Оценка динамики растительности на ключевых участках в северной части Волго-Ахтубинской поймы

Автор: Бармин А.Н., Иолин М.М., Бондарева В.В., Герасимова К.А., Голуб В.Б.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 4 т.24, 2015 года.
Бесплатный доступ
В статье изложены результаты наблюдений в период 1973-2009 гг. за динамикой флоры и растительных сообществ на ключевых участках в северной части Волго-Ахтубинской поймы. В 2009 г. выявлены изменения флоры и растительности, вызванные снижением пастбищной нагрузки и нарушением регулярности сенокошения.
Волго-ахтубинская пойма, динамика растительности, экологические шкалы л. г. раменского, dca-ординация
Короткий адрес: https://sciup.org/148314978
IDR: 148314978 | УДК: 633.2.03
Текст краткого сообщения Оценка динамики растительности на ключевых участках в северной части Волго-Ахтубинской поймы
Волго-Ахтубинская пойма и дельта р. Волги – уникальный регион с естественной пойменной растительностью. Существование здесь, в зоне пустыни, лугов, болот и лесов обязано ежегодным весенне-летним половодьям, которые с 1959 г. регулируются каскадом волжских водохранилищ. Искусственные попуски заменили естественные половодья, которые отличались большей продолжительностью и более высоким подъемом воды. Характер современного растительного покрова долины Нижней Волги определяется также ее хозяйственным использованием. До половодья обычно повсеместно пасут скот, после половодья и отрастания травы эти же угодья используют как сенокосы. Затем луга вновь используют как пастбища. Из-за более раннего окончания половодья в условиях зарегулированного водного стока убирать сено на лугах начинают на 1-1.5 месяца раньше, а в связи с механизацией темпы сеноуборочной компании выше, чем, например, в середине прошлого века. Соответственно раньше начинают и выпас сельскохозяйственных животных по стерне и отаве. По этой причине после зарегулирования водного стока возможность пастбищного использования поймы возросла.
Данная статья продолжает серию публикаций, посвященных мониторингу растительного покрова долины Нижней Волги под воздействием антропогенных и
природных факторов (Голуб и др., 2009; Старичкова и др., 2009; Иолин и др., 2011; Сорокин и др., 2012; Голуб и др., 2013, 2015; Бондарева, Голуб, 2014).
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Методика исследований и обработка материала во многом были такие же, как и при оценке динамики растительности на ключевых участках средней части Волго-Ахтубинской поймы (Бондарева, Голуб, 2014). Поэтому в этом разделе мы обратим внимание лишь на те аспекты нашей работы, которые отличались от изложенных ранее.
В 1959 г. геоботанической партией Всесоюзного аэрогеологического треста в Волго-Ахтубинской пойме в северной части Волго-Ахтубинской поймы были заложены 4 стационарных участка: у ерика Семеновский (28 га), у озера Бабечье (52 га), у хутора Стасов (42 га), у хутора Никонов (35 га) (рис. 1). Первый участок расположен в Ленинском районе Волгоградской области, три остальных – в Ахтубинском районе Астраханской области. Все 4 участка по своей геоморфологии находятся в центральной части поймы вдали от больших дорог и населенных пунктов.
Для каждого участка были составлены топографические карты в масштабе 1:2000. На участках были проведены почвенные и геоботанические изыскания. Площадки, на которых в 1959 г. были сделаны почвенные разрезы и геоботанические описания, были нанесены на карты. Эти описания сохранились. После 1959 г. участки посещали в 1973, 1981 и 2009 гг. Ориентируясь по картам, повторные геоботанические описания проводили примерно в тех же местах, что и в 1959 г. Описания, сделанные во все годы исследований, аккумулированы на основе программы TURBOVEG в базе данных Lower Volga Phytosociological Database, зарегистрированной в международной системе «Global Index of Vegetation-Plot Databases» с индексом EU-RU-002 (Голуб и др., 2009; Golub et al., 2012).
Всего при подготовке настоящей статьи мы сопоставляли описания, сделанные на 75 учетных площадках. Следует заметить, что геоботанические описания 1959 г. были неполные, поэтому, оценивая изменения флоры и состав растительных сообществ, мы сравнивали данные геоботанических описаний только последних трех лет учетов.
