Оценка функциональной активности ядрышковых организаторов хромосом у пихты сибирской в различных экологических условиях

Автор: Калашник Надежда Александровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-1 т.18, 2016 года.
Бесплатный доступ
Проведены исследования функциональной активности ядрышковых организаторов хромосом у пихты сибирской, произрастающей в различных экологических условиях на территории Южного Урала: в контрастных природных ландшафтах (равнины, возвышенности, предгорья, поймы и долины рек), а также при промышленном загрязнении различной интенсивности в сравнении с контролем. Полученные результаты свидетельствуют о повышении функциональной активности ядрышковых организаторов хромосом у пихты сибирской, как в условиях техногенного загрязнения, так и в природных экстремальных условиях.
Ядрышковые организаторы хромосом, пихта сибирская, промышленное загрязнение, природные условия, южный урал
Короткий адрес: https://sciup.org/148204462
IDR: 148204462 | УДК: 502.3:
Estimation the functional activity of chromosomes nucleolar organizers at fir Siberian in various ecological conditions
Researches the functional activity of chromosomes nucleolar organizers at fir Siberian, growing in various ecological conditions in the territory of South Ural are conducted: in contrast natural landscapes (plains, heights, foothills, flood plains and valleys of the rivers), and also in case of industrial pollution of various intensity in comparison with control. The received results demonstrate increase of functional activity of chromosomes nucleolar organizers at fir Siberian, both in the conditions of technogenic pollution and in natural extreme conditions.
Текст научной статьи Оценка функциональной активности ядрышковых организаторов хромосом у пихты сибирской в различных экологических условиях
По мнению многих исследователей, ядрышковые организаторы хромосом несут функции, присущие адаптивным системам, поскольку участвуют в жизненно важной функции организма – механизме белкового синтеза. В основе интенсификации любых биосинтетических процессов лежит изменение активности генома, в том числе, той его части, которая ответственна за синтез рРНК [12, 19, 21]. Функциональное состояние генов рРНК возможно оценить путем цитологического изучения ядрышек интерфазного ядра, размеры которых находятся в прямой зависимости от степени активности этих генов [11]. Информативным показателем активности белоксинтезирующей системы в клетке является ядерно-ядрышковое отношение [36]. Изменение ядерно-ядрышкового отношения свидетельствует об изменении уровня биосинтеза белка. Возрастание объема ядрышка, связанное с усилением биосинтетических процессов в клетке, приводит к уменьшению ядерно-ядрышкового отношения за счет увеличения ядрышкового материала в клетке [22]. Следует также отметить, что именно ядрышковая активность оказалась наиболее показательным цитологическим критерием оценки стрессового воздействия на организм при создании шкалы чувствительности для экологического мониторинга [8].
Исследования нуклеолярных районов хромосом у различных видов хвойных достаточно многочисленны, подобные работы проводились на примере сосны обыкновенной [1, 2, 4, 17, 24, 29, 34, 37, 38], ели сибирской [9, 20, 23, 26, 31], лиственницы Сукачева [25, 26, 32], где авторами, в основном, изучался полиморфизм ядрышкообразующих хромосом. На примере этих же хвойных видов проводились исследования функциональной активности ядрышковых организаторов хромосом [14-16]. По пихте сибирской имеются лишь единичные результаты исследования структуры кариотипа в различных частях ареала и условиях произрастания [5-7, 13]. Ядрышкообразующая система у пихты сибирской практически не изучалась, в связи с чем исследования в этом направлении, безусловно, являются актуальными.
Цель работы: исследовать функциональную активность ядрышковых организаторов хромосом у
пихты сибирской, произрастающей в различных экологических условиях на территории Южного Урала: в контрастных природных ландшафтах (равнины, возвышенности, предгорья, поймы и долины рек), а также при промышленном загрязнении различной интенсивности в сравнении с контролем.
Материал и методы. В качестве объектов для исследования выбраны средневозрастные естественные насаждения пихты сибирской, произрастающие на территории Челябинской области и Башкортостана, всего исследовано 9 пробных площадей (ПП) из различных экологических условий. На выбранных ПП оценивалось жизненное состояние древостоев согласно классификации В.А. Алексеева [3]. При анализе результатов учитывались особенности климатических условий районов исследования, а также комплексная характеристика интенсивности загрязненности территорий по состоянию атмосферного воздуха, водоемов и почвенного покрова [18, 30].
