Оценка динамики растительности южной части Волго-Ахтубинской поймы на трансекте в районе села Хошеутово

Автор: Голуб В.Б., Бармин А.Н., Иолин М.М., Старичкова К.А., Сорокин А.Н., Шарова И.С., Николайчук Л.Ф.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5-1 т.13, 2011 года.
Бесплатный доступ
Проведен анализ повторных геоботанических учетов (1955, 1971, 1982, 2010 гг.) на трансекте, пересекающей Волго-Ахтубинскую пойму в районе с. Хошеутово (Астраханская область). В 2010 г. отмечена локальная ксерофитизация растительного покрова вблизи ферм и дамб.
Экологические шкалы л.г. раменского, dca-ординация, волго-ахтубинская пойма, индикация
Короткий адрес: https://sciup.org/148205540
IDR: 148205540 | УДК: 633.2.03
Estimate of vegetation dynamics along the transect in the southern part of the Volga-Akhtuba floodplain near Hosheutovo village
Analysis of repeated observations (1955, 1971, 1982, 2010) on the transect crossing the Volga-Akhtuba floodplain near Hosheutovo village in the Astrakhan region was implemented. Local xerophytization of vegetation was found in the vicinity of farms and dams in 2010.
Текст научной статьи Оценка динамики растительности южной части Волго-Ахтубинской поймы на трансекте в районе села Хошеутово
С 1954 по 2010 г. в Волго-Ахтубинской пойме периодически проводили учеты на стационарных трансектах. Итоги наблюдений за динамикой растительности за этот период в северной части поймы были подведены ранее [2, 3, 16, 17]. В настоящей статье осуществлен анализ изменения флоры и растительных сообществ на трансекте, расположенной в южной части поймы.
МАТЕРИАЛ И МЕТОДИКА
В 1955 г. Прикаспийской экспедицией Московского государственного университета (МГУ) в районе с. Хошеутово Астраханской области заложена геоботаническая трансекта (рис. 1). Это было осуществлено до постройки Куйбышевского гидроузла, работа которого в наибольшей степени влияет на гидрологический режим низовий р. Волги [1, 9, 19]. Линия трансекты нанесена на аэрофотоснимки, на которых обведены однородные контуры, пересекаемые ею. Вдоль трансекты, в центральной ее части, проходила грунтовая дорога. Она соединяла c. Хошеутово, расположенное на коренном берегу левобережья долины Нижней Волги, с с. Заволжское, находящимся в восточной части Волго-Ахтубинской поймы на берегу р. Волги. Во время половодий дорога затапливалась и не использовалась. Общая длина трансекты составляла 18 км. Геоботанические описания делали с двух сторон дороги, в нескольких десятках метров от нее.
Сохранились полевые дневники участников Прикаспийской экспедиции МГУ Г.С. Шилова и Л.В. Петровой с описаниями 73 пробных площа-
док, сделанных на трансекте. В дневниках содержится подробная характеристика мест геоботанических описаний на контуре и сами описания, включающие список видов растений с указанием их обилия. После 1955 г. авторы данной статьи посещали трансекту в 1971, 1982 и 2010 гг. Описания осуществляли: в 1955 г. – 08.09-12.09, в 1971 г. – 01.09-03.09, в 1982 г. – 12.07-15.07, в 2010 г. – 08.0816.08. Повторные геоботанические описания проводили примерно в тех же местах, что и в 1955 г. В последний год работы географические координаты пробных площадок на трансекте устанавливали и записывали c помощью GPS-приемника.
В настоящее время полевая дорога, вдоль которой расположена трансекта, используется населением мало, а на некоторых отрезках совершенно заброшена. К 2010 г. два отрезка поймы, пересекаемые трансектой (4,4 км), были обвалованы и переведены в орошаемую пашню. Геоботанические описания, которые были сделаны на обвалованной территории, в обработке материала для данной статьи мы не использовали.
По техническим причинам в 1971 и 1982 гг. не все геоботанические площадки, заложенные в 1955 г., были описаны. Всего при подготовке настоящей статьи мы сопоставили следующее количество описаний, сделанных на трансекте в разные годы: 1955 г. – 53, 1971 г. – 40, 1982 г. – 46, 2010 г. – 53. Следует подчеркнуть, что в 2010 г. были исследованы все сохранившиеся в естественном состоянии участки трансекты. Поэтому сопоставление описаний 1955 и 2010 гг. мы провели более детально, поскольку представилась возможность сравнить одинаковое их количество в эти годы.
