Оценка генетического потенциала оленей ненецкой породы с целью создания технологий направленных селекционных процессов в северном оленеводстве

Автор: Максимчик М.А.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Растениеводство
Статья в выпуске: 1 (77), 2025 года.
Бесплатный доступ
Северное оленеводство - ведущая отрасль животноводства на Крайнем Севере России, которая является одной из древнейших форм хозяйственной деятельности и источником доходов коренных народов. Рентабельность оленеводства, как и любой отрасли животноводства, во многом зависит от эффективности селекционно-племенной работы. Несмотря на огромные достижения в генетике и селекции, информационных технологий, племенная работа в северном оленеводстве ведется устаревшими, традиционными методами. Для дальнейшего развития оленеводства необходимо совершенствовать уровень племенной работы за счет применения современных методов селекции и разведения. Такими подходами являются внедрение и использование в оленеводстве маркер-ассоциированной и геномной селекций. Генофонды популяций северных оленей формируются в результате микроэволюционных процессов, в основном дрейфа генов при реализации случайных спариваний особей, давления естественного отбора и антропогенных технологических факторов. В результате миграции особей из других стад и хозяйств, скрещивания с дикими особями, неконтролируемой элиминации животных генетические процессы в популяциях трудно поддаются прогнозу. Тем не менее для оценки генетического разнообразия, минимизации инбридинга в процессе разведения возникает необходимость проведения генетического анализа популяций (пород, стад) северных оленей. При этом выборочная оценка должна осуществляться прижизненно преимущественно по особям, которые представляют репродуктивную ценность и внесут существенный вклад в формирование будущего генофонда. В представленной статье проведена оценка внутрипопуляционной генетической дифференциации стада северных оленей ненецкой породы.
Северные олени, ненецкая порода, генотип, генетическая дифференциация, микросателлиты, селекция, фенотип
Короткий адрес: https://sciup.org/149147831
IDR: 149147831 | УДК: 636.294 | DOI: 10.19110/1994-5655-2025-1-64-74
Evaluation of the genetic potential of the Nenets reindeer breed for creation of the technologies of directed selection processes in northern reindeer herding
Northern reindeer herding is the leading branch of animal husbandry in the Russian Far North, which is one of the oldest forms of economic activity and a source of income for indigenous peoples. The profitability of reindeer herding, like any branch of animal husbandry, largely depends on the efficiency of selection and breeding work. In spite of the enormous achievements in genetics and selection, in information technologies, the breeding work in northern reindeer herding is carried out by outdated common methods. The further development of reindeer herding requires improving the level of breeding work through the use of modern selection and breeding methods. The new approaches include introduction and use of marker-associated and genomic selection in reindeer herding. The gene pools of reindeer populations are formed as a result of microevolutionary processes, mainly gene drift against random mating of individuals, pressure of natural selection, and anthropogenic technological factors. As a result of migration of individuals from other herds and farms, crossbreeding with wild individuals, uncontrolled elimination of animals, genetic processes in populations are difficult to predict. However, to assess the genetic diversity and minimize inbreeding, it is necessary to conduct the genetic analysis of reindeer populations (breeds, herds). Sample estimation should be carried out live-animal, mainly for individuals that are reproduction-valuable and will make a significant contribution to the formation of the future gene pool. This article gives an assessment of intrapopulation genetic differentiation of herd of the Nenets reindeer breed.
Текст научной статьи Оценка генетического потенциала оленей ненецкой породы с целью создания технологий направленных селекционных процессов в северном оленеводстве
Оленеводство на Российском Севере на протяжении многих веков служит ключевым источником мяса, шерсти, меха и кожевенных материалов. Домашние северные олени, разводимые местным населением, стали основным средством передвижения, что дало возможность освоить удаленные северные районы. В медицине используются различные продукты оленеводства, такие как рога и панты, сыворотка крови, органы внутренней секреции и прочие внутренние органы.