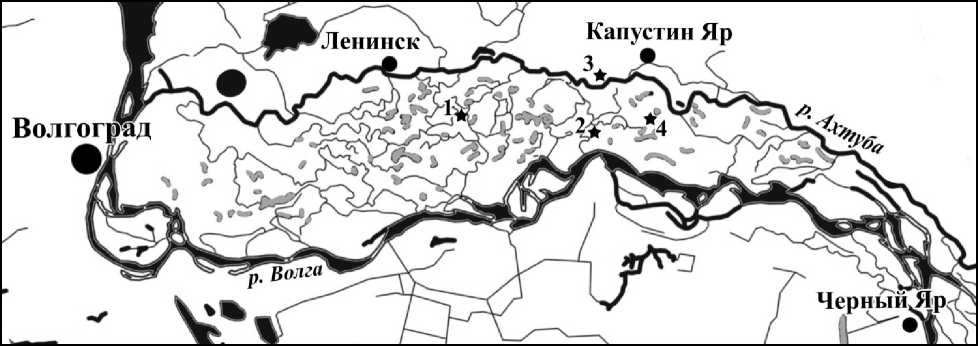
Рис. 1. Схематическая карта района проведения полевых исследований:
звездочками показано расположение ключевых участков: 1 – у ерика Семеновский (Волгоградская обл.), 2 – у озера Бабечье, 3 – у хутора Стасов, 4 – у хутора Никонов
Геоботанические описания в 1973 г. были сделаны в период 07.08 – 17.08, в 1981 г. – 13.08 – 17.08, в 2009 г. – 08.08 – 15.08.
Названия высших растений даем по их списку в базе «Flora Europaea», помещенной на сайте Эдинбургского королевского ботанического сада . Для некоторых названий таксонов необходимо сделать пояснения.
Xanthium strumarium s.l. – неотличимые в вегетативном состоянии таксоны Xanthium strumarium ssp. strumarium xX. strumarium ssp. italicum иX. strumarium.
Euphorbia esula s.l. – два недостаточно хорошо различимые в вегетативном состоянии таксона подсекции Esula Boiss.: E. esula ssp. esula и E. esula ssp. tommasiniana .
Polygonum sect. – включает группу не всегда хорошо отличимых в вегетативном состоянии видов рода Polygonum и их гибридов : P. arenastrum , P. aviculare , P. bellardii , P. neglectum , P. patulum , P. salsugineum , P. arenarium.
Carex agr. – агрегация плохо различимых в вегетативном состоянии таксонов, которая включает C. melanostachya , C. acutiformis , C. riparia . По нашему мнению, в этой агрегации преобладает первый из перечисленных видов.
Plantago major s.l. – неотличимые в вегетативном состоянии P. major ssp. major и P. major ssp. intermedia .
Lotus corniculatus s.l. – плохо различимые в вегетативном состоянии таксоны: Lotus corniculatus var. schoelleri , L. stepposus, L. tenuis .
Scirpus maritimus s.l. = Bolboschoenus glaucus + Scirpus maritimus ssp. maritimus.
Taraxacum officinale group – сборная группа мелких видов, близких к T. officinale.
Кроме того, несколько пар видов представлены их суммой, поскольку в вегетативном состоянии их не всегда удавалось различить: Eleocharis palustris + E. uniglumis , Alisma plantago-aquatica + A. laceolatum (явно преобладает последний), Scutellaria galericulata + S. hastifolia , Scirpus lacustris + S. hippolyti.
При анализе динамики флоры мы ограничили список видов только теми, встречаемость которых в геоботанических описаниях хотя бы в каком-либо году наблюдений равнялась или превышала 15%. К числу доминантов формально были отнесены два вида, отмеченные в геоботанических описаниях с наибольшим обилием. Для сопоставления мы взяли только те из них, которые встретились чаще, чем в 10% описаний хотя бы в одном из годов наших учетов.
Cостав растительных сообществ устанавливали и сравнивали с помощью программы TWINSPAN в среде JUICE (Hill, 1979; Tichý, 2002). При этом обрабатывали общую совокупность описаний за три года: 1973, 1981 и 2009 (225 описаний). В синоптическую табл. 6 включены только виды, встречаемость которых равна или превышает 50% в любой из групп.
При всех статистических оценках величины считали достоверными, если p-значение соответствующей статистики не превышало уровень значимости 0.05.
Данные о метеорологических и гидрологических факторах получены в органах гидрометеослужбы. За объем половодий мы условно принимаем сток воды в створе Волгоградской ГЭС в течение второго квартала, в период которого проводятся специальные попуски воды в нижний бьеф гидроузла (Грин, 1971).
