Оценка функциональной активности ядрышковых организаторов хромосом у ели сибирской в различных экологических условиях

Автор: Калашник Надежда Александровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Лесные ресурсы
Статья в выпуске: 3-3 т.15, 2013 года.
Бесплатный доступ
Проведены исследования функциональной активности ядрышковых организаторов хромосом у ели сибирской, произрастающей в различных экологических условиях на территории Южного Урала: в контрастных природных ландшафтах (высокогорья, равнины, поймы и долины рек), в зонах естественных геохимических аномалий, а также при промышленном загрязнении различной интенсивности в сравнении с контролем. Полученные результаты свидетельствуют о повышении функциональной активности ядрышковых организаторов хромосом у ели сибирской как в условиях техногенного загрязнения, так и в природных экстремальных условиях.
Ядрышковые организаторы хромосом, ель сибирская, промышленное загрязнение, экстремальные природные условия, южный урал
Короткий адрес: https://sciup.org/148201901
IDR: 148201901 | УДК: 502.3:
Estimation the functional activity of nucleolar organization chromosomes in Siberian spruce under different ecological conditions
Functional activity of nucleolar organization chromosomes have been studied in Siberian spruce growing in South Urals under different ecological conditions: in contrasting natural landscapes (high mountains, plains, river valleys and floodplains), in zones of natural geochemical anomalies and in areas with different levels of industrial pollution. The results provide evidence for enhancing the technogenic pollution and extreme environmental factors on functional activity of nucleolar organization chromosomes in Siberian spruce.
Текст научной статьи Оценка функциональной активности ядрышковых организаторов хромосом у ели сибирской в различных экологических условиях
По существующим представлениям, ядрышковые организаторы хромосом (ЯОХ) выполняют функции, присущие адаптивным системам. Прежде всего, это участие ЯОХ в жизненно важной функции организма – механизме белкового синтеза. В основе интенсификации любых биосинтетических процессов лежит изменение активности генома, в том числе, той его части, которая ответственна за синтез рРНК [6, 8, 10]. Функциональное состояние генов рРНК возможно оценить путем цитологического изучения ядрышек интерфазного ядра, размеры которых находятся в прямой зависимости от степени активности этих генов [5]. Информативным показателем активности белоксинтезирующей системы в клетке является ядерно-ядрышковое отношение [18]. Изменение ядерно-ядрышково-го отношения свидетельствует об изменении уровня биосинтеза белка. Возрастание объема ядрышка, связанное с усилением биосинтетических процессов в клетке, приводит к уменьшению ядерно-ядрышкового отношения за счет увеличения ядрышкового материала в клетке [11]. Поэтому неудивительно, что именно ядрышковая активность оказалась наиболее показательным цитологическим критерием оценки стрессового воздействия на организм при создании шкалы чувствительности для экологического мониторинга [2].
Исследования нуклеолярных районов хромосом у различных видов хвойных достаточно
многочисленны, подобные работы проводились и на примере ели сибирской [3, 9, 12, 13, 16], где авторами, в основном, изучался полиморфизма ядрышкообразующих хромосом. Однако вопросы исследования функциональной активности ЯОХ у ели сибирской в литературе практически не обсуждались. В настоящей работе представлены результаты исследования функциональной активности ЯОХ у ели сибирской, произрастающей в различных экологических условиях на территории Южного Урала: в контрастных природных ландшафтах (высокогорья, равнины, поймы и долины рек), в зонах естественных геохимических аномалий, а также при промышленном загрязнении различной интенсивности в сравнении с контролем.
Материал и методы. В качестве объектов для исследования выбраны средневозрастные естественные насаждения ели сибирской, произрастающие на территории Челябинской области и Башкортостана, всего исследовано 12 пробных площадей (ПП) из различных экологических условий. На выбранных ПП оценивалось жизненное состояние древостоев согласно классификации В.А. Алексеева [1]. При анализе результатов учитывались климатические условия и геохимические особенности районов исследования, а также комплексная характеристика интенсивности загрязненности территорий по состоянию атмосферного воздуха, водоемов и почвенного покрова [7; 15].