Северное оленеводство в России, как и любая другая отрасль сельского хозяйства, нуждается в поддержке со стороны государства и сталкивается с нехваткой квалифицированных специалистов, особенно в области селекционно-племенной работы.
Цель исследований – получить экспериментальные данные для создания технологий направленной селекции в северном оленеводстве.
Основные задачи: провести анализ перспективности развития северного оленеводства в Ямало-Ненецком автономном округе (ЯНАО) и Арктической зоне (АЗ) Российской Федерации (РФ); изучить структуру стада и фенотипические характеристики различных половозрастных групп северных оленей ненецкой породы; дать генетическую оценку экспериментального стада северных оленей Ямальской опытной станции (ЯОС).
Материалы и методы
Работа проведена в Приуральском районе ЯНАО на опытном (экспериментальном) стаде домашних северных оленей (ненецкая порода уральского экотипа) ЯОС – обособленного подразделения ФГБУН ФИЦ Тюменского научного центра СО РАН. По основным инфекционным заболеваниям опытное стадо благополучно имеет многолетний постоянный маршрут каслания в Приуральском районе. При проведении исследований использованы архивные материалы и литературные источники, материалы исполнительных органов государственной власти ЯНАО в области статистики и сельского хозяйства, а также сведения из ранее проведенных научно-исследовательских работ по соответствующей тематике. Для выявления тенденций изменения поголовья оленей в ЯНАО и АЗ РФ применялись методы анализа, синтеза, сравнения и обобщения. Для наглядного представления статистической информации применяли графический и табличный методы. Информационной базой исследования послужили данные Федеральной службы государственной статистики. Методика исследований также включала совокупность зоотехнических, генетических, биохимических, селекционных, ветеринарных и статистических методов. Популяционно-генетические параметры и хозяйственно-полезные признаки оценивали классическими методами с использованием корреляционного, регрессионного и дисперсионного анализов.
Исследованию подлежали северные олени в возрасте от 6 месяцев до 7 лет. Возраст определяли по состоянию зубов, внешнему виду, а также данным зоотехнического учета. Оценка экстерьера и упитанности – глазомерно, согласно «Инструкции по бонитировке северных оленей». Все обследованные олени имели индивидуальные ушные номера (телятам присвоили номера при бонитировке). Для определения фенотипических особенностей оленей проводили взвешивание на площадочных электронных весах. С помощью мерных циркуля и палки, а также ленты устанавливали промеры с точностью до 1 см: высота в холке и крестце, косая длина туловища, обхват пясти, глубина груди, ширина груди за лопатками, обхват груди, косая длина зада, ширина в маклоках и седалищных буграх, длина и ширина головы.
Во время бонитировки от животных брали биологический материал (кусочек ушного хряща) для последующих генетических исследований. Отбор, упаковку и доставку собранных образцов для генетического анализа и оценки генетического разнообразия домашних северных оленей осуществляли в герметичных контейнерах, биксах и сумках-холодильниках с сопроводительными документами, оформленными в соответствии с ГОСТ 57343-2016, «Правилами генетической экспертизы племенного материала крупного рогатого скота» (рассмотрены и одобрены на Научно-техническом совете Минсельхоза России (протокол № 27 от 29 октября 2002 г.)).
