина этой части трансекты - 2,1 км. Геоботанические описания, сделанные ранее на обвалованной площади, были исключены из рассмотрения.
После окончания работы геоботанической партии ВАГТ участки и трансекту посещали в
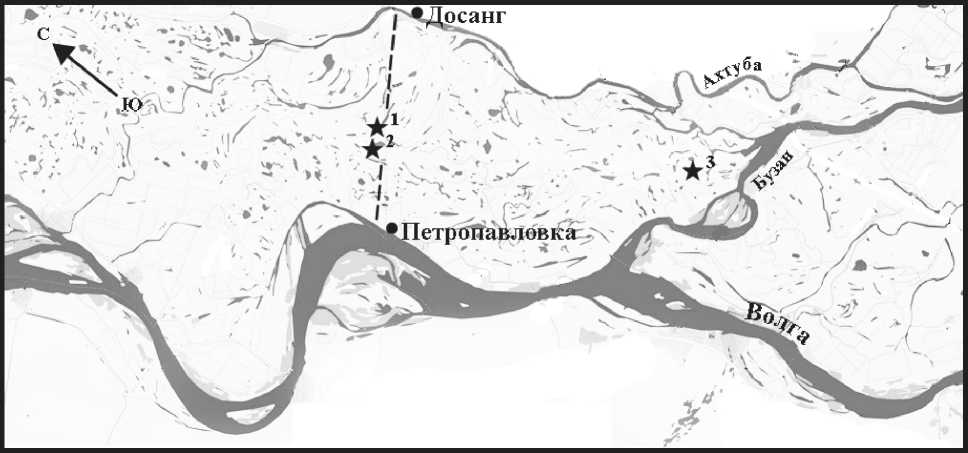
Рис. 1. Схематическая карта южной части Волго-Ахтубинской поймы.
Пунктиром обозначено расположение трансекты, звездочками - ключевые участки (1 - у ерика Хора, 2 - близ ильменя Круглый Чураков, 3 - в урочище Пастухова тоня)
1972, 1973, 1981, 1999 и 2010 гг. При подготовке данных для анализа выяснилось, что наибольшее число полностью сопоставимых учетных площадок на ключевых участках и необвалованной части трансекты имеется в 1961, 1973, и 2010 гг. На трансекте их было - 25, на ключевом участке у ер. Хора - 17, у ильменя Круглый Чураков - 21, Пастуховой тони - 16. Поэтому далее мы сопоставляем геоботанические описания только за эти годы. Всего учетных площадок, на которых все эти три года осуществляли описания, было 79, а общее количество геоботанических описаний, включенных в обработку - 237. Описания 1961 г. были неполными, поэтому при анализе динамики флоры и растительности их не учитывали.
Геоботанические описания в 1961 г. были проведены 14.08-16.08 и 18.10-21.10, в 1973 г. -29.07-05.08 и в 2010 г. - 09.08-16.08.
Все используемые в данной статье описания учетных площадок представлены в геоботанической базе данных долины Нижней Волги [13].
Перед обработкой из описаний были удалены виды рода Cuscuta, так как геоботаники не всегда их отмечали и правильно определяли. Некоторые виды растений, которые плохо различали между собой, были объединены в агрегации (agr.), а также понимались в широком смысле (s.l.) или как сумма таксонов.
Alisma lanceolatum + A. plantago aquatica
Scirpus maritimus s.l. = S. m. ssp. maritimus + Bolboschoenus glaucus
Carex acutiformis + C. melanostachya Eleocharis palustris + E. uniglumis
Euphorbia esula s.l. = Euphorbia esula ssp. tommasiniana + Euphorbia esula ssp. esula
Polygonum arenarium ssp. pulchellum = P. arenarium + P. patulum + P. aviculare + P. bellardii + P.
neglectum + P. patulum + P. salsugineum
Scutellaria galericulata + S. hastifolia
Названия видов растений даем по их списку в базе «Flora Europaea» [26], помещенной на сайте Эдинбургского королевского ботанического сада:
Обилие растений указываем в баллах: менее 1% проективного покрытия – +;1–5% – 1; 6–15% – 2; 16–25% – 3; 26–50% – 4; более 50% – 5. К числу растений-доминантов формально относили виды растений, проективное покрытие которых превышало 15%.
Для расчета экологических ступеней по шкалам Л.Г. Раменского (увлажнения, богатства и засоленности почвы, пастбищной дигрессии) применяли «метод пересечения большинства интервалов» [17]. Последний основан на определении моды в статистическом ряду распределения чисел, входящих в интервалы ограничительных ступеней экологических шкал [12]. При этом методе результаты индикации факторов среды не слишком сильно зависит от длины флористического списка. Поэтому в обработку по шкалам Л.Г. Раменского мы включили и геоботанические описания 1961 г., имевшие сокращенные флористические списки.
Сравнение распределения совокупности описаний по ступеням показателей шкал Л.Г. Раменского, рассчитанных для каждого года наблюдений, проводили вначале по тесту Краскела-Уоллиса. Решался вопрос: относятся ли сопоставляемые группы к одной или к разным генеральным совокупностям? Когда нулевая гипотеза не подтверждалась (т. е., сопоставляемые выборки относились к разным генеральным совокупностям), сравнение выборок произвели попарно с использованием теста Манна-Уитни [4, 5].