Описание пробных площадей. В районе г. Златоуста одна ПП находится на расстоянии 1,5-2 км от промышленной зоны города, вторая ПП расположена на территории свалки промышленных отходов, обе ПП подвержены многолетнему сильному техногенному загрязнению. В районе промышленной зоны г. Златоуста находятся металлургический комбинат, завод металлоконструкций, абразивный завод и ряд других производств. В районе г. Аша ПП находится в черте города, вблизи от железнодорожного полотна, подвержена также поливалентному техногенному загрязнению со стороны промышленных предприятий города - металлургического и химического заводов. В районе г. Сим ПП находится на окраине города, вблизи (200-300 м) от автотрассы. Помимо крупной автомагистрали, источником загрязнения в городе являются выбросы ОАО «Агрегат». Загрязнение этих ПП можно охарактеризовать как умеренное. В районе пос. Вознесенка ПП находится в регионе, который характеризуется сложным геологолитологическим строением и повышенным содержанием в материнских породах редкоземельных элементов (кадмия, стронция, кобальта, селена, марганца, меди, титана, хрома, ванадия и др.). В районе горы Иремель (высота над уровнем моря 1582 м) ПП находится на верхней границе распространения леса и характеризуется суровыми климатическими условиями произрастания (продолжительным низкотемпературным зимним периодом, резкими колебаниями дневных и ночных температур, сильными ветрами, обильными атмосферными осадками). В районе горы Таганай (высота над уровнем моря 1117 м) ПП находится рядом с Откликным гребнем на верхней границе распространения леса и также отличается суровостью климатических условий, характерных для высокогорий. В районе пос. Веселовка ПП находится на расстоянии 20 км южнее г. Златоуста, в долине реки Ай. В районе пос. Точильный ПП находится на расстоянии 30 км северо-восточнее г. Аша в предгорье Воробьиных гор. ПП в районе пос. Караидель и пос. Павловка находятся вблизи Павловского водохранилища на достаточно большом расстоянии от промышленных предприятий. В районе реки Инзер ПП находится на территории Южноуральского заповедника, в долине реки, на расстоянии 20 км от поселка Инзер (высота над уровнем моря 281 м). Эти ПП определены нами как оптимальные, и использованы в качестве контрольных.
Для насаждений при техногенном загрязнении, когда выбросы загрязнителей значительно превышают ПДК, характерно наличие большого числа суховершинных деревьев и деревьев с «ведьмиными метлами», усыхание боковых побегов, пожелтение или покраснение хвои. В этих условиях состояния насаждений охарактеризованы как «сильно ослабленные» и
«ослабленные». Насаждения в районах высокогорий, уже находящиеся в репродуктивном состоянии, характеризуются низкоростлостью и даже карликовостью, имеют резко выраженные признаки угнетенности в виде покореженных верхушечных и боковых побегов, асинхронности побегообразования и отсутствия охвоения в некоторых частях кроны. В оптимальных и контрольных условиях состояния насаждений определены как «здоровые».
В качестве материала для проведения исследований использована меристематическая ткань проростков семян. В качестве методов использовали общепринятые рекомендации окрашивания ядрышек, в том числе адаптированные применительно к хвойным породам [14, 17]. Давленые препараты изучали при помощи микроскопа ЛЮМАМ Р8, используя объективы х40 или х100. Исследовали с каждой пробной площади не менее 100 клеток (микрофотографии клеток с ядрышками представлены на рис. 1). В результате исследований определяли максимальное и среднее число ядрышек на клетку, а также ядерно-ядрышковые отношения. Статистическую обработку результатов проводили общепринятыми методами [4].