При проведении исследований использовали оборудование ЦКП «Биоресурсы и биоинженерия сельскохозяйственных животных» ФГБНУ ФИЦ ВИЖ им. Л. К. Эрнста, в том числе анализ отобранного генетического материала. Выделение ДНК проводили с помощью наборов для выделения геномной ДНК серии «ДНК-Экстран» (ЗАО «Синтол», Россия), в соответствии с протоколом фирмы-производителя. Анализ полиморфизма осуществляли по отношению к девяти STR-локусам: NVHRT21, NVHRT24, NVHRT76, RT1, RT6, RT7, RT9, RT27, RT30. Полимерную цепную реакцию (ПЦР) проводили в конечном объеме 15 мкл. Реакционная смесь состояла из 1,5 мкл 10х, 1,5 мкл 2мМ раствора dNTPs, 10 мМ смеси праймеров, 0,1 мкл (1UE) Таq-полимеразы («Диалат Лтд», Россия), к которой добавляли 1 мкл (50-100 нг) геномной ДНК. Состав ПЦР буфера: 16,6 мМ (NH4)2SO4, 67,7 мМ Трис-HCl (pH = 8,8), 0,1 объема Tween 20. После начальной денатурации (95º С, 7 мин) проводили 40 циклов амплификации в следующем температурно-временном режиме: 95º С, 60 с; 58º С, 60 с; 72º С, 60 с. Амплификацию выполняли на термоциклере Labcycler (SensoQuest, Германия). Фрагменты исследовали на генетическом анализаторе ABI3130xl («Applied Biosystems», США). Размеры аллелей определяли с помощью программного обеспечения Gene Mapper v. 4 («Applied Biosystems», США). Для обработки результатов анализа формировали матрицу генотипов в формате Microsoft Excel.
Reindeer» (ООО «Гордиз», Россия) методом мультиплексного ПЦР-анализа с последующей детекцией флуоресцентно-меченых фрагментов в условиях капиллярного электрофореза. Специфичность генотипирования проверяли, используя контрольные образцы, входящие в состав набора.
На основе установленного микросателлитного профиля по 16 локусам исследуемую выборку северных оленей в программе Structure 2.3.4 (Pritchard Lab, Stanford University, USA) подразделяли от 2 до 10 кластеров. Уровень кластеризации, обеспечивающий максимальную внутреннюю однородность и внешнюю дифференциацию, использовали для деления животных на группы в зависимости от апостериорной вероятности (Q) принадлежности к кластерам. Из особей, вероятность членства которых в конкретном кластере была максимальной, сформировали группы. С применением программы GenAlEx 6.503 [1] по группам, выделенным в каждом кластере, определили основные генетические параметры: число наблюдаемых (NA) и эффективных (NE) аллелей, индекс Шеннона (I), уровень наблюдаемой (HO) и ожидаемой (HE) гетерозиготности, индексы фиксации (Fis, Fst), число приватных аллелей, их частоту (q) и сумму частот (Σq). Генетические дистанции между кластерами (k) и популяцией в целом (Pop) рассчитали различными способами: DN, uDN, Gst, G’stN, G’stH, G’’st, Dest [2, 3].
Статистический анализ проведен путем вычисления среднего арифметического и стандартной ошибки, досто-
Таблица 1
Рекомендуемая структура стада северных оленей для товарных хозяйств [5]
Таблица 2
Table 1
Recommended structure of the reindeer herd for commercial farms [5]
|
Половозрастные группы |
В процентах к общему количеству оленей на начало года |
|
|
Короткие кочевки |
Длинные кочевки |
|
|
Важенки |
53 |
52 |
|
Нетели |
10 |
10 |
|
Телеваженки |
12 |
10 |
|
Телехоры |
6 |
6 |
|
Третьяки |
4 |
4 |
|
Быки-производители |
3 |
3 |
|
Быки-кастраты |
12 |
15 |
Половозрастная структура стада северных оленей
Ямальской опытной станции на август 2024 года
Table 2
Age and sex structure of the reindeer herd at the Yamal Experimental Station for August 2024
Результаты и их обсуждение
Половозрастная структура и фенотипическая характеристика экспериментального стада северных оленей Ямальской опытной станции
Северное оленеводство отличается замкнутым циклом воспроизводства, благодаря чему в каждом стаде представлены все возрастные и половые категории животных. Ключевым экономическим аспектом для достижения максимального валового производства и эффективного воспроизводства поголовья является правильное соотношение различных половозрастных групп. Структура стада во многом зависит от срока использования взрослого поголовья (самцов, важенок и ездовых быков). В связи с этим, на первом этапе была проведена оценка структуры стада ЯОС с рекомендуемыми показателями (табл. 1 и 2).