Широко использовали пакет программ JUICE 7.0. [27, ].
Дополнительно к шкалам для выявления направления изменений растительности применили DCA-ординацию геоботанических описаний с помощью встроенного в пакет программ JUICE модуля «Ordinations», взятого из программного пакета R-project [29].
Выделение групп растительных сообществ осуществляли путем кластерного анализа на основе расчета евклидова расстояния, примененного к количественным данным, и связывания кластеров методом Варда (Ward’s method). Расчеты проведены с помощью программы РС-ORD 5.0 в среде JUICE 7.0. [27]. Уровень кластеризации был выбран в результате определения значения максимальной «четкости классификации» [21].
В каждой выделенной группе мы устанавливали верные виды [22]. Верность измеряли phi-коэффициентом, расчет которого предложили чешские геоботаники [23]. Поскольку на величину phi-коэффициента влияет размер групп, предварительно их выравнивали [28]. После стандартизации phi-коэффициент в целевой группе может достигнуть высокого значения даже в случаях, когда верность определенного вида к этой группе статистически несущественна. Поэтому в дополнение к phi-коэффициенту для каждого вида группы вычисляли статистическое значение верности, используя критерий Фишера [23]. Всю процедуру расчетов phi-коэффициента и критерия Фишера реализовывали с помощью пакета программ JUICE 7.0. Величину phi-коэффициента, выше которой таксон относили к верному, определяли субъективно, с расчетом, чтобы количество таких таксонов было не слишком маленьким и не слишком большим. Это число было одинаковым для всех групп и равнялось 0,25.
При всех статистических оценках величины считали достоверными, если p-значение соответствующей статистики не превышало уровень значимости 0,05.
Данные о метеорологических и гидрологических факторах получены в Астраханской гидрометеообсерватории. За объем половодий мы условно принимали сток воды в створе Волгоградской ГЭС в течение второго квартала, когда проводят специальные попуски воды в нижний бьеф гидроузла [14].
Косвенно о пастбищной нагрузке судили по количеству поголовья скота в зоне долины Нижней Волги. Это вся Астраханская область и три южных района Волгоградской области (Ленинский, Светлоярский и Среднеахтубинский). Сведения о поголовье скота получены в органах статистики Астраханской и Волгоградской областей.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Характеристика экологических факторов . В годы учетов минимальный объем попуска воды в нижний бьеф Волгоградского гидроузла был в 1973 г. Это привело к небольшому уровню подъема воды во время половодья в 1973 г. (табл. 1), что не обеспечило полноценного затопления поймы. Кроме того, десятилетие, предшествовавшее 1973 г., также отличалось минимальным стоком, как, в среднем, за год, так и за период половодья (табл. 2). Следует обратить внимание на постоянное нарастание в последние десятилетия увлажнения поймы за счет осадков в теплый период года и связанное с этим увеличение гидротермического коэффициента. Это фактор, хотя и в небольшой мере, компенсирует ухудшение увлажнения долины Нижней Волги, которое произошло за счет сокращения объемов половодий после зарегулирования водного стока.
Что касается пастбищной нагрузки, то в десятилетия, предшествующие учетам в 1961, 1973 и в 2010 гг., в период 1964-1973 гг. в Волго-Ахтубинской пойме выпасалось максимальное количество скота.
В последние годы в южной части Волго-Ах-тубинской поймы стали создаваться буровые установки для разведки и добычи нефти. Они окружены валом, защищающим их от половодий, по валам же к ним подведены автомобильные дороги. Одно из таких сооружений, занимающее около 4 га, с координатами 46°53’09’’ с.ш. и 47°49’11’’ в.д. находится в 130 м от участка у ерика Хора, в 250 м от участка у ильменя Круглый Чураков и в 220 м от линии трансекты.