Косвенно о пастбищной нагрузке мы судим по количеству поголовья скота в зоне долины Нижней Волги. Это вся Астраханская область и три южных района Волгоградской области (Ленинский, Светлоярский и Среднеахтубинский). Сведения о поголовье скота получены в органах статистики Астраханской и Волгоградской областей.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЯ
Факторы среды. Следует отметить, что за период наблюдений отмечается нарастание увлажненности территории исследований за счет осадков, что характерно для всей Прикаспийской низменности (Титкова, 2003). Но увлажнение северной части Волго-Ахтубинской поймы уменьшилось после зарегулирования водного стока Волги за счет сокращения объемов половодий. В годы учетов самым маловодным был 1973 г. (табл. 1 и 2).
Если судить по поголовью скота, то минимальная пастбищная нагрузка была в 20002009 гг. Наши личные наблюдения свидетельствуют также о том, что и сенокосное использование лугов в этот период значительно снизилось. Мы многократно видели массивы лугов, оставшиеся осенью нескошенными, особенно в глубине поймы вдали от населенных пунктов и дорог. В предыдущие периоды весь травостой в пойме тщательно выкашивался на всей территории.
Таблица 1
Среднемноголетние показатели экологических факторов
|
Годы |
Средняя сумма осадков, мм |
со „ О св О 1-| н О св и а rt о и р |
и со о ° ю м ^ a 2 1 |
1S SO^ И И « о О о Й щ У И S s s я о S о н -&и S И и |
Объем водного стока в створе Волгоградской ГЭС, км 3 |
Й <0 Я и со О )у § § g s S g g р DS s о и « ё у й О щ й 5 Cd о S § 5 5 у S о о р У ^ « 6 и rt о ° о со |
У о ° S Ь и н о ° к В СО Е _ со >5 с? н св ° & о н m м и о СО О о у Ч g со S х g g s @ О Д н И р. ° й ” й И о S к ц G ч о и |
||
|
за год |
за период с t ° C более 10ºC |
||||||||
|
за год |
за второй квартал |
||||||||
|
1964-1973 |
223 |
110 |
7.2 |
3415 |
0.32 |
225 |
100 |
751 |
560 |
|
1972-1981 |
283 |
137 |
8.8 |
3443 |
0.40 |
232 |
93 |
759 |
571 |
|
2000-2009 |
285 |
157 |
10.0 |
3608 |
0.43 |
256 |
106 |
770 |
346 |
Температуры и осадки приводятся по данным гидрометеостанции в с. Черный Яр
Таблица 2
Метеорологические и гидрологические показатели в годы учетов на ключевых участках
|
Год |
Объем водного стока в створе Волгоградской ГЭС, км3 |
Максимальный уровень подъема воды по рейке водомерного поста в с. Черный Яр |
Сумма осадков за I VII месяцы (гидрометеостанция Черный Яр), мм |
|
|
за год |
за второй квартал |
|||
|
1973 |
174 |
78 |
726 |
170 |
|
1981 |
293 |
128 |
796 |
167 |
|
2009 |
238 |
93 |
746 |
161 |
Флористический состав. Количество видов и их агрегаций, встречаемость которых хотя бы в одном из лет учетов достигала 15%, составило 53 (табл. 3). Анализируя эту таблицу, можно сделать следующие заключения.
-
1. Ярко выделяется повышенная встречаемость растений, характерных для пастбищного использования угодий в 1981 г. Это Artemisia abrotanum , Plantago major s.l., Taraxacum officinale group, Bidens frondosa + B. tripartita , Setaria viridis.
-
2. Можно отметить высокую представленность грубых высокорослых растений в 2009 г. из группы разнотравья, отрицательно реагирующих на сенокошение: Lythrum virgatum , Carex agr., Cirsium arvense , Stachys palustris , Alisma plantago-aquatica + A. lanceolatum , Artemisia pontica , Phragmites australis , Thalictrum flavum , Lycopus exaltatus.
-
3. Одновременно в 2009 г. уменьшилась встречаемость злаков, характерных для сенокосных угодий: Bromus inermis , Elymus repens , Hierochloë repens , Phalaris arundinacea .
Что касается состава доминантов растительных сообществ, то это Bromus inermis , Carex agr., Asparagus officinalis , Eleocharis palustris + E. uniglumis , Elymus repens , Cirsium arvense , Carex praecox , Inula britannica . У всех этих видов представленность в сообществах растений значительно флуктуировала. Тенденцию к направленному увеличению встречаемости имеет агрегации видов Alisma plantago-aquatica + A. lanceolatum и Eleocharis palustris + E. uniglumis. Среди доминантов в 2009 г. резко уменьшилась встречаемость Bromus inermis одновременно с ростом представленности Cirsium arvense и Carex agr.