Анализ половозрастной структуры стада свидетельствует, что в хозяйстве наблюдается дефицит маточного поголовья. Так, суммарное количество важенок и нетелей составляет 47,6 %, что ниже рекомендуемого показателя на 14,4 %. Указанное несоответствие обусловлено увеличением количества быков-кастратов (на 3,7 %), третьяков (на 4,5) и телят-самцов (на 5,3 %). В свою очередь, смещение половой структуры стада в сторону преобладания самцов негативно сказывается на количестве получаемого молодняка, а также увеличивает конкуренцию между производителями. Поэтому одним из подходов улучшения селекционно-племенной работы в стаде является оптимизация его половозрастной структуры путем своевременной направленной выбраковки тех или иных особей.
В табл. 3 представлена фенотипическая характеристика стада северных оленей ненецкой породы ЯОС по результатам бонитировки, проведенной в 2023 г. Анализируя цифровые значения, можно констатировать, что важенки, по сравнению с хорами, имеют меньшую на 1,9 см (р ≤ 0,01) длину головы, на 4,5 см (р ≤ 0,001) высоту в холке, на 4,9 см высоту в крестце (р ≤ 0,001), на 0,8 см обхват пясти (р ≤ 0,001) и на 2,4 см длину таза (р ≤ 0,05).
Самцы второго года (третьяки) уступали хорам лишь по ширине головы (на 1,0 см; р ≤ 0,001) и груди (на 4,1 см; р ≤ 0,001). Таким образом, можно заключить, что третьяки по ряду показателей соответствуют взрослым животным и представляют физиологически зрелых особей. Вместе с этим, необходимо учитывать, что на момент проведения бонитировки хоры находятся в истощенном состоянии после гона и не отражают истинного потенциала набора массы, поэтому, на наш взгляд, для определения генетических возможностей мясной продуктивности также имеет целесообразность фенотипическая оценка быков-кастратов.
Телята, рожденные в 2023 г., не имели достоверных различий в зависимости от пола, а их живая масса составляла 53–56 % от массы взрослых особей. Таким образом, можно констатировать, что основной набор массы тела у молодняка происходит в первые полгода постнаталь-
Таблица 3
Фенотипическая характеристика северных оленей Ямальской опытной станции по результатам бонитировки в 2023 году
Table 3
Phenotypic characteristics of reindeer at the Yamal Experimental Station by the quality indicators in 2023
|
Показатель |
Хоры (n=10) |
Третьяки (n=10) |
Телехоры (n=10) |
Важенки (n=40) |
Телеваженка (n=10) |
|
Длина головы, см |
35,6±0,7 |
35,9±0,7 |
27,1±0,5 |
33,7±0,2** |
27,2±0,5 |
|
Ширина головы, см |
12,0±0,2 |
11,0±0,2 *** |
9,7±0,3 |
11,4±0,1 |
9,5±0,4 |
|
Высота в холке, см |
97,5±0,4 |
97,4±1 |
85,7±1,7 |
93,0±0,5 *** |
83,4±1,2 |
|
Высота крестец, см |
97,3±0,7 |
96,6±1,3 |
84,7±1,6 |
92,4±0,4 *** |
80,6±1,3 |
|
Ширина груди, см |
24,5±0,9 |
20,4±0,4 *** |
17,8±1,1 |
22,4±0,2 *** |
16,8±0,7 |
|
Глубина груди, см |
37,0±1,1 |
36,1±1,1 |
29,0±1,1 |
36,9±0,3 |
30,6±0,7 |
|
Обхват груди, см |
113,1±2,7 |
112,9±1,0 |
93,6±1,5 |
112,7±0,7 |
92,5±1,3 |
|
Косая длина туловища, см |
94,1±2,3 |
98,8±2,6 |
75,1±3,4 |
95,8±0,7 |
77,0±2,1 |
|
Обхват пясти, см |
12,0±0,3 |
11,9±0,2 |
9,2±0,4 |
11,2±0,1 *** |
20,3±10,7 |
|
Полуобхват таза, см |
34,9±1,4 |
37,4±1,5 |
30,5±0,9 |
34,2±0,5 |
31,4±1,7 |
|
Ширина в маклаках, см |
21,8±0,5 |
20,4±0,4 |
16,2±0,5 |
22,1±0,2 |
16,5±0,5 |
|
Ширина в седалищных буграх, см |
11,3±0,3 |
10,7±0,3 |
9,9±0,3 |
11,6±0,2 |
20,9±11,6 |
|
Ширина в тазобедренном сочленении, см |
23,0±1,1 |
23,0±1,0 |
18,1±0,5 |
23,8±0,2 |
19,7±1,0 |
|
Длина таза, см |
33,1±1,1 |
31,1±0,7 |
22,9±1,5 |
30,7±0,4* |
24,0±0,7 |
|
Живая масса, кг |
85,4±4,9 |
83,6±4,1 |
45,5±1,6 |
80,0±1,2 |
45,6±1,7 |
Примечание. Различия достоверны: *р ≤ 0,05; **р ≤ 0,01; ***р ≤ 0,001 по отношению к хорам.