Флористический состав. При анализе флористического состава геоботанических описаний
Таблица 1. Метеорологические и гидрологические показатели в годы учетов на ключевых участках и трансекте
|
Год |
Объем водного стока в створе Волгоградской ГЭС, км3 |
Максимальный уровень подъема воды по рейке водомерного поста в г. Астрахань |
Сумма осадков за I-VII месяцы (гидрометобсерватория Астрахань), мм |
|
|
за год |
за второй квартал |
|||
|
1961 |
228 |
120 |
246 |
69 |
|
1973 |
174 |
78 |
230 |
181 |
|
2010 |
210 |
91 |
263 |
131 |
Таблица 2. Среднемноголетние показатели экологических факторов
|
Годы |
Сумма осадков, мм |
U cti cti ф £ ф н t^ cti m о Ч о |
и О Ф Ф ^ О \о и cti и t^ cti m о Ч о |
о к д я 0J S ЯГ S „ дя >• "& о m я S я « Я :Я Я Я Ч X 0J о и . у н 2 ^ аз д о ч Я |
Объем водного стока в створе Волгоградской ГЭС, км3 |
2 Ч о . и ^ Я И 2 д о с - К О Q Д о ~ я Я д % а я о $ х о а Sou д и < Я Д V 5 S g я cti |
и о д ^ ® S Ч й д О^о Ч Я X аз м и Я g О ° g ” у Й д 3 о она о я 2 2 § § ё S о аз X X |
||
|
за год |
за период с t°C более 10°С |
за год |
за второй квартал |
||||||
|
1952-1961 |
193 |
101 |
9,6 |
3696 |
0,27 |
247 |
114 |
270 |
224 |
|
1964-1973 |
199 |
104 |
9,7 |
3554 |
0,29 |
208 |
100 |
244 |
323 |
|
2001-2010 |
222 |
133 |
11,5 |
4128 |
0,32 |
253 |
104 |
275 |
189 |
Примечание: температуры и осадки приводятся по данным гидрометобсерватории в г. Астрахань
за 1973 и 2010 гг. в первом из этих двух лет учета было выявлено отсутствие самых обычных для Волго-Ахтубинской поймы гигрофитов Butomus umbellatus, Alisma lanceolatum + A. plantago-aquatica. В 2010 г. из числа пустынных эфемеров полностью отсутствовал Descurainia sophia, а Alyssum desertorum был представлен гораздо реже, чем в 1973 г. Эти явления объясняются разной мощностью половодий 1973 г. и 2010 г. Плохое затопление поймы 1973 г. привело к выпадению Butomus umbellatus, Alisma lanceolatum + A. plantago-aquatica. В то же время, более мощное половодье 2010 г., обеспечившее высокий уровень подъема воды, вероятно, уничтожило остатки эфемеров, которые развиваются до начало попусков воды в нижний бьеф Волгоградского гидроузла. Эта же причина вызвала более высокую встречаемость в 2010 г. Xanthium strumarium, плоды которого разносятся водой. Меньшую встречаемость легкоразруша-емых эфемеров в 2010 г. можно также отчасти объяснить более поздним посещением участков и трансекты, чем это было осуществлено в 1973 г.
Показатели шкал Л.Г. Раменского. Совокупность показателей увлажнения за 2010 г. достоверно отличалась от 1973 г. и не отличалась от 1961 г.; в 2010 г. увлажнение учетных площадок было выше, чем в 1973 г.
Шкалы Л.Г. Раменского диагностируют наименьше влияние выпаса на растительность в 1961 г. Совокупность показателей ступеней пастбищной дигрессии в этом году достоверно отличаются от 1973 г. и 2010 г. Максимальных значений эта дигрессия на отдельных учетных площадках достигала в 1973 г. (табл. 3, рис. 2 ПД).
Совокупность показателей богатства и засоления почвы в 2010 г. достоверно отличалось от таких же совокупностей в 1961 г. и 1973 г. Среднее и медиана выборки 2010 г. были несколько выше, чем в предыдущие годы учетов. Но, судя по рис. 2 БЗ, наиболее заметно уменьшение дисперсии этого показателя в 2010 г. в сравнение с показателями 1961 г. и 1973 г.
DCA ординация . Обращаясь к результатам DCA-ординации (рис. 3), можно сказать, что лучше всего удается интерпретировать первую ее ось. Она отражает комплексное влияние на флористический состав растительных сообществ двух факторов: увлажнения и пастбищной дигрессии (табл. 4). Вдоль этой оси вправо уменьшается увлажнение почвы на учетных площадках и увеличивается влияние выпаса. Такое взаимодействие
Таблица 3. Достоверные (+) и недостоверные (-) различия распределения учетных площадок по ступеням шкал Л.Г. Раменского, оцененные тестом Манна-Уитни
|
Увлажнение |
||
|
Год |
1973 |
2010 |
|
1961 |
+ |
|
|
1973 |
+ |
|
|
Пастбищная дигрессия |
||
|
Год |
1973 |
2010 |
|
1961 |
+ |
+ |
|
1973 |
- |
|
Богатство и засоление почвы
|
Год |
1973 |
2010 |
|
1961 |
- |
+ |
|
1973 |
+ |
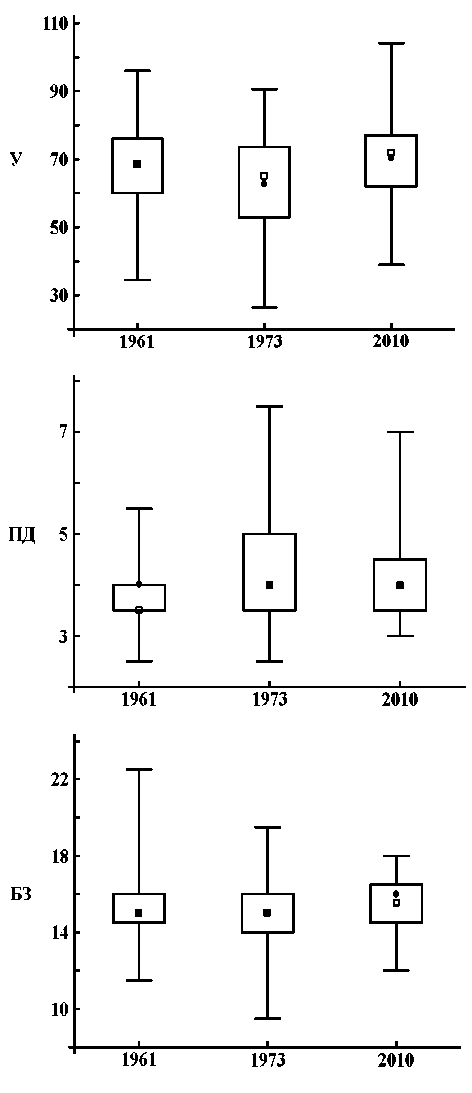
Рис. 2. Статистические параметры распределения учетных площадок по ступеням шкал Л.Г. Раменского в разные годы: У — увлажнение, ПД — пастбищная дигрессия, БЗ — богатство - засоления почвы.