Таблица 3Встречаемость видов растений и их агрегаций, %
|
Год учета |
1973 |
1981 |
2009 |
|
Bromus inermis |
8843 |
9169 |
4315 |
|
Eleocharis palustris + E. uniglumis |
8521 |
88 32 |
8344 |
|
Rorippa palustris + R. brachycarpa |
75 |
21 |
16 |
|
Carex agr. |
4911 |
518 |
7123 |
|
Lythrum virgatum |
61 |
76 |
76 |
|
Inula britannica |
7911 |
87 |
795 |
|
Senecio jacobaea |
79 |
68 |
21 |
|
Asparagus officinalis |
568 |
5312 |
573 |
|
Hierochloë repens |
52 |
48 |
37 |
|
Elymus repens |
5916 |
558 |
318 |
|
Galium verum |
53 |
61 |
44 |
|
Euphorbia esula s.l. |
60 |
89 |
80 |
|
Tragopogon brevirostris ssp. podolicus |
67 |
48 |
21 |
|
Allium angulosum |
56 |
31 |
235 |
|
Lotus corniculatus s.l. |
48 |
51 |
29 |
|
Carex praecox |
4317 |
4715 |
3711 |
|
Convolvulus arvensis |
41 |
52 |
61 |
|
Cirsium arvense |
394 |
41 |
6919 |
|
Rumex thyrsiflorus |
37 |
39 |
16 |
|
Gratiola officinalis |
33 |
49 |
44 |
|
Polygonum sect. |
31 |
9 |
16 |
|
Scutellaria galericulata + S. hastifolia |
29 |
36 |
33 |
|
Achillea cartilaginea |
25 |
24 |
24 |
|
Xanthium strumarium s.l. |
25 |
59 |
57 |
|
Eryngium planum |
24 |
39 |
29 |
|
Galium rubioides |
21 |
37 |
35 |
|
Rubia tatarica |
20 |
24 |
21 |
|
Vicia cracca |
20 |
27 |
31 |
|
Stachys palustris |
20 |
27 |
36 |
|
Scirpus maritimus s.l. |
19 |
28 |
20 |
|
Euphorbia palustris |
15 |
27 |
16 |
|
Scirpus lacustris + S. hippolyti |
15 |
21 |
12 |
|
Potentilla bifurca |
12 |
16 |
12 |
|
Alisma plantago-aquatica + A. lanceolatum |
12 |
16 |
20 |
|
Mentha arvensis |
11 |
13 |
17 |
|
Rumex hydrolapathum |
20 |
27 |
. |
|
Phalaris arundinacea |
16 |
16 |
9 |
|
Bidens frondosa + B. tripartita |
13 |
26 |
16 |
|
Butomus umbellatus |
12 |
33 |
7 |
|
Eleocharis acicularis |
29 |
1 |
. |
|
Althaea officinalis |
9 |
20 |
19 |
|
Artemisia pontica |
9 |
13 |
27 |
|
Plantago major s.l. |
9 |
40 |
3 |
|
Taraxacum officinale group |
9 |
21 |
3 |
|
Sonchus arvensis |
4 |
16 |
7 |
|
Rumex stenophyllus |
3 |
17 |
. |
|
Setaria viridis |
9 |
15 |
8 |
|
Artemisia abrotanum |
7 |
15 |
8 |
|
Phragmites australis |
9 |
9 |
13 |
|
Chenopodium polyspermum |
8 |
9 |
20 |
|
Thalictrum flavum |
8 |
9 |
19 |
|
Beckmannia eruciformis |
1 |
5 |
15 |
|
Lycopus exaltatus |
. |
. |
16 |
Надстрочные индексы – число встреч вида в качестве доминанта.
DCA-ординация. Абсциссу диаграммы ординации (DCA 1) можно интерпретировать как комплексную ось, отражающую одновременно степень увлажнения и пастбищной дигрессии (рис. 2). Ее координаты хорошо коррелируют с соответствующими показателями шкал Л.Г. Раменского. Имеется отрицательная зависимость со шкалой увлажнения и положительная - с показателями пастбищной дигрессии. Что касается второй оси DCA-ординации, то она интерпретируется гораздо хуже, отражая слабое влияние на растительность увлажнения, ее переменности и богатства почвы (табл. 4).