Описание пробных площадей. В районе г. Златоуста одна ПП находится на расстоянии 1,5-2 км от промышленной зоны города, вторая ПП расположена на территории свалки промышленных отходов, обе ПП подвержены многолетнему сильному техногенному загрязнению. В районе промышленной зоны г. Златоуста находятся металлургический комбинат, завод металлоконструкций, абразивный завод и ряд других производств. В районе Национального парка «Таганай» ПП находится в предгорье горы Таганай на незначительном удалении от г. Златоуст. В районе г. Аша ПП находится в черте города, вблизи от железнодорожного полотна, подвержена также поливалентному техногенному загрязнению со стороны промышленных предприятий города – металлургического и химического заводов. В районе г. Сим ПП находится на окраине города, вблизи (200-300 м) от автотрассы. Помимо крупной автомагистрали, источником загрязнения в городе являются выбросы ОАО «Агрегат». Загрязнение этих ПП можно охарактеризовать как умеренное. В районе пос. Веселовка ПП находится на расстоянии 20 км южнее г. Златоуста, в долине реки Ай. В районе пос. Точильный ПП находится на расстоянии 30 км северовосточнее г. Аша в предгорье Воробьиных гор. ПП в районе пос. Караидель и пос. Павловка находятся вблизи Павловского водохранилища на достаточно большом расстоянии от промышленных предприятий. Эти
ПП определены нами как оптимальные, и использованы в качестве контрольных.
В целом, для насаждений из условий техногенного загрязнения, когда выбросы загрязнителей значительно превышают ПДК, характерно наличие большого числа суховершинных деревьев, усыхание боковых побегов, пожелтение, а чаще покраснение хвои. В этих условиях состояния насаждений охарактеризованы как «сильно ослабленные» и «ослабленные». В оптимальных и контрольных условиях состояния насаждений определены как «здоровые».
В качестве материала для проведения исследований использована меристематическая ткань проростков семян. В качестве методов использовали общепринятые рекомендации окрашивания ядрышек [33], в том числе адаптированные применительно к хвойным породам [27]. Давленые препараты изучали при помощи микроскопа БИМАМ Р 13, используя окуляры х7, объективы х25 или х40, насадку х1.6 или х 2.5. Исследовали с каждой пробной площади не менее 100 клеток (микрофотографии клеток с ядрышками представлены на рис. 1). В результате исследований определяли максимальное и среднее число ядрышек на клетку, а также показатели ядерно-ядрышковых отношений. Статистическую обработку результатов проводили общепринятыми методами [10].
Таблица 1. Показатели ядрышковой активности у пихты сибирской в различных экологических условиях
|
Местонахождение ПП |
Число ядрышек на клетку |
Ядерно-ядрышковое отношение |
|||
|
максимальное значение |
среднее значение ±ошибка |
коэффициент вариации |
среднее значение ±ошибка |
коэффициент вариации |
|
|
г. Златоуст (7-ой участок) ** |
11 |
5,51±0,20 |