Значения: - минимальное и максимальное,
□ - верхние и нижние квартили, ■ - медиана, • - среднее арифметическое факторов вполне объяснимо: чем выше экотопы над меженью реки, тем они суше. Они раньше освобождаются от воды и больше подвержены воздействию сельскохозяйственных животных. Средние значения проекций точек геоботанических описаний в разные годы на эту ось были обратно пропорциональны объему половодий (табл. 5). Максимальная дисперсия проекций гео- ботанических описаний была в 1973 г. Т.е., можно говорить о том, что наибольшая разнородность местообитаний по увлажнению и пастбищной нагрузке в южной части Волго-Ахтубинской поймы была в 1973 г.
Проекции описаний на вторую ось в 1961 и 1973 гг. также коррелируют со ступенями увлажнения, но с меньшими значениями, чем на первую и с противоположным знаком. Следует заметить, что в 1973 г., когда пастбищная нагрузка в годы учетов была максимальная, вторая ось ординации, наряду с первой, отразила достоверное влияние этого фактора.
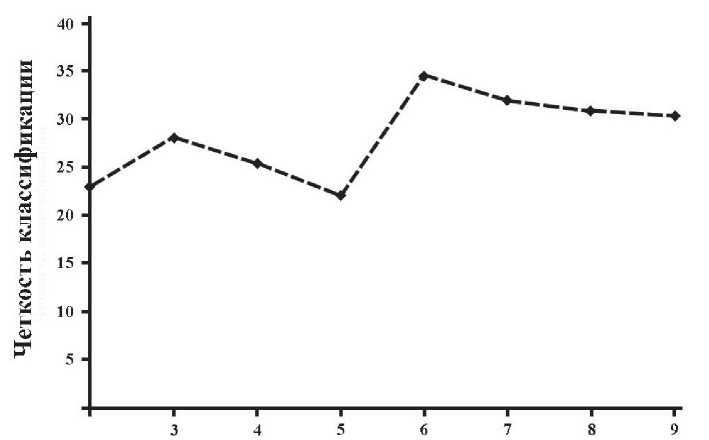
Сообщества . Максимальное значение четкости классификации общей выборки геоботанических описаний за 1973 и 2010 гг. (158 описаний) достигается при ее расчленении на 6 групп (рис. 4). В табл. 6 эти группы, в основном, расположены в вдоль двух связанных между собой градиентов: увлажнения и пастбищной дигрессии. Справа расположены более сырые местообитания с меньшей пастбищной дигрессией, слева – более сухие, с большей пастбищной дигрессией.
Первая группа - это наиболее ксерофитное сообщество (среднестепного увлажнения) с сильно выраженным влиянием выпаса скота. Занимает вершины невысоких грив и выровненные повышенные участки поймы. Диагностическими таксонами являются ксерофитные растения: Descurainia sophia, Alyssum desertorum, Bromus tectorum, Eremopyrum triticeum, Artemisia austriaca, Salsola kali, Bassia hyssopifolia, Dodartia orientalis, Acroptilon repens, Glycyrrhiza glabra, Calamagrostis epigejos. Среди перечисленных видов, первые четыре - это пустынные эфемеры. Пять следующих индицируют повышенную пастбищную дигрессию, а Bassia hyssopifolia - дополнительно и засоление почвы. В 2010 г. данный фитоценон был в 6 раз реже представлен, чем в 1973 г., когда Волго-Ахтубинская пойма была плохо затоплена во время половодья, а луга были подвержены особенно интенсивному пастбищному использованию.
Вторая группа - это местообитания сухолугового увлажнения с умеренным влиянием выпаса, представленные преимущественно на тех же экотопах, что и предыдущая группа. Диагностические виды этого сообщества Glycyrrhiza glabra и Calamagrostis epigejos. В число доминантов входит первый из этих двух видов. Рассматриваемый фи-тоценон был представлен на 4-х площадках в 1973 г. и на 25 - в 2010 г. В основном, рассматриваемый фитоценон заместил в 2010 г. сообщество первой группы. Это явление индицирует уменьшение ксерофитизации высоких пойменных местообитаний и сельскохозяйственной эксплуатации лугов 2010 г. в сравнении с 1973 г. О последнем говорит также высокое обилие в 2010 г. Glycyrrhiza glabra - вида, который плохо переносит ежегодное удаление надземной массы.