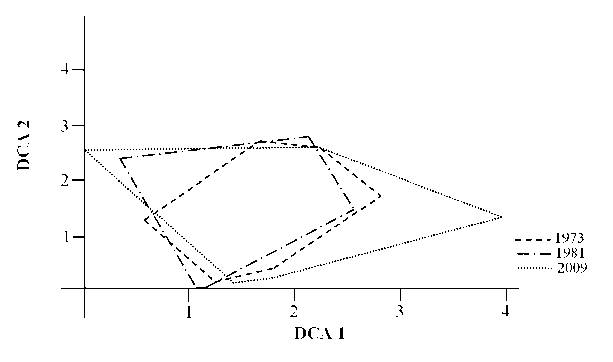
Рис. 2. Диаграмма DCA-ординации геоботанических описаний учетных площадок: линии обводят внешние границы «облаков» описаний, сделанных в разные годы. Собственные значения осей, характеризующее долю общей информации: ось 1 = 0.55, ось 2 = 0.29
Таблица 4
Коэффициенты корреляции значений координат геоботанических описаний на оси DCA-ординации с показателями шкал Л. Г. Раменского
Год I 1973 1981 2009 Все годы
1-я ось DCA-ординации
|
Шкала увлажнения |
-0.75 |
-0.77 |
-0.61 |
-0.70 |
|
Шкала пастбищной дигрессии |
0.52 |
0.52 |
0.60 |
0.55 |
|
Шкала богатства почвы |
0.06* |
0.25 |
-0.20* |
0.01* |
|
Шкала переменности увлажнения |
-0.02* |
0.12* |
-0.19* |
-0.03* |
2-я ось DCA-ординации
|
Шкала увлажнения |
-0.37 |
-0.37 |
-0.19* |
-0.32 |
|
Шкала богатства почвы |
0.28 |
0.37 |
0.04* |
0.21 |
|
Шкала переменности увлажнения |
0.27 |
0.03* |
0.25 |
0.18 |
|
Шкала пастбищной дигрессии |
0.13* |
0.01* |
0.10* |
-0.07* |
Звездочкой помечены недостоверные коэффициенты корреляции.
Средние значения проекций геоботанических описаний на оси DCA-ординации почти не менялись, но их дисперсия вдоль первой оси стала больше (табл. 5). Это видно и на рис. 2: «облако» описаний в 2009 г. заметно более вытянуто вдоль первой оси в сравнение с 1981 г. и особенно – с 1973 г. в сторону уменьшения увлажнения местообитаний.
Таблица 5
Средние значения проекций точек геоботанических описаний на оси DCA-ординации (x ср. ) и величины дисперсий этих проекций (σ2)
|
1-я ось DCA-ординации |
2-я ось DCA-ординации |
||||||
|
Годы |
1973 |
1981 |
2009 |
Годы |
1973 |
1981 |
2009 |
|
x ср. |
3.7 |
3.6 |
3.7 |
x ср. |
1.9 |
1.9 |
1.7 |
|
2 σ |
0.30 |
0.31 |
0.49 |
2 σ |
0.24 |
0.26 |
0.29 |
При сравнении положений «облаков» описаний по их положению в пространстве двух осей с использованием критерия Манна-Уитни было установлено, что они в 1973 и 1981 гг. достоверно не отличались друг от друга. Но в 2009 г. это «облако» по совокупности своих координат достоверно отличалось от 1973 г.
Растительные сообщества . Характеризуя в целом результаты обработки совокупности описаний программой TWINSPAN, можно отметить, что группы учетных площадок оказались в основном расположены в табл. 6 вдоль двух связанных между собой градиентов: увлажнения и пастбищной дигрессии. Слева направо увлажнение падает, пастбищная дигрессия возрастает.