36,30 |
4,42±0,15 |
33,94 |
|
г. Златоуст (свалка) ** |
14 |
5,94±0,23 |
38,72 |
4,14±0,12 |
28,99 |
|
Национальный парк "Таганай" *** |
10 |
5,02±0,18 |
35,86 |
5,42±0,16 |
29,52 |
|
пос. Веселовка |
9 |
4,45±0,14 |
31,46 |
6,57±0,19 |
28,92 |
|
г. Аша * |
13 |
5,15±0,19 |
36,89 |
5,44±0,18 |
33,09 |
|
г. Сим * |
12 |
4,94±0,15 |
30,36 |
5,93±0,20 |
33,73 |
|
пос. Точильный |
8 |
4,52±0,16 |
35,40 |
6,26±0,23 |
36,74 |
|
пос. Караидель |
10 |
4,42±0,13 |
29,41 |
5,94±0,17 |
28,62 |
|
пос. Павловка |
10 |
5,22±0,19 |
36,40 |
6,17±0,18 |
29,17 |
Примечание: * - умеренное загрязнение; ** - сильное загрязнение; *** - предгорья; без обозначений - контрольные и фоновые условия
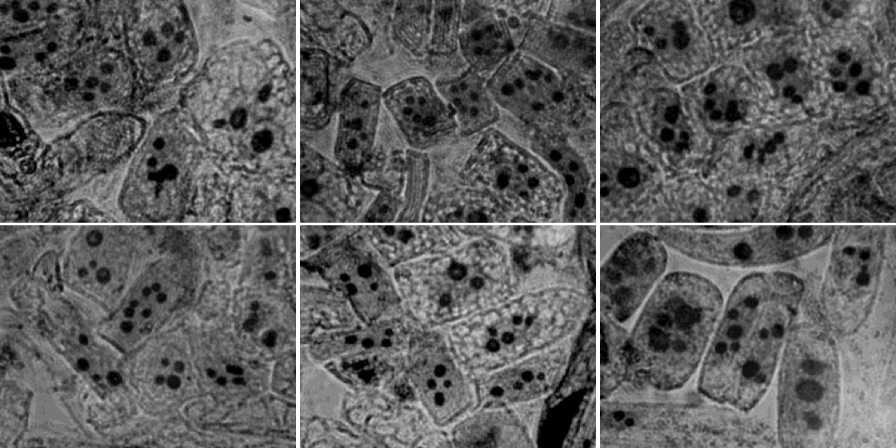
Рис. 1. Микрофотографии меристематических клеток пихты сибирской с различным числом ядрышек
Результаты и их обсуждение. Результаты проведенных исследований показали, что среднее число ядрышек на клетку варьирует незначительно и составляет на различных ПП 4-5 ядрышек. Максимальное же число ядрышек на исследованных ПП варьирует от 8 до 14, причем наиболее высокое значение этого показателя наблюдается в условиях сильного и умеренного техногенного загрязнения . В этих же условиях наблюдаются более низкие значения ядерно-ядрышковых отношений, они на 1-2 единицы ниже, чем в относительно чистых и оптимальных условиях (чем меньше этот показатель, тем больше объем ядрышек), что, несомненно, определяет тенденцию увеличения актив- ности ядрышкообразующей системы у пихты сибирской в стрессовых условиях (таблица). Полученные данные согласуются с результатами многочисленных исследований ядрышкообразующих районов хромосом других хвойных видов (сосны обыкновенной, ели сибирской и лиственницы Сукачева), анализ которых показывает наличие высокого полиморфизма по числу, частоте встречаемости и локализации вторичных перетяжек в их кариотипах [1, 2, 4, 9, 17, 20, 23-26, 29, 31, 32, 34,37,38]. Кроме того, некоторые исследователи отмечают отчетливую тенденцию увеличения числа нуклеолярных районов хромосом в экологически неблагоприятных условиях (болота, высокогорья, север- ные широты, промышленное загрязнение) [24, 26, 28, 35, 38]. Приведенные результаты исследования функциональной активности ядрышковых организаторов хромосом у пихты сибирской согласуются с данными полученными нами ранее на примере сосны обыкно- венной, ели сибирской и лиственницы Сукачева, которые также свидетельствуют о повышении ядрышковой активности в природных экстремальных условиях и условиях техногенного загрязнения [14-16].
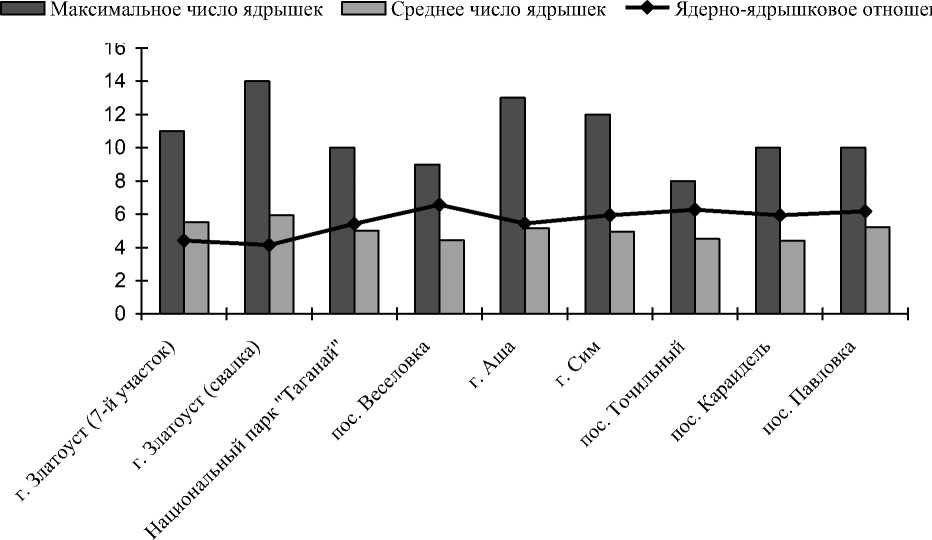
Рис. 2. Показатели ядрышковой активности пихты сибирской в различных экологических условиях
Выводы:
-
1. В стрессовых условиях у пихты сибирской наблюдается некоторое увеличение функциональной активности ядрышковых организаторов хромосом, то есть активизируются механизмы, обеспечивающие организмам процессы адаптивных изменений.