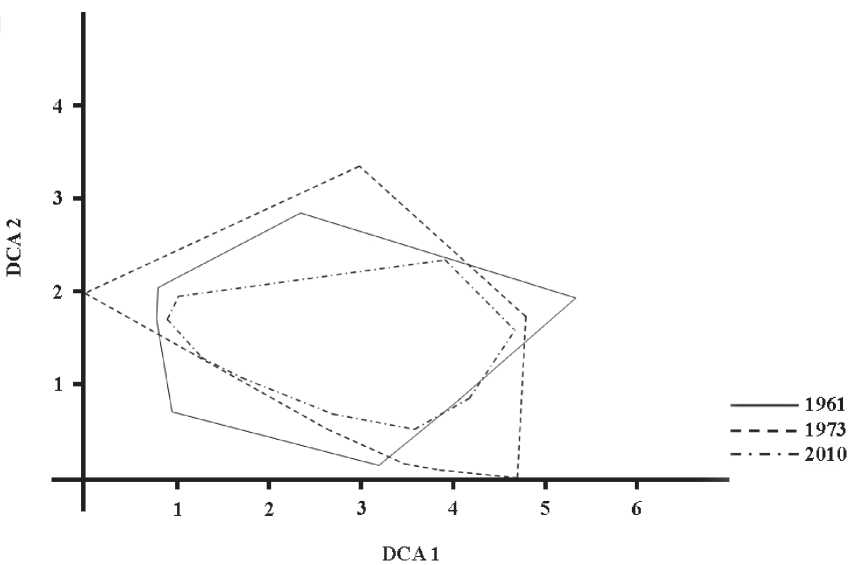
Рис. 3. Диаграмма DCA-ординации геоботанических описаний учетных площадок: линии обводят внешние границы «облаков» описаний, сделанных в разные годы. Собственное значение осей, характеризующее долю общей информации: ось 1 = 0,49, ось 2 = 0,23
Таблица 4. Коэффициенты корреляции, рассчитанные между значениями координат геоботанических описаний вдоль 1-й и 2-й оси DCA-ординации и ступенями шкал Л.Г. Раменского
|
Год |
1961 |
1973 |
2010 |
Все годы учетов |
||||
|
DCA1 |
DCA2 |
DCA1 |
DCA2 |
DCA1 |
DCA2 |
DCA1 |
DCA2 |
|
|
Шкала увлажнения |
-0,77 |
0,40 |
-0,78 |
0,67 |
-0,82 |
0,20* |
-0,77 |
0,50 |
|
Шкала пастбищной дигрессии |
0,46 |
0,09* |
0,72 |
-0,42 |
0,56 |
-0,20* |
0,61 |
-0,24 |
|
Шкала богатства и засоления почвы |
0,14* |
0,27 |
0,08* |
0,20* |
0,06* |
0,18* |
0,08* |
0,24 |
Примечание: звездочкой помечены недостоверные коэффициенты корреляции
Таблица 5. Средние значения проекций точек геоботанических описаний на оси DCA-ординации (xср) и величины дисперсий этих проекций ( σ 2)
|
Годы |
1961 |
1973 |
2010 |
|||
|
DCA1 |
DCA2 |
DCA1 |
DCA2 |
DCA1 |
DCA2 |
|
|
хср. |
2,33 |
1,19 |
2,70 |
1,08 |
2,52 |
1,25 |
|
0 2 |
0,82 |
0,25 |
0,90 |
0,32 |
0,81 |
0,13 |
Третья группа - влажнолуговое сообщество со слабым влиянием выпаса. Почва, если судить по шкалам Л.Г. Раменского, слабо солончаковатая. Величина phi-коэффициента ни для одного из видов этой группы не достигала 0,25, что свидетельствует о том, что в третью группу вошли виды с широкой экологической амплитудой. Поэтому мы не указываем для этого фитоценона диагностических видов. В данном случае, говоря об экологической амплитуде видов, мы имеем их амплитуду в рамках вы- борки из 158 геоботанических описаний 1973 и 2010 гг. Если такой фитоценон относили бы к какой-либо ассоциации, то в соответствие с существующими взглядами, он бы считался центральным [20, 24, 25]. Сообщество этой группы было 34 раза отмечено в 1973 г. и только 2 - в 2010 г. Значительная часть площадок третьей группы в 2010 г. переместилась в следующую, четвертую группу.
Четвертая группа, также как и предыдущее сообщество, представляет собой влажный луг,
Число кластеров
Рис. 4. Изменение показателя четкости классификации с увеличением числа кластеров
хотя немного более сырой. Влияние выпаса слабое. Диагностические виды: Eleocharis palustris + E. uniglumis, Carex acutiformis + C. melanostachya. Доминирует в большем числе случаев Elymus repens. В 2010 г. сообщество встречалось более чем в два раза чаще в сравнение с 1973 г., что связано с повышенным увлажнением поймы в год последнего учет, проведенного на ключевых участках и трансекте.
Пятая группа - это сырые луга, на которых выражено слабое влияние выпаса. Диагностическим видом является комплекс: Carex acutiformis + C. melanostachya. Он же доминирует в сообществе. Данный фитоценон был представлен одинаковым числом площадок (9) как в 1973 г., так и в 2010 г.
Шестая группа, как и пятая - это сырые луга. Диагностические виды: Butomus umbellatus, Alisma lanceolatum + A. plantago-aquatica, Sagittaria sagittifolia. Доминант Eleocharis palustris + E. uniglumis. Сообщество было представлено только один раз в 1973 г. и на 10 площадках - в 2010 г. Т. е., такие сообщества в 1973 г. были очень редки.