Note. Differences are reliable at * р ≤ 0.05; ** р ≤ 0.01; *** р ≤ 0.001 for males.
ного онтогенеза, индивидуальная вариабельность которого зависит от ряда факторов: времени рождения (ранний или поздний отел), молочности важенки, состояния здоровья как матери, так и самого теленка, генетической наследственности и т. д. Стоит отметить, что для оценки истинного генетического потенциала животных указанные паратипические факторы в условиях северного оленеводства практически невозможно в полной мере учесть и исключить, поэтому без должного племенного учета внедрение современных подходов селекции северных оленей невозможно.
В табл. 4 представлены результаты бонитировки телят, рожденных в 2024 г. Установлено, что самцы превосходят телеваженок на 0,8 см по длине головы (р ≤ 0,05), на 2,1 см по высоте в холке (р ≤ 0,05), на 2,2 см по ширине в тазобедренном сочленении (р ≤ 0,05) и на 2,6 кг по живой массе (р ≤ 0,05). Сравнивая результаты бонитировки за последние два года, можно заключить, что телята, рожденные в 2024 г., по ряду показателей уступают молодняку, полученному в 2023 г. Так, масса телеваженок была ниже на 8,7 кг (р ≤ 0,001), а телехоров – на 5,6 кг (р ≤ 0,001). Основной причиной такой значительной разницы, вероятно, стали неблагоприятные климатические условия во время отела, затянувшаяся весна и поздние сроки вегетации растительности, что не дало возможности молодым животным достаточно набрать массу.
Вместе с этим, стоит отметить, что при рассмотрении показателей между индивидуу- мами выявлена значительная вариабельность признаков. Так, по массе тела среди телеваженок наблюдались различия, достигающие 73,6 % (диапазон находился в пределах от 26,5 до 46,0 кг), а по телятам-самцам – до 94,8 % (диапазон в пределах от 26,7 до 52,0 кг). Полученные данные требуют глубокого анализа для выявления факторов, приводящих к такому значительному расхождению по массе тела.
Согласно полученным промерам, у молодняка были рассчитаны основные индексы телосложения (табл. 5). Установлено, что самки обладают более растянутым телосложением при меньшей компактности и массивности. Тазо-грудной индекс как косвенный маркер мясной продуктивности был более выраженным у самцов, которым также была свойственна большая шилоза-дость.
В целом, проведенный фенотипический анализ свидетельствует, что исследуемая популяция имеет типичное для уральского экотипа ненецкой породы телосложение.