-
2. Ядрышкообразующая система у исследованного вида является одним из важных элементов, с помощью которого осуществляется адаптация организмов в экологически неблагоприятных условиях.
-
3. Исследованные показатели ядрышковой активности могут быть рекомендованы для оценки степени стрессового воздействия на насаждения пихты сибирской при мониторинговых наблюдениях.
Список литературы Оценка функциональной активности ядрышковых организаторов хромосом у пихты сибирской в различных экологических условиях
- Абатурова, Г.А. Кариотипы сосны обыкновенной в европейской части СССР//Научные основы селекции хвойных древесных пород. -М.: Наука, 1978. С. 66-82.
- Абатурова, Г.А. Кариологическая характеристика популяций сосны обыкновенной в азиатской части СССР/Г.А. Абатурова, О.П. Шершукова//Цитология и генетика. 1981. Т. 15. № 1. С. 18-22.
- Алексеев, В.А. Диагностика жизненного состояния деревьев и древостоев//Лесоведение. 1989. № 4. С. 51-57.
- Бударагин, В.А. Анализ кариотипов изолированных популяций сосны обыкновенной (Pinus sylvestris L.) в Северном и Центральном Казахстане//Генетика. 1973. Т. 9. № 9. С. 41-52.
- Бударагин, В.А. Кариотип пихты сибирской Казахстанского Алтая//Цитология. 1972. Т. 14., №1. С. 130-133.
- Бударагин, В.А. Кариотип пихты сибирской Джунгарского Алатау. -Леса и древесные породы Северного Казахстана. -Л.: Наука, 1974. С. 81-84.
- Бударагин, В.А. Кариотипы основных хвойных видов Казахстана//Тр.Каз.НИИЛХа. -Алма-Ата: Кайнар, 1980. Т.2: Защитное лесоразведение и вопросы селекции в Северном Казахстане. С. 116-122.
- Буторина, А.К. Анализ чувствительности различных критериев цитогенетического мониторинга/А.К. Буторина, В.Н. Калаев//Экология. 2000. № 3. С. 206-210.
- Владимирова, О.С. Кариологические особенности ели сибирской Picea obovata Ledeb. из разных мест произрастания//Цитология. 2002. Т. 44. № 7. С. 712-718.
- Вольф, В.Г. Статистическая обработка данных. -М.: Колос, 1966. 255 с.
- Дуброва, Н.А. Ядрышковые организаторы хромосом как адаптивный элемент вида//Журнал общей биологии. 1989. Т. 50. №2.С. 213-217.
- Дуброва, Н.А. Изучение полиморфизма ядрышкообразующих хромосом у видов рода Actaea L. (Ranunculaceae Juss.)/Н.А. Дуброва, Л.А. Малахова//Цитология и генетика. 1980. Т.14. №5. С.3-8.
- Калашник, Н.А. Характеристика естественных насаждений хвойных видов Южного Урала по цитогенетическим показателям//Вестник Оренбург. гос. ун-та. 2009. № 6. С. 157-160.
- Калашник, Н.А. Оценка функциональной активности ядрышковых организаторов хромосом у ели сибирской в различных экологических условиях//Известия Самарского научного центра Российской академии наук. 2013. Т.15, № 3(3). С. 1049-1052.
- Калашник, Н.А. Оценка функциональной активности ядрышковых организаторов хромосом у лиственницы Сукачева в условиях техногенного загрязнения//Вестник Оренбургского госуд. университета. 2013. № 10 (159). С. 195-197.
- Калашник, Н.А. Функциональная активность ядрышковых организаторов хромосом у сосны обыкновенной в условиях техногенного загрязнения//Известия Самарского научного центра Российской академии наук. 2013. Т.15, №3(4). С. 1302-1305.
- Кириченко, О.И. Изменчивость морфологических и кариологических признаков некоторых популяций сосны обыкновенной (Pinus sylvestris L.) на Украине: Автореф. дисс. … к.б.н. -Воронеж, 1984. 20 с.