Таблица 6
Синоптическая таблица сообществ, выделенных с помощью программы TWINSPAN
|
Номер группы |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|
Общее количество площадок в группе |
3 |
2 |
6 |
10 |
9 |
25 |
58 |
111 |
1 |
|
в 1973 г. |
1 |
3 |
3 |
2 |
10 |
12 |
44 |
||
|
в 1981 г. |
1 |
2 |
1 |
2 |
3 |
9 |
15 |
42 |
|
|
в 2009 г. |
1 |
2 |
5 |
4 |
6 |
31 |
25 |
1 |
|
|
Среднее значение ступени по шкале У |
105 |
93 |
82 |
88 |
81 |
78 |
73 |
63 |
54 |
|
ЛГ Р ПД |
2 |
2 |
3 |
3 |
3 |
3 |
3 |
3 |
5 |
|
Среднее число видов на площадке |
6 |
9 |
13 |
14 |
21 |
19 |
19 |
19 |
5 |
|
Typha angustifolia |
1004 |
. 1 |
. |
40 : |
. |
. 1 |
. |
. 1 |
. |
|
Sparganium erectum |
671 |
. |
. |
10 ! |
. |
. |
. |
. |
. |
|
Campylium sp. |
. |
505 |
|||||||
|
Lemna trisulca |
672 |
50+ 1 |
. |
. |
. |
. |
. |
. |
. |
|
Solanum kitagawae |
. |
1 004 |
. |
20 |
22 |
. |
. |
. |
. |
|
Sagittaria sagittifolia |
671 |
501 1 |
17 |
70+ i |
. |
.1 |
2 |
. 1 |
. |
|
Polygonum amphibium |
33 |
501 |
67+ |
50+ |
11 |
. |
10 |
1 |
. |
|
Sium latifolium |
33 |
501 |
33 |
10 + |
. |
9 |
. |
. |
|
|
Scirpus lacustris + S . hippolyti |
671 |
. |
. |
60+ |
11 |
. 1 681 |
16 |
1 |
. |
|
Butomus umbellatus |
33 |
50+ |
80+ : |
44 |
4 I |
19 |
12 I |
. |
|
|
Thalictrum flavum |
. |
50 21 ! |
83+ |
30 I |
44 |
16 ! |
10 |
4! |
. |
|
Lythrum virgatum |
33 |
501 |
50+ |
1002 |
671 |
84+ |
88+ |
60+ |
. |
|
Lysimachia vulgaris |
33 |
1002 i |
33 |
10 i |
33 |
12 i |
5 |
1i |
. |
|
Carex acuta |
33 |
1005 |
1005 |
. |
561 |
. |
2 |
. |
. |
|
Stachys palustris |
. |
1002 i |
100+ |
80+ i |
89+ |
44 i |
40 |
4i |
. |
|
Mentha arvensis |
. |
. |
831 |
20 |
56+ |
4 |
26 |
3 |
. |
|
Rubia tatarica |
. |
50+ 1 i |
. |
20 |
56+ |
24 i |
33 |
14 i |
. |
|
Althaea officinalis |
. |
501 |
. |
30 |
56+ |
36 |
21 |
5 |
. |
|
Chenopodium polyspermum |
. |
50+ |
30 |
67+ |
32 ; |
10 |
2 ; |
. |
|
|
Achillea cartilaginea |
. |
. |
67+ |
90+ ! |
67+ |
48 I |
36 |
3! |
. |
|
Cirsium arvense |
. |
. |
67+ |
702 |
891 |
76+ |
81+ |
24 |
. |
|
Xanthium strumarium s.l. |
. |
.i |
33 |
40 i |
78+ |
24 i |
52+ |
51+ i |
. |
|
Vicia cracca |
. |
. |
33 |
20 |
56+ |
36 |
40 |
15 |
. |
|
Alisma lanceolatum + A. plantago-aquatica |
. |
. i |
17 |
80+ i |
12 i |
31 |
2i |
. |
|
|
Sonchus arvensis |
. |
. |
50+ |
. |
56+ |
12 |
12 |
2 |
. |
|
Bidens frondosa + B. tripartita |
. |
. । |
67+ |
1 |
16 + i |
10 |
10 i |
. |
|
|
Convolvulus arvensis |
. |
. |
17 |
. + 60+ |
56+ |
64+ |
74+ |
41 |
. |
|
Inula britannica |
. |
17 |
30 |
44 |
961 : |
97+ |
86+ |
||
|
Senecio jacobaea |
. |
. |
17 |
10 ! |
22 |
36 ! |
45 |
78+ ! |
. |
|
Hierochloë repens |
. |
. |
17 |
20 |
33 |
28 |
62+ |
49 |
. |
|
Galium rubioides |
. |
.i |
17 |
.i |
33 |
76+ i |
47 |
18 i |
. |
|
Euphorbia palustris |
. |
. |
17 |
20 |
. |
64+ |
19 |
12 |
. |
|
Echinochloa crus-galli |
. |
. i |
33 |
. i |
78+ |
12 i |
7 |
1i |
. |
|
Scutellaria galericulata + S. hastifolia |
. |
. |
17 |
20 |
11 |
24 |
52+ |
31 |
. |
|
Eleocharis palustris + E. uniglumis |
. |
. । |
. |
1001 |
44 |
64+ + i |
1003 |
941 ; |
. |
|
Gratiola officinalis |
. |
. |
. |
30 |
44 |
60+ |
40 |
45 |
. |
|
Carex agr. |
. |
20 |
22 |
80 +1 : |
781 |
44 |
|||
|
Phalaris arundinacea |
. |
. |
. |
10 । |
44 |
80+ । |
10 |
. |
. |
|
Phragmites australis |
. |
. |
. |
. |
33 |
76+ |
3 |
. |
. |
|