Результаты и их обсуждение. Результаты проведенных исследований показали, что среднее число ядрышек на клетку варьирует незначительно и составляет на различных ПП 5-7 ядрышек. Максимальное же число ядрышек на исследованных ПП варьирует от 10 до 16, причем наиболее высокое значение этого показателя наблюдается в условиях сильного техногенного загрязнения и высокогорий. В этих же условиях наблюдаются более низкие значения ядерно-ядрышковых отношений, они на 3-4 единицы ниже, чем в относительно чистых и оптимальных условиях (чем меньше этот показатель, тем больше объем ядрышек), что, несомненно, определяет тенденцию увеличения активности ядрышкообразующей системы у ели сибирской в стрессовых условиях (табл. 1). Полученные данные согласуются с результатами исследований у ели сибирской ядрышкообразующих районов хромосом, анализ которых показывает наличие высокого полиморфизма по числу и частоте встречаемости и особенно локализации вторичных перетяжек в их кариотипах. Так, М.В. Крук-лис [9] отмечает, что у ели сибирской наблюдается большое число вторичных перетяжек, однако только две вторичные перетяжки имеют постоянную локализацию. По данным Н.С. Медведевой и Е.Н. Муратовой [12] у ели сибирской вторичные перетяжки наблюдаются у многих хромосом, но более часто у пяти пар, а по результатам исследований О.С. Владимировой [3]
у 8 пар хромосом. Е.Н. Муратова [13] также отмечает, что у ели сибирской вторичные перетяжки как с постоянной, так и с непостоянной локализацией наблюдаются в 9 хромосомных парах и указывает, что северные популяции характеризуются более высоким их числом, чем южные.
Таблица 1. Показатели ядрышковой активности у ели сибирской в различных экологических условиях
|
Местонахождение пробной площади |
Число ядрышек на клетку |
Ядерно-ядрышковое отношение |
|||
|
максимальное значение |
среднее значение ±ошибка |
коэффициент вариации |
среднее значение ±ошибка |
коэффициент вариации |
|
|
г. Златоуст** |
12 |
6,13 + 0,12 |
19,58 |
6,44 + 0,28 |
43,48 |
|
г. Златоуст (свалка)** |
14 |
6,19 + 0,16 |
25,85 |
6,42 + 0,29 |
45,17 |
|
г. Аша* |
12 |
6,01 + 0,18 |
29,95 |
7,71 + 0,32 |
41,50 |
|
г. Сим* |
10 |
5,72 + 0,16 |
27,97 |
8,13 + 0,26 |
31,98 |
|
пос. Точильный |
10 |
5,93 + 0,15 |
25,30 |
9,23 + 0,43 |
46,59 |
|
пос. Веселовка |
10 |
5,22 + 0,12 |
22,99 |
9,07 + 0,38 |
41,90 |
|
пос. Вознесенка*** |
12 |
5,33 + 0,14 |
26,27 |
7,34 + 0,25 |
34,06 |
|
пос. Караидель |
10 |
5,88 + 0,16 |
27,21 |
8,18 + 0,21 |
25,67 |
|
пос. Павловка |
10 |
6,14 + 0,13 |
21,17 |
7,07 + 0,35 |
49,50 |
|
**** гора Таганай |
14 |
6,55 + 0,17 |
25,95 |
5,83 + 0,22 |
37,74 |
|
гора Иремель |
16 |
6,72 + 0,19 |
28,27 |
6,02 + 0,29 |
48,17 |
|
река Инзер |
10 |
5,65 + 0,16 |
28,32 |
8,05 + 0,22 |
27,33 |
Примечание: * – умеренное загрязнение; ** – сильное загрязнение; *** – геохимические аномалии; **** – высокогорья; без обозначений – контрольные и оптимальные условия
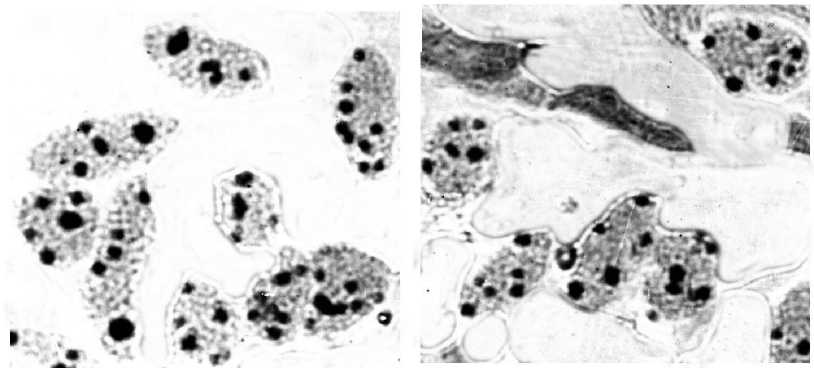
Рис. 1. Микрофотографии меристематических клеток ели сибирской с различным числом ядрышек
Выводы:
-
1. В стрессовых условиях у ели сибирской наблюдается увеличение функциональной активности ЯОХ, то есть активизируются механизмы, обеспечивающие организмам процессы адаптивных изменений.