ЗАКЛЮЧЕНИЕ
Сопоставление состава растительного покрова на ключевых участках и трансекте ВАГТ в южной части Волго-Ахтубинской поймы за 1961, 1973, 2010 гг. свидетельствует о том, что наиболее ксерофитным он был в 1973 г. В этом же году фиксировали и наибольшие значения пастбищной дигрессия растительности. Эти явления вызваны малой водностью 1973 г. и предшествующего десятилетия, высокой интенсивностью сельскохозяйственного использования угодий в этот период.
В 2010 г. явных прямых специфических влияний действующей буровой нефтяной уста- новки на растительный покров рассмотренных двух участков и трансекты выявлено не было. Но мониторинг за возможным влиянием этого сооружения на растительность необходимо продолжить. Это не единственное подобная конструкция в южной части Волго-Ахтубинской поймы. Обвалование и постройка дамб, в целом, нарушают естественные условия функционирования пойменных экосистем. По нашим наблюдениям, участки, на которых были сооружены бурильные установки и сопровождающие их инфраструктурные объекты, после прекращения их эксплуатации не подвергаются рекультивации, пойменный режим на них не восстанавливается.
В 2010 г. в южной части поймы, кроме рассмотренных ключевых участков ВАГТ, повторно была обследована трансекта, заложенная в 1953 г. Прикаспийской экспедицией Московского государственного университета [7]. На ней, также как на участках и трансекте ВАГТ, не были выявлены значительные изменения растительного покрова после зарегулирования водного стока.
В среднем и северном отрезках Волго-Ах-тубинской поймы изменения растительности в последние годы учетов более значительны. Там были установлены отчетливо выраженные явления ксерофитизации и рудерализации растительности в 2008-2013 гг. в сравнение с 50-60-ми и, особенно, с 20-ми годами прошлого столетия [2, 3, 6, 8, 9, 11, 15, 18, 19]. Но и они были неповсеместные. Ксерофитизация и рудерализация растительности в этих районах коснулись самых высоких участков поймы, а также мест, в которых произошло углубление русла Волги и заиление вторичных водотоков. Степень рудерализации была обычно выше вблизи транспортных артерий и различного рода сооружений
Везде в долине Нижней Волги значительное
Таблица 6. Группы сообществ, выделенные с помощью кластерного анализа
|
Номер группы |
1 |
2 |
3 |
4 |
5 |
6 |
|
|
Общее количество площадок в группе |
21 |
25 |
36 |
47 |
18 |
11 |
|
|
в 1973 г. |
18 |
4 |
34 |
14 |
9 |
1 |
|
|
в 2010 г. |
3 |
21 |
2 |
33 |
9 |
10 |
|
|
Среднее значение ступени по шкале Л. Г. Раменского |
У |
45 |
63 |
66 |
71 |
79 |
78 |
|
пд |
6 |
5 |
4 |
4 |
4 |
4 |
|
|
БЗ |
15 |
15 |
17 |
16 |
16 |
15 |
|
|
Среднее число видов на площадке |
15 |
15 |
12 |
14 |
12 |
13 |
|
|
Descurainia sophia |
67+ |
4 + |
3 + |
. |
. |
. |
|
|
Alyssum desertorum |
62+ |
8+ |
3+ |
. |
6+ |
. |
|
|
Artemisia austriaca |
711 |
32+ |
. |
9+ |
. |
. |
|
|
Salsola kali |
29+ |
4+ |
. |
. |
. |
. |
|
|
Bromus tectorum |
24+ |
. |
. |
. |
. |
. |
|
|
Eremopyrum triticeum |
291 |
4 + |
. |
2 + |
. |
. |
|
|
Bassia hyssopifolia |
24+ |
4 + |
. |
. |
. |
. |
|
|
Dodartia orientalis |
76 + |
56+ |
39+ |
43+ |
11 + |
. |
|
|
Acroptilon repens |
90+ |
80+ |
421 |
49+ |
28+ |
18+ |
|
|
Glycyrrhiza glabra |
811 |
963 |
111 |
40+ |
. |
9+ |
|
|
Calamagrostis epigejos |
431 |
36+ |
. |
2+ |
. |
. |
|
|
Eleocharis palustris + E. uniglumis |
. |
40+ |
781 |
912 |
781 |
1004 |
|
|
Carex acutiformis + C. melanostachya |
5 1 |
32+ |
531 |
851 |
1003 |
55+ |
|
|
Butomus umbellatus |
. |
. |
. |
13+ |
11 + |
55 + |
|
|
Alisma lanceolatum + A. plantago-aquatica |
. |
4 + |
. |
9 + |
17+ |
45+ |
|
|
Sagittaria sagittifolia |
. |
. |
. |
. |
. |
27+ |
|
|
Carex praecox |
48+ |
40+ |
141 |
171 |
. |
. |
|
|
Cynanchum acutum |
29+ |
24+ |
. |
4+ |
. |
. |
|
|
Artemisia santonicum |
24+ |
16+ |
. |
2 + |
. |
. |
|
|
Artemisia pontica |
29+ |
12+ |
. |
6 + |
11 + |
. |
|
|
Bromus inermis |
38+ |
481 |