Bromus inermis |
. |
.i |
. |
.i |
33 |
881 i |
57+ |
96 2 i |
100+ |
|
Polygonum sect. |
. |
. |
. |
. |
33 |
20 |
14 |
22 |
100+ |
|
Asparagus officinalis |
. |
. i |
. |
.i |
56+ |
961 i |
64+ |
53+ + i |
. |
|
Euphorbia esula s.l. |
. |
. |
. |
. |
44 |
32 |
83+ |
84+ |
. |
|
Elymus repens |
. |
. 1 |
. |
. 1 |
. |
64+ : |
22 |
i |
. |
|
Tragopogon brevirostris ssp. podolicus |
. |
. |
. |
. |
. |
24 ! |
22 |
75+ |
. |
|
Galium verum |
. |
16 |
24 |
91+ |
|||||
|
Carex praecox |
. |
. |
. |
. |
. |
12 । |
14 |
761 । |
. |
|
Lotus corniculatus s.l. |
. |
. |
. |
. |
. 1 |
4 |
28 |
70+ |
. |
Eryngium planum
Artemisia austriaca
Carex stenophylla
Artemisia campestris ssp. campestris
i
i i
12 ; 56+ : .
. I 1 I 100+ . 1 2 1002 . I. I 1001
1) В строке «средние значения ступеней шкал Л.Г. Раменского»: У – увлажнение, ПД –
пастбищная дигрессия. 2) Вертикальными линиями указаны разделители 1-4 порядков в
соответствии с алгоритмом TWINSPAN. 3) Встречаемость видов приводится в %. 4) Надстрочные числа – значения медианы обилия растений по шкале В.Б. Голуба (Нешатаев, 2001). 5) Полужирным шрифтом выделены числа в ячейках, в которых значения медианы обилия превышают 1.
Выделенные группы мы интерпретируем следующим образом.
Группа 1. Местообитания прибрежно-водной растительности с доминированием рогоза ( Typha angustifolia ). Были представлены на одной пробной площадке во все годы учетов.
Группы 2 и 3. Сообщества с доминированием осоки острой ( Carex acuta ). В 1981 г. местообитания, занятые этим сообществом, по показателю шкалы Л.Г. Раменского были более влажными, чем в 1973 и 2009 гг. В 1981 г. эти фитоценозы насыщены гидрофитами и гигрофитами, такими как Lemna trisulca , Sagittaria sagittifolia , Lysimachia vulgaris.
Группы 4 и 5. Сырые луга с высокой встречаемостью грубого разнотравья ( Lythrum virgatum , Stachys palustris , Achillea cartilaginea , Cirsium arvense , Alisma lanceolatum + A. plantago-aquatica ). Число площадок с этими сообществами в 2009 г. было почти в два раза больше, чем 1973 и 1981 гг.
Группа 6. Также как и предыдущие две группы, это сырые луга, но, в отличие от них, здесь большую роль во флористическом составе играют злаки: Phalaris arundinacea , Phragmites australis , Bromus inermis , Elymus repens. Фитоценозы этой группы в полтора раза уменьшили представленность в 2009 г. в сравнении с 1973 и 1981 гг. Перечисленные злаки хорошо переносят ежегодное сенокошение.
Группа 7. Влажные луга с доминированием ситнягов ( Eleocharis palustris + E. uniglumis ) с высоким участием в фитоценозах этой группы Lythrum virgatum , Cirsium arvense и Euphorbia esula s.l. Встречаемость этих лугов возросла в два раза в сравнении с 1973 и 1981 гг.
Группа 8. Луга с доминированием костра безостого ( Bromus inermis ) и большой встречаемостью пырея ползучего ( Elymus repens ). По показателю увлажнения относятся к свежелуговому. Эта группа сообществ уменьшила свое участие в 2009 г. в 1.5 раза в сравнении с предыдущими двумя годами учетов.
Группа 9. Сухие луга с доминированием осоки узколистной ( Carex stenophylla) , участием полыни австрийской ( Artemisia austriaca ) и полыни полевой ( A. campestris ssp. campestris ) . Представлены были только на одной площадке в 2009 г. на участке у оз. Бабечье на высокой гриве. В 1973 и 1981 гг. травостой этой площадки относился к 8-й группе растительных сообществ.