Описания, сделанные во все годы исследований, аккумулированы в базе данных EU-RU-002 [7, 20]. Краткая характеристика базы данных EU-RU-002 помещена также в Интернете на сайте В качестве системы управления базой данных использовалась программа TURBOVEG [22].
Названия высших растений приводятся в основном по их списку в базе «Flora Europaea» (2010), помещенной на сайте Эдинбургского королевского ботанического сада: [21]. В тех случаях, когда названия видов не соответствуют этой базе, таксоны приведены с указанием авторов. Низшие растения (мхи и лишайники), зафиксированные в 1982 и 2010 гг., перед обработкой были удалены из описаний, поскольку в 1955 и в 1971 гг. их присутствие не регистрировали.
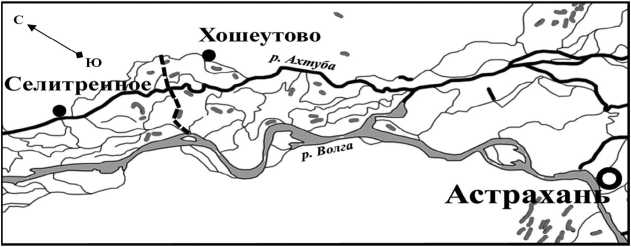
Рис. 1. Схематическая карта Волго-Ахтубинской поймы в районе проведения полевых исследований: пунктиром обозначена геоботаническая трансекта
Для некоторых названий таксонов необходимо сделать пояснения.
Scirpus maritimus s.l. понимается как группа видов: S. maritimus L., S. glaucus Lam. и S. planiculmis F.W. Schmidt. Сведения о распространении последних двух видов в южных районах России появились недавно [11, 18]. Отличаются эти таксоны преимущественно по плодам, которые не всегда бывают представлены. За три последних года работы в северной части поймы (2008-2010 гг.), когда мы стали различать перечисленные выше виды, только однажды вне пробной учетной площадки нами был встречен экземпляр S. planiculmis. Во всех прочих случаях, когда побеги имели плоды, использовавшиеся для определения видов, это всегда был S. glaucus.
Xanthium strumarium s.l. – неотличимые в вегетативном состоянии таксоны Xanthium strumarium ssp. strumarium x X. strumarium ssp. italicum и X. strumarium.
Euphorbia esula s.l. – два недостаточно хорошо различимые в вегетативном состоянии таксона подсекции Esula Boiss.: E. esula ssp. esula и E. esula ssp. tommasiniana.
Polygonum sect. – включает группу не всегда хорошо отличимых в вегетативном состоянии таксонов секции Polygonum и гибридов рода Polygonum: P. arenastrum, P. aviculare, P. neglectum, P. patulum, P. arenarium ssp. pulchellum, P. samarense , P. salsugi-neum, P. arenastrum x P. patulum.
Кроме того, несколько пар видов представлены их суммой, поскольку в вегетативном состоянии их не всегда удавалось различить: Chenopodium album + Ch. acerifolium, Eleocharis palustris + E. uniglumis, Rorippa brachycarpa + R. palustris, Scutellaria galeri-culata + S. hastifolia, Vicia cracca + V. tenuifolia (в подавляющем большинстве случаев встречается первый вид).
Lotus corniculatus gr. – группа неотличимых в вегетативном состоянии видов: L. tenuis , L. ucraini-cus Klokov, L. schoelleri Schweinf., L. stepposus Kramina.
Taraxacum officinale gr. – сборная группа мелких видов, близких к T. officinale.
Medicago sativa ag. = M. sativa ssp. sativa + M. sativa ssp . caerulea.
При анализе динамики флоры мы ограничили список видов вне зависимости от их обилия только теми, встречаемость которых в геоботанических описаниях хотя бы в каком-либо году наблюдений превышала 10%.
К числу доминантов формально были отнесены два вида, отмеченные в геоботанических описаниях с наибольшим обилием. Для сопоставления мы взяли только те из них, которые встретились чаще, чем в 10% описаний хотя бы в одном из годов наших учетов на трансекте.
Для расчета экологических ступеней по шкалам Л. Г. Раменского (увлажнения, богатства и засоленности почвы, пастбищной дигрессии) применяли «метод пересечения большинства интервалов» [14]. Последний основан на определении моды в статистическом ряду распределения чисел, входящих в интервалы ограничительных ступеней экологических шкал растений [6]. Поэтому индикация условий среды не очень сильно зависит от длины флористического списка при условии, что геоботаники описывают однородную в экологическом отношении площадку.