. |
30+ |
. |
. |
|
|
Lythrum virgatum |
. |
44 + |
61+ |
64+ |
89+ |
82+ |
|
|
Hierochloe repens |
10+ |
16+ |
531 |
53+ |
781 |
732 |
|
|
Althaea officinalis |
10+ |
32+ |
19+ |
36+ |
56+ |
73+ |
|
|
Rubia tatarica |
. |
8+ |
19+ |
17+ |
39+ |
64+ |
|
|
Asparagus officinalis |
57+ |
52+ |
751 |
79+ |
61+ |
641 |
|
|
Elymus repens |
951 |
962 |
922 |
1003 |
561 |
361 |
|
|
Euphorbia esula s.l. |
62+ |
68+ |
72+ |
74+ |
50+ |
641 |
|
|
Scirpus maritimus s.l. |
. |
24+ |
36+ |
30+ |
33 + |
18+ |
|
|
Inula britannica |
5+ |
44+ |
72+ |
77+ |
61+ |
64+ |
|
|
Convolvulus arvensis |
52+ |
44+ |
31+ |
77+ |
83+ |
64+ |
|
|
Eryngium planum |
38+ |
32+ |
47+ |
15+ |
11+ |
. |
|
|
Senecio jacobaea |
14+ |
40+ |
53+ |
38+ |
11 + |
45+ |
|
|
Xanthium strumarium |
. |
12+ |
14+ |
15+ |
6 + |
36+ |
|
|
Glycyrrhiza echinata |
33 + |
44 + |
58+ |
51+ |
39+ |
36+ |
|
|
Scutellaria galericulata + S. hastifolia |
. |
8 + |
11 + |
30+ |
22 + |
27+ |
|
|
Polygonum arenarium ssp. pulchellum |
52+ |
28+ |
19+ |
23+ |
28+ |
18+ |
|
|
Tragopogon brevirostris ssp. podolicus |
43+ |
36+ |
44+ |
23+ |
22+ |
9+ |
|
|
Euphorbia palustris |
5+ |
16+ |
8+ |
23+ |
39+ |
9+ |
|
Примечания: 1. Встречаемость видов приводится в %. 2. В синоптическую таблицу включены только виды, встречаемость которых превышает 25% в любом из фитоценонов. 3. Значения встречаемости диагностических видов (имеющих значение phi ≥ 0,25) закрашены светло-серым цветом. 4. Надстрочными индексами указаны медианы в ранжированном ряду значимых показателей обилия растений влияние на изменения в растительности оказало уменьшение пастбищной нагрузки и регулярности уборки травы на сено, произошедшие после перехода сельского хозяйства России на рыночные экономические отношения. Эти два антропогенных фактора особенно сильное воздействие оказали на растительность и почвы восточной части дельты р. Волги. Здесь, в отличие от ее западной части, превращенной в орошаемую пашню, сохранился естественный растительный покров. В этом районе дельты следствием уменьшение антропогенного влияния стало увеличение величины надземной массы растений. В результате уменьшилось прямое испарение с поверхности почвы и произошло ее рассоление. В свою очередь, это привело во многих случаях к смене галофитных сообществ гликофитными [10, 11].
Список литературы Оценка динамики растительности южной части Волго-Ахтубинской поймы
- Авакян А.Б., Шарапов В.А. Водохранилища гидроэлектростанций СССР. М.: Энергия, 1977. 400 с.
- Бондарева В.В., Голуб В.Б. Оценка динамики растительности на ключевых участках в средней части Волго-Ахтубинской поймы//Известия Самарского научного центра РАН. 2014. Т. 16. № 5(2) С. 1630-1636.
- Бондарева В.В., Голуб В.Б. Оценка динамики растительности Волго-Ахтубинской поймы (на примере трансекты в районе с. Болхуны)//Поволжский экологический журнал. 2015. № 3. С. 243-252.
- Боровиков В.А. Statistica. Искусство анализа данных на компьютере 2-е изд. СПб.: Питер, 2003. 688 с.
- Глотов К.В., Животовский Л.А., Хованов Н.В., Хромов-Борисов Н.Н. Биометрия. Учебн. пособие. Москва-Ижевск, 2005. 381 с.
- Голуб В.Б., Бармин А.Н., Бондарева В.В., Иолин М.М., Герасимова К.А. Оценка динамики растительности на ключевых участках в северной части Волго-Ахтубинской поймы//Самарская Лука: проблемы региональной и глобальной экологии. 2015. Т. 24. № 4. С. 210-220.
- Голуб В.Б., Бармин А.Н., Иолин М.М., Старичкова К.А., Сорокин А.Н., Шарова И.С., Николайчук Л.Ф. Оценка динамики растительности южной части Волго-Ахтубинской поймы на трансекте в районе села Хошеутово//Известия Самарского научного центра РАН. 2011. Т. 13. № 5. С. 107-113.
- Голуб В.Б., Бондарева В.В., Сорокин А.Н., Бармин А.Н., Иолин М.М, Николайчук Л.Ф. Динамика луговой растительности северной части Волго-Ахтубинской поймы (1928-2009 гг.) 1. Динамика флоры//Вестник Волжского университета им. В.Н. Татищева. Серия «Экология». Тольятти, 2011. Вып. 12. С. 110-120.