- Комплексный доклад о состоянии окружающей среды Челябинской области в 2004 году. Челябинск: Министерство радиационной и экологической безопасности Челябинской области. Управление Федеральной службы по надзору в сфере природопользования по Челябинской области. 2005. 221 с.
- Корочкин, Л.И. Взаимодействие генов в развитии. -М.: Наука, 1976. 280 с.
- Круклис, М.В. Кариологические особенности Picea obovata Ldb.//Лесоведение. 1971. №2. С. 76-84.
- Кулаева, О.Н. О регуляции экспрессии генов в растительных клетках//Физиология растений. 1978. Т. 25. Вып. 5. С. 909-1008.
- Машкин, С.Н. Сезонная динамика числа и размеров ядрышек и ядерно-ядрышковых отношений у представителей подсемейства сливовых при их интродукции/С.Н. Машкин, М.И. Назарова//Цитология. 1976. Т.18. № 12. С. 1438-1443.
- Медведева, Н.С. Кариологическое исследование ели сибирской (Picea obovata Ledeb.) из Якутской АССР/Н.С. Медведева, Е.Н. Муратова//Известия СО АН СССР. Сер. биол. наук. 1987. Вып. 1. № 6. С.15-21.
- Муратова, Е.Н. Кариологическое исследование болотных и суходольных популяций сосны обыкновенной (Pinus sylvestris L.)/Е.Н. Муратова, Т.С. Седельникова//Экология. 1993. № 6. С.41-50.
- Муратова, Е.Н. Кариологическое исследование лиственницы Сукачева (Larix sukaczewii N. Dyl.): Нуклеолярные районы и структурные перестройки/Е.Н. Муратова, Н.Е. Чубукина//Цитология и генетика. 1985. Т. 19. № 6. С. 419-425.
- Муратова, Е.Н. Кариосистематика семейства Pinaceae Lindl. Сибири и Дальнего Востока: Автореф. дисс. … д.б.н. -Новосибирск, 1995. 32 с.
- Муратова, Е.Н. Методики окрашивания ядрышек для кариологического анализа хвойных//Ботан. журнал. 1995. Т. 80, №2. С. 82-85.
- Особенности формирования популяций сосны обыкновенной. -М.: Наука, 1984. 128 с.
- Правдин, Л.Ф. Сосна обыкновенная. Изменчивость, внутривидовая систематика и селекция. -М.: Наука, 1964. 190 с.
- Проблемы экологии: Принципы их решения на примере Южного Урала/Под.ред. Н.В. Старовой. -М.: Наука, 2003. 287 с.
- Путенихин, В.П. Ель сибирская на Южном Урале и в Башкирском Предуралье (популяционно-генетическая структура)/В.П. Путенихин, З.Х. Шигапов, Г.Г. Фарукшина. -М.: Наука, 2005. 180 с.
- Путенихин, В.П. Лиственница Сукачева на Урале: изменчивость и популяционно-генетическая структура/В.П. Путенихин, Г.Г. Фарукшина, З.Х. Шигапов. -М.: Наука, 2004. 276 с.
- Сабанеева, Е.В. Специфичность окрашивания ядрышковых организаторов азотнокислым серебром//Цитология. 1989. Т. 31. №1. С. 5-14.
- Сунцов, А.В. Цитогенетика и эмбриология сосны обыкновенной в изолированных популяциях Центральной Тувы: Автореф. дисс. … к.б.н. -Красноярск, 1984.16 с.
- Шафикова, Л.М. Характеристика кариотипа сосны обыкновенной при промышленном загрязнении/Л.М. Шафикова, Н.А. Калашник//Лесоведение. 2000. № 2. С. 30-36.
- Шахбазов, В.Г. Некоторые особенности ядрышка и ядра в клетках гибридного лука/В.Г. Шахбазов, Н.Г. Шестопалова//Докл. АН СССР. 1971. Т.196. № 5. С. 1207-1208.
- Шишниашвили, Р.М. Кариотип сосны Сосновского//Цитология. 1968. Т.10. № 2. С. 255-258.
- Kalashnik, N. Karyotype varyability of Scots pine (Pinus sylvestris L.) in Ural mountain populations/N. Kalashnik, N. Starova//Cytogenetic studies of forest trees and shrub species. Contributions by member of the IUFRO Cytogenetic Working Party/Eds. by Borzan Z. and Schlarbaum S.E. -Zagreb, 1997. P. 131-147.