Сравнение распределения совокупности описаний по ступеням показателей шкал Л.Г. Раменского, рассчитанных для каждого года наблюдений, вначале проводили с помощью теста Краскела-Уоллиса. Решался вопрос: относятся ли сопоставляемые группы к одной или к разным генеральным совокупностям? Когда нулевая гипотеза не подтверждалась (т.е., сопоставляемые выборки относились к разным генеральным совокупностям), сравнение выборок производили попарно с использованием теста Манна-Уитни. Для оценки определения смещения средних значений использовали критерий Стьюдента [4, 5]. При всех статистических оценках величины считали достоверными, если p-значение соответствующей статистики не превышало уровень значимости 0.05.
Состав растительных сообществ устанавливали с помощью программы TWINSPAN в среде JUICE [23, 24]. При этом обрабатывали общую совокупность описаний за 1955 и 2010 гг. (106 описаний). В синоптическую табл. 2 включены только виды, встречаемость которых превышает 50% в каком-либо из фи-тоценонов.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Флористический состав. Количество видов и их агрегаций, встречаемость которых хотя бы в один
Таблица 1 . Встречаемость видов растений и их агрегаций, %
Что касается состава доминантов растительных сообществ на территории, пересекаемой трансектой, то в их число входят Carex melanostachya, C. praecox, Elymus repens, Eleocharis palustris + E. uniglumis, E. acicularis, Hierochloë repens, Agrostis stolonifera, Bromus inermis, Butomus umbellatus, Asparagus officinalis и Glycyrrhiza glabra . Среди этих видов некоторую тенденцию к направленному уменьшению встречаемости можно обнаружить у Butomus umbellatus (в %): 1955 г. – 11, 1971 г. – 5, 1982 г. – 4, 2010 г. – 4, а также у Asparagus officinalis : 1955 г. – 11, 1971 г. – 8, 1982 г. – 2, 2010 г. – 2. У остальных видов этот показатель флуктуировал.
Сообщества. При анализе синоптической табл. 2 мы не будем обсуждать сообщества, которые составлены только из одного описания. Предположим, что это случайные комбинации видов. В этой таблице такой группой является 4-я.
Характеризуя в целом результаты обработки программой TWINSPAN общей совокупности описаний за 1955 и 2010 гг., можно отметить, что группы описаний оказались в основном расположены в табл. 2 вдоль двух связанных между собой градиентов: увлажнения и пастбищной дигрессии. Слева от главного разделителя расположены более сырые местообитания с меньшей пастбищной дигрессией, справа – более сухие, с большей пастбищной дигрессией.
Первая и вторая группы – это растительные группировки с доминированием Potamogeton lucens. Эти фитоценозы были представлены только в 2010 г. во временных, пересыхающих к осени водоемах центральной поймы.
Таблица 2 . Группы растительных сообществ, выделенные с помощью программы TWINSPAN
Примечания: У – увлажнение, БЗ – богатство и засоленность почвы, ПД – пастбищная дигрессия; вертикальными линиями указаны разделители 1-3 порядков в соответствии с алгоритмом TWINSPAN; встречаемость видов приводится в %; надстрочные числа – значения медианы обилия растений по шкале В.Б. Голуба [13] в ряду описаний, в которых данный вид встречается; полужирным шрифтом выделены числа в ячейках, в которых значения медианы обилия превышают единицу.
В 1955 г. местообитания этих групп занимали менее тесно связанные с водой сообщества 3-й группы. Поскольку в 1955 г. описания на трансекте производились на месяц позже, чем в 2010 г., то к этому времени многие водоемы уже значительно высохли. Надземные побеги P. lucens, легко и бы- стро разрушаемые при высыхании, могли быть уже незаметны. Вероятно, подобное могло произойти и с побегами Nymphoides peltata, отмеченного в 2010 г., но не упомянутым в описаниях 1955 г. Поэтому бóльшую представленность гигрофитных сообществ в 2010 г. в сравнении с 1955 г. можно объяс- нить более ранним сроком проведения полевых исследований.
Небольшое перераспределение представленности местообитаний разной степени увлажнения произошло между 5-й и 6-й группами. Встречаемость сообществ последней более ксерофитной группы с большей выраженностью пастбищной дигрессии немного увеличилась в 2010 г. Произошло это за счет сокращения количества описаний, индицирующих более мезофитные и менее подверженные выпасу экотопы (местообитания 5-й группы). Анализ показал, что «переход» описаний из 5-й в 6-ю группу произошел на экотопах, прилегающих снаружи к обвалованным участкам. Очевидно, обвалование приводило к изменению гидрологического и гидрогеологического режима не только внутри площади, огороженной дамбой, но и на прилегающей территории с наружной ее стороны.