-
2. Ядрышкообразующая система данного вида является одним из важнейших элементов, с помощью которого осуществляется адаптация организмов в экологически неблагоприятных условиях.
-
3. Исследованные показатели ядрышковой активности могут быть рекомендованы для оценки степени стрессового воздействия на насаждения ели сибирской при мониторинговых наблюдениях.
Список литературы Оценка функциональной активности ядрышковых организаторов хромосом у ели сибирской в различных экологических условиях
- Алексеев, В.А. Диагностика жизненного состояния деревьев и древостоев//Лесоведение. 1989. № 4. С. 51-57.
- Буторина, А.К. Анализ чувствительности различных критериев цитогенетического мониторинга/А.К. Буторина, В.Н. Калаев//Экология. 2000. № 3. С. 206-210.
- Владимирова, О.С. Кариологические особенности ели сибирской Picea obovata Ledeb. из разных мест произрастания//Цитология. 2002. Т. 44. № 7. С. 712-718.
- Вольф, В.Г. Статистическая обработка данных. -М.: Колос, 1966. 255 с.
- Дуброва, Н.А. Ядрышковые организаторы хромосом как адаптивный элемент вида//Журнал общей биологии. 1989. Т. 50. №2. С. 213-217.
- Дуброва, Н.А. Изучение полиморфизма ядрышкообразующих хромосом у видов рода Actaea L. (Ranunculaceae Juss.)/Н.А. Дуброва, Л.А. Малахова//Цитология и генетика. 1980. Т.14. №5. С. 3-8.
- Комплексный доклад о состоянии окружающей среды Челябинской области в 2004 году. -Челябинск: Министерство радиационной и экологической безопасности Челябинской области. Управление Федеральной службы по надзору в сфере природопользования по Челябинской области, 2005. 221 с.
- Корочкин, Л.И. Взаимодействие генов в развитии. -М.: Наука, 1976. 280 с.
- Круклис, М.В. Кариологические особенности Picea obovata Ldb.//Лесоведение. 1971. №2. С. 76-84.
- Кулаева, О.Н. О регуляции экспрессии генов в растительных клетках//Физиология растений. 1978. Т. 25. Вып. 5. С. 909-1008.
- Машкин, С.Н. Сезонная динамика числа и размеров ядрышек и ядерно-ядрышковых отношений у представителей подсемейства сливовых при их интродукции/С.Н. Машкин, М.И. Назарова//Цитология. 1976. Т. 18. № 12. С. 1438-1443.
- Медведева, Н.С. Кариологическое исследование ели сибирской (Picea obovata Ledeb.) из Якутской АССР/Н.С. Медведева, Е.Н. Муратова//Изв. СО АН СССР. Сер. биол. наук. 1987. Вып. 1. № 6. С.15-21.
- Муратова, Е.Н. Кариосистематика семейства Pinaceae Lindl. Сибири и Дальнего Востока: Автореф. дисс. … докт. биол. наук. -Новосибирск, 1995. 32 с.
- Муратова, Е.Н. Методики окрашивания ядрышек для кариологического анализа хвойных//Ботан. журн. 1995. Т. 80, №2. С. 82-85.
- Проблемы экологии: Принципы их решения на примере Южного Урала/Под ред. Н.В. Старовой. -М.: Наука, 2003. 287 с.
- Путенихин, В.П. Ель сибирская на Южном Урале и в Башкирском Предуралье (популяционно-генетическая структура)/В.П. Путенихин, З.Х. Шигапов, Г.Г. Фарукшина. -М.: Наука, 2005. 180 с.
- Сабанеева, Е.В. Специфичность окрашивания ядрышковых организаторов азотнокислым серебром//Цитология. 1989. Т. 31, №1. С. 5-14.
- Шахбазов, В.Г. Некоторые особенности ядрышка и ядра в клетках гибридного лука/В.Г. Шахбазов, Н.Г. Шестопалова//Докл. АН СССР. 1971. Т. 196, № 5. С. 1207-1208.