- Голуб В.Б., Бондарева В.В., Сорокин А.Н., Бармин А.Н., Иолин М.М., Николайчук Л.Ф. Динамика луговой растительности северной части Волго-Ахтубинской поймы (1928-2009 гг.) 2. Динамика растительных сообществ//Вестник Волжского университета им. В.Н. Татищева. Серия «Экология». Тольятти, 2011. Вып. 12. С. 120-130.
- Голуб В.Б., Старичкова К.А., Бармин А.Н., Иолин М.М., Сорокин А.Н., Николайчук Л.Ф. Оценка динамики растительности в дельте Волги//Аридные экосистемы. 2013. Т. 19. № 3 (56). С. 54-64.
- Голуб В.Б., Бондарева В.В., Шитиков В.К., Бармин А.Н., Иолин М.М. Дополнительные данные о динамике засоления почвы и растительности в дельте р. Волги//Аридные экосистемы. 2015. Т. 21. №3 (64). С. 48-55.
- Голуб В.Б., Добрачев Ю.П., Пастушенко Н.Ф., Яковлева Е.П. О способах оценки экологических условий местообитаний по шкалам Л.Г. Раменского//Биологические науки. 1978. № 7. С. 131-136.
- Голуб В.Б., Сорокин А.Н., Ивахнова Т.Л., Старичкова К.А., Николайчук Л.Ф., Бондарева В.В. Геоботаническая база данных долины Нижней Волги//Известия Самарского научного центра РАН. 2009. Т. 11. № 1 (4). С. 577-582.
- Грин Г.Б. Попуски в нижние бьефы. М.: Энергия, 1971. 95 с.
- Иолин М.М., Сорокин А.Н., Старичкова К.А., Бармин А.Н., Николайчук Л.Ф., Голуб В.Б. Оценка динамики растительности Волго-Ахтубинской поймы на трансекте в районе с. Капустин Яр//Поволжский экологический журнал. 2011. № 4. С. 431-442.
- Кузьмина Ж.В., Трешкин С.Е., Каримова Т.Ю. Динамические изменения наземных экосистем поймы и дельты Нижней Волги под влиянием зарегулирования речного стока и климатических флуктуаций//Аридные экосистемы. 2015. Т. 21. № 4 (65). С. 39-53.
- Раменский Л.Г, Цаценкин И.А., Чижиков О.Н., Антипин Н.А. Экологическая оценка кормовых угодий по растительному покрову. М.: Государственное издательство сельскохозяйственной литературы, 1956. 471 с.
- Сорокин А.Н., Бондарева В.В., Бармин А.Н., Старичкова К.А., Иолин М.М., Николайчук Л.Ф., Голуб В.Б. Динамика флористической структуры сообществ с доминированием Carex acuta в северной части Волго-Ахтубинской поймы//Экология. 2012. № 3. С. 177-183.
- Старичкова К.А., Бармин А.Н., Иолин М.М., Шарова И.С., Сорокин А.Н., Николайчук Л.Ф., Голуб В.Б. Оценка динамики растительности на трансекте в северной части Волго-Ахтубинской поймы//Аридные экосистемы. 2009. T. 15. № 4 (40). C. 36-48.
- Чепинога В.В. Флора и растительность водоемов Байкальской Сибири//Издательство Института географии им. В.Б. Сочавы СО РАН. Иркутск, 2015. 468 с.
- Botta-Dukát Z., Chytrý M., Hájková P., Havlová M. Vegetation of lowland wet meadows along a climatic continentality gradient in Central Europe//Preslia. 2005. Vol. 77. P. 89-111.
- Braun-Blanquet J. Prinzipien einer Systematik der Pflanzengesellschaften auf floristischer Grundlage//Jahr-buches der St. Gallischen Naturwissenschaftlichen Gesellschaft. 1921. Vol. 57 (2) P. 305-351.
- Chytrý M., Tiсhý L., Holt J., Botta-Dukát Z. Determination of diagnostic species with statistical fidelity measures//Journal of Vegetation Science. 2002. Vol. 13. P. 79-90.
- Dengler J., Berg C. & Jansen F. 2005. New ideas for modern phytosociological monographs//Annali di Botanica. Nuova serie. Vol. 5. P. 193-210.
- Dierschke H. 1994. Pflanzensoziologie. Grundlagen und Methoden. Stuttgart: Verlag Eugen Ulmer, 684 S.
- Flora Europaea. Royal Botanic Garden Edinburgh. 2010. Published on the Internet. http://rbg-web2.rbge.org.uk/FE/fe.html, accessed July 2010.
- Tiсhý L. JUICE, software for vegetation classification//Journal of Vegetation Science. 2002. Vol. 13. P. 451-453.
- Tiсhý L., Chytrý M. Statistical determination of diagnostic species for site groups of unequal size//Journal of Vegetation Science. 2006. Vol. 17. P. 809-818.
- Zelený D., Tiсhý L. Linking JUICE and R: New developments in visualization of unconstrained ordination analysis//18th Workshop of European Vegetation Survey in Rome. Roma: La Sapienza Univerzita. 2009. p. 123.
- URL: http://rbg-web2.rbge.org.uk/FE/fe.html (дата обращения 20.09.2016).
- URL: http://www.sci.muni.cz/botany/juice/(дата обращения 20.09.2016).