Седьмая группа представлена одинаковым числом описаний в 1955 и 2010 гг. Восьмая, наиболее ксерофитная группа, с большой представленностью пастбищных видов растений, состоит из двух описаний 2010 г. Она объединяет фитоценозы, сформировавшиеся на высоких незатапливаемых во время половодий гривах вблизи животноводческой фермы. Наличие этой группы, также как и увеличение представленности 6-й группы, может свидетельствовать о процессах ксерофитизации, происходящих в растительном покрове.
Индикация условий среды с использованием шкал Л.Г. Раменского. Статическая обработка не выявила каких-либо достоверных изменений распределения описаний по ступеням шкал Л.Г. Раменского за разные годы. Причиной этого может быть не только действительное отсутствие изменений, но и небольшое число сопоставленных описаний.
Таблица 3 . Коэффициенты корреляции между показателями шкал Л.Г. Раменского
В то же время было установлено наличие корре- I ляции между рядами показателей увлажнения и ] рядами ступеней пастбищной дигрессии (табл. 3), что уже проявилось и при рассмотрении сообществ (см. табл. 2). Интересно, что ступени шкал увлаж- *
нения и пастбищной дигрессии имели между собой разную тесноту связи в 1955 и в 2010 гг. В первом
|
Шкалы |
Богатства и засоленности почвы |
Пастбищной дигрессии |
||
|
1955 г. |
2010г. |
1955 г. |
2010 г. |
|
|
Увлажнения |
-0.09* |
-0.25* |
-0.46 |
-0.91 |
|
Богатства и засоленности почвы |
х |
х |
-0,29* |
0,24* |
Примечание: недостоверные значения корреляции отмечены звездочкой.
случае коэффициент корреляции был равен (-0,46), во втором – (-0,91). И в том, и в другом случае с уменьшением увлажнения пастбищная дигрессия нарастала. Это вполне естественно. Чем выше эко-топы находятся над меженью реки, тем на меньший срок они затапливаются и могут больше времени использоваться под выпас. Причиной большей величины коэффициента корреляции между ступенями увлажнения и пастбищной дигрессии в 2010 г., по сравнению с 1955 г., мог явиться более растянутый диапазон варьирования местообитаний сопоставляемых шкал в последний год учетов.
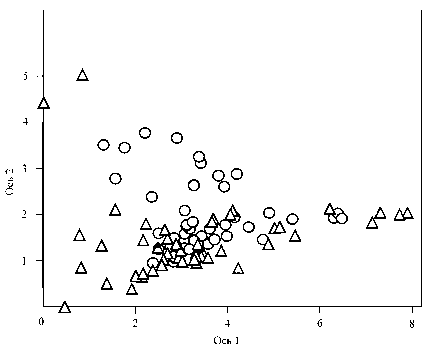
Рис. 2. Диаграмма DCA-ординации описаний 1955 г. ( CD ) и 2010 г. ( ) в поле 1-й и 2-й ординацион-ных осей. Собственные значения осей, характеризующие долю общей информации: для оси 1 = 0,59, для оси 2 = 0,28; длина градиента вдоль оси 1 = 8,63, оси 2 = 3,75.
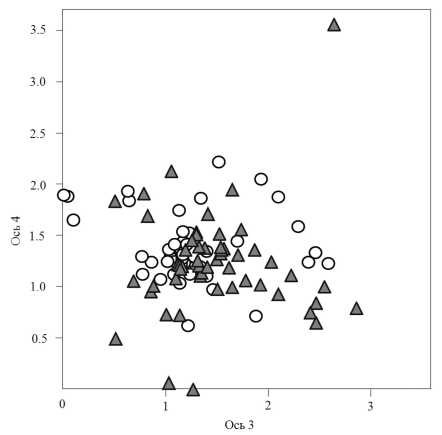
Рис. 3. Диаграмма DCA-ординации описаний 1955 г. ( CD ) и 2010 г. ( ) в поле 3-й и 4-й ординацион-ных осей. Собственные значения осей, характеризующие долю общей информации: для оси 3 = 0,22, для оси 4 = 0, 22; длина градиента вдоль оси 3 = 3,53, оси 4 = 3.47.