ЗАКЛЮЧЕНИЕ
Результаты сопоставления флоры и растительности на ключевых участках северной части Волго-Ахтубинской поймы по данным 1973, 1981 и 2009 гг. позволяют сделать следующие выводы.
Наиболее мощное половодье из трех лет учетов, имевшее место в 1981 г., привело к высокой представленности в фитоценозах с доминированием осоки острой гидрофитов и гигрофитов.
Максимальное количество скота, выпасавшееся в зоне долины Нижней Волги в период 1972-1981 гг., отразилось на травостое участков: в 1981 г. больше всего было растений, характерных для пастбищных угодий.
В последний год учетов на участках, одновременно с уменьшением участия злаков, значительно больше, чем в 1973 и 1981 гг. было представлено грубое разнотравье. Это явление мы приписываем снижению пастбищной нагрузки и нерегулярному сенокошению. Разрастание бурьянистого высокотравья на лугах при прекращении их уборки на сено – это обычный процесс в поймах рек России. Он сопровождается увеличением увлажнения поверхности почвы (Работнов, 1984; Андреев, 1985).
В отличие от результатов геоботанических наблюдений, проведенных на стационарных трансектах в северной части поймы (Старичкова и др., 2009; Сорокин и др., 2010; Иолин и др., 2011), на ключевых участках, расположенных в этом же районе, в последний год учетов не было выявлено, в целом, ни значительной ксерофитизации, ни рудерализации растительности. Связано это с двумя обстоятельствами.
Первым является то, что все участки расположены в центральной части поймы, на которых нет очень высоких грив, характерных для прирусловья больших водотоков. Эти гривы в наибольшей степени подверглись иссушению в результате снижения объемов половодий после зарегулирования водного стока. А поскольку обычно пастбищная нагрузка сильней выражена на более сухих экотопах, где имеется возможность более длительного выпаса скота, то явления пастбищной дигрессии и рудерализации на таких местоположениях проявляются резче.
Второе обстоятельство – это удаленность участков от дорог и населенных пунктов. Трансекты же были проложены вдоль дорог, соединяющих села и города. Вблизи них выпас скота осуществляется с большей интенсивностью, и регулярность сенокошения выше. Следует заметить, что сенокошение и выпас скота являются дополнительными причинами, способствующими иссушению почвы. Ослабление действия этих факторов на ключевых участках в определенной мере нейтрализовала ухудшение увлажнения лугов во время половодий. Тем не менее, на одной из высоких грив на ключевом участке у оз. Бабечье была установлена ксерофитизация травостоя.
Третьей причиной меньшей наблюдаемой динамики травостоя на ключевых участках в сравнении с трансектами является тот факт, что период времени, за который она анализировалась на них, был бóльшим. На трансектах исходными датами учетов являлись 1954-1955 гг., а на ключевых участках – 1973 г.
В заключение можно сказать, что отмеченная в 2009 г. на ключевых участках динамика растительности преимущественно связана с изменениями хозяйственного использования лугов и, в гораздо меньшей степени, с изменениями климатических и гидрологических условий. Более того, эффект влияния этих факторов возможно был скрыт снижением пастбищной нагрузки и нерегулярностью уборки травостоя при сенокошении.
Работа выполнена при финансовой поддержке РФФИ (проект №14-04-00011).
Авторы благодарят Родман Л.С. за предоставление материалов геоботанической съемки стационарных участков в 1959 г.
Список литературы Оценка динамики растительности на ключевых участках в северной части Волго-Ахтубинской поймы
- Андреев Н.Г. Луговедение. М.: Агропромиздат. 1985. 256 с.
- Бондарева В.В., Голуб В.Б. Оценка динамики растительности на ключевых участках средней части Волго-Ахтубинской поймы // Изв. Самарского науч. центра РАН. 2014. Т. 16. № 5 (5). С. 1630-1636.
- Голуб В.Б., Сорокин А.Н., Ивахнова Т.Л. и др. Геоботаническая база данных долины Нижней Волги // Изв. Самарского науч. центра РАН. 2009. Т. 11. № 1 (4). C. 577-582.
- Голуб В.Б., Старичкова К.А., Бармин А.Н. и др. Оценка динамики растительности в дельте Волги // Аридные экосистемы. 2013. Т. 19. № 3 (56), С. 54-64.
- Голуб В.Б., Бондарева В.В., Шитиков В.К., Бармин А.Н., Иолин М.М. Дополнительные данные о динамике засоления почвы и растительности в дельте р. Волги // Аридные экосистемы. 2015. Т. 21. №3 (64), с. 48-55.