Таблица 4 . Коэффициенты корреляции между координатами учетных площадок 1955 и 2010 гг. в поле осей DCA-ординации и показателями шкал Л. Г. Раменского
|
Ось DCA-ординации |
Ось 1 |
Ось 2 |
Ось 3 |
Ось 4 |
||||
|
Год |
1955 |
2010 |
1955 |
2010 |
1955 |
2010 |
1955 |
2010 |
|
Шкала: |
||||||||
|
увлажнения |
0,79 |
0,86 |
-0,38 |
|||||
|
богатства и засоленности |
-0,41 |
-0,39 |
-0,39 |
|||||
|
пастбищной дигрессии |
-0,65 |
-0,89 |
0,55 |
0,30 |
0,46 |
|||
Примечание: недостоверные значения корреляции в таблице не приведены.
DCA-ординация. Достоверные значения корреляции показателей шкал Л.Г. Раменского были обнаружены с четырьмя осями DCA-ординации (табл. 4). Наиболее информативна первая ось, координаты которой имеют сильную корреляцию со ступенями шкал увлажнения и пастбищной дигрессии. Поэтому ось 1 можно интерпретировать как ось увлажнения – пастбищной дигрессии. Визуально хорошо видна бóльшая дисперсия описаний в поле 1-й и 2-й осей DCA-ординации в 2010 г., чем в 1955 г. (рис. 2). Это соответствует большему интервалу разброса значений ступеней увлажнения и пастбищной дигрессии шкал Л.Г. Раменского в 2010 г. в сравнении с 1955 г. В меньшей степени различие в дисперсиях видно в поле 3-й и 4-й осей DCA-ординации (рис. 3).
Оценка смещений координат «облаков» описаний относительно осей DCA-ординации за 1955 и 2010 гг. показала, что по критерию Манна-Уитни оно было достоверно вдоль 2-ой, 3-ей и 4-ой оси, а по критерию Стьюдента смещение средних значений координат было существенно только вдоль 2ой и 3-ей оси. Это оси, которые по шкалам Л.Г. Раменского не столь хорошо интерпретируются, как 1-я. Можно полагать, что изменения во флористическом составе сообществ могут быть и не связаны с изменениями условий увлажнения, богатства и засоленнноси почвы и влиянием выпаса сельскохозяйственных животных.
ВЫВОДЫ
Существует некоторая противоречивость полученных результатов. С одной стороны, мы отмечаем уменьшение встречаемости на трансекте в 2010 г., по сравнению с 1955 г., рудеральных и пастбищных видов растений, а с другой – увеличение представленности сообществ с более выраженной пастбищной дигрессией и большей ксерофитизаци-ей растительного покрова. Это противоречие мы объясняем тем, что в целом в районе трансекты пастбищная нагрузка к 2010 г. уменьшилась. Однако локально, вблизи ферм и валов, ограждающих пашню, она увеличилась. На обвалованных территориях скот может находиться и во время половодий. Кроме того, создание дамб нарушило гидрологический и гидрогеологический режим и за пределами собственно обвалованной территории.
Изменения в растительном покрове на трансекте в южной части Волго-Ахтубинской поймы вблизи с. Хошеутово менее выражены, чем те, которые были зафиксированы на трансектах в северной части поймы, у г. Ленинска и с. Капустин Яр [10, 17]. Это может быть связано с тем, что в северной части, в отличие от южной, в последние десятилетия, произошло углубление русла р. Волги, что привело к ухудшению обводнения поймы в этом районе [8, 12, 15].
Работа выполнена при поддержке Российского фонда фундаментальных исследований (грант 0905-00183).
Список литературы Оценка динамики растительности южной части Волго-Ахтубинской поймы на трансекте в районе села Хошеутово
- Авакян А.Б., Шарапов В.А. Водохранилища гидроэлектростанций СССР. М.: Энергия. 1977. 400 с.
- Бармин А.Н., Голуб В.Б., Иолин М.М., Асанова Г.З. 2010а. Индикация изменений условий среды в северной части Волго-Ахтубинской поймы при использовании шкал Л.Г. Раменского и DCA-ординации//Изв. высш. учеб. заведений. Раздел геодезия и аэрофотосъемка. 2010. № 5. С. 21-24.
- Бармин А.Н., Иолин М.М., Шарова И.С. и др. Использование шкал Л.Г. Раменского и DCA-ординации для индикации изменений условий среды в Волго-Ахтубинской пойме//Изв. Самар. НЦ РАН. 2010. Т. 12, № 1. С. 54-57.
- Боровиков В.А. Statistica. Искусство анализа данных на компьютере. 2-е изд. СПб.: Питер, 2003. 688 с.
- Глотов Н.В, Животовский Л.А., Хованов Н.В., Хромов-Борисов Н.Н. Биометрия. Учебное пособие. М.; Ижевск. 2005. 381 с.
- Голуб В.Б., Добрачев Ю.П., Пастушенко Н.Ф., Яковлева Е.П. О способах оценки экологических условий местообитаний по шкалам Л.Г. Раменского//Науч. докл. высш. шк. Биол. науки. 1978. № 7. С. 131-136.
- Голуб В.Б., Сорокин А.Н., Ивахнова Т.Л. и др. Геоботаническая база данных долины Нижней Волги//Изв. Самар. НЦ РАН. 2009. Т. 11, № 1(4). C. 577-582.
- Горелиц О.В., Землянов И.В., Синенко Л.Г. Оценка морфометрических характеристик русла при планировании мероприятий по водообеспечению территорий Нижней Волги//Сб. докл. Междунар. конф. «Управление водно-ресурсными системами в экстремальных условиях». М., 2008. С. 306-307.
- Грин Г.Б. Попуски в нижние бьефы. М.: Энергия. 1971. 95 с.
- Иолин М.М., Сорокин А.Н., Старичкова К.А. и др. Оценка динамики растительности Волго-Ахтубинской поймы на трансекте в районе с. Капустин Яр//Поволжск. экол. журн. 2011. (в печати).
- Клинкова Г.Ю. Род 6 (468). Bolboschoenus (Aschers.) Palla -Клубнекамыш//Флора Нижнего Поволжья. Т. 1. М.: Т-во науч. изд. КМК, 2006. С. 264-267.
- Коротаев В.Н., Бабич Д.Б., Чалов Р.С. (Ред.). Атлас русловой морфодинамики Нижней Волги. М.: Изд-во МГУ, 2009. 232 с.
- Нешатаев Ю.Н. О некоторых задачах и методах классификации растительности//Растительность России. 2001. № 1. С. 17-35.
- Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипин Н.А. Экологическая оценка кормовых угодий по растительному покрову. М.: Сельхозгиз, 1956. 471 с.
- Рычагов Г.И., Коротаев В.Н. (Ред.). Нижняя Волга: геоморфология, палеогеография и русловая морфодинамика. М.: ГЕОС, 2002. 242 с.
- Сорокин А.Н., Бармин А.Н., Иолин М.М. и др. Опыт использования шкал Л.Г. Раменского и DCA-ординации для индикации изменений условий среды на трансекте в районе с. Капустин Яр//Вестник Волжского университета им. В.Н. Татищева. Серия «Экология». Тольятти, 2010. Вып. 10. С. 74-80.
- Старичкова К.А., Бармин А.Н., Иолин М.М. и др. Оценка динамики растительности на трансекте в северной части Волго-Ахтубинской поймы//Аридные экосистемы. 2009. T. 15, № 4 (40). C. 36-48.
- Татанов И.В. Таксономический обзор рода Bolboschoenus (Aschers.) Palla (Cyperaceae)//Новости систематики высших растений. Т. 39. М.; СПб.: Т-во науч. изд. КМК, 2007. С. 46-149.
- Цаценкин И.А. Растительность и естественные кормовые ресурсы Волго-Ахтубинской поймы и дельты р. Волги//Природа и сельское хозяйство Волго-Ахтубинской долины и дельты р. Волги. М.: Изд-во МГУ, 1962. С. 118-192.
- Dengler J., Jansen F., Glöckler F. et al. The Global Index of Vegetation-Plot Databases (GIVD): a new resource for vegetation science//J. Veg. Sci. 2011. V. 22. (In press).
- Flora Europaea. Royal Botanic Garden Edinburgh. Published on the Internet. http://rbgweb2. rbge.org.uk/FE/fe.html, accessed July 2010.
- Hennekens S.M., Schaminée J.H.J. TURBOVEG, a comprehensive data base management system for vegetation data//J. Veg. Sci. 2001. V. 12. P. 589-591.
- Hill M.O. TWINSPAN -a FORTRAN program for arranging multivariate data in an ordered two way table by classification of the individuals and the attributes. Ithaca, NY, 1979. 48 p.
- Tiсhý L. JUICE, software for vegetation classification//J. Veg. Sci. 2002. V. 13. P. 451-453.