Генетическая характеристика экспериментального стада северных оленей в за висимости от половой принадлежности
Спаривание одомашненных северных оленей, как правило, осуществляется случайным образом, а генофонд их популяций формируется под влиянием естественных условий среды. Миграция особей из различных стад и хозяйств, а также возможность скрещивания с дикими оленями и неконтролируемая элиминация животных создают сложности в прогнозировании генетических процессов в популяциях. Поэтому для анализа генетического раз-
Таблица 4
Фенотипическая характеристика телят ненецкой породы Ямальской опытной станции по результатам бонитировки в 2024 году
Table 4
Phenotypic characteristics of calves of the Nenets breed at the Yamal Experimental Station by the quality indicators in 2024
|
Показатель |
Телехоры (n=33) |
Телеваженки (n=47) |
Разница |
Достоверность (Р) |
|
Длина головы, см |
26,5±0,4 |
25,7±0,2 |
0,8 |
0,042 |
|
Ширина головы, см |
9,8±0,1 |
9,8±0,1 |
0,0 |
0,945 |
|
Высота в холке, см |
78,9±0,7 |
76,8±0,6 |
2,1 |
0,041 |
|
Высота крестец, см |
77,9±1,5 |
76,0±1,0 |
1,9 |
0,296 |
|
Ширина груди, см |
20,6±0,5 |
20,3±0,4 |
0,3 |
0,653 |
|
Глубина груди, см |
30,9±0,4 |
30,1±0,3 |
0,8 |
0,110 |
|
Обхват груди, см |
90,7±1,6 |
90,5±0,7 |
0,2 |
0,898 |
|
Косая длина туловища, см |
70,5±0,7 |
70,7±0,8 |
-0,2 |
0,871 |
|
Четверть обхвата таза, см |
10,6±0,4 |
10,1±0,3 |
0,5 |
0,275 |
|
Ширина в маклаках, см |
18,6±0,8 |
18,7±0,7 |
0,0 |
0,973 |
|
Ширина в седалищных буграх, см |
9,0±0,3 |
9,3±0,2 |
-0,3 |
0,473 |
|
Ширина в тазобедренном сочленении, см |
24,9±0,6 |
22,7±0,6 |
2,2 |
0,028 |
|
Живая масса, кг |
39,9±0,9 |
37,3±0,7 |
2,6 |
0,039 |
Индексы телосложения телят, рассчитанные по результатам бонитировки 2024 года
Table 5
Physique indices of calves calculated by the quality indicators of 2024
При воспроизводстве бисексуальных видов генетическое разнообразие популяции определяется полом, представленным с меньшей численностью и эффективной величиной популяции. В оленеводстве отношение половозрелых самцов к самкам колеблется в широких пределах в зависимости от внешних условий, направления и сложившихся традиций ведения отрасли. В нашем исследовании численность генотипированных самцов и самок составила соответственно 49 и 253 животных или 165 эффективных особей, что в три раза больше минимального эффективного размера популяции, необходимого для снижения вероятности нарастания инбридинга и дрейфа генов.
Исходя из сказанного выше, нами с использованием микросателлитных маркеров была проведена сравнительная оценка генетического разнообразия северных оленей ЯОС в зависимости от половой принадлежности. В результате исследования установили (табл. 6), что по локусу NVHRT76 между самцами и самками наблюдается достоверное (р ≤ 0,05) несоответствие частот аллеля NVHRT7697. Этот аллель выявлен у 13,4 % важенок, тогда как у хоров его частота составляла 3,1 %. По локусу аллель RT30188 встречался в 2,5 раза чаще у хоров (р ≤ 0,001), тогда как аллель RT30202, наоборот, в 8,4 раз реже (р ≤ 0,001). Особо стоит отметить микросателлиту RT30190, частота распространения которого у важенок составила 0,320, а у самцов он не был обнаружен. Аллель NV21170 встречался у хоров с частотой 0,02, у самок – 0,215 (р ≤ 0,001). Локус RT9 характеризовался большей распространенностью среди важенок тандемных повторов RT9120 и RT9128, тогда как среди самцов наиболее часто встречался RT9126 (0,969 против 0,628; р ≤ 0,001). По локусу RT27 достоверная разница (р ≤ 0,05) установлена в отношении микросателлита RT27133, который не был выявлен у хоров, а среди важенок встречался с частотой 0,049.
В табл. 7 показаны ДНК-микросателлиты, выявленные среди важенок и не обнаруженные среди самцов. Уста-
Таблица 6
Характеристика частот ДНК-микросателлитов у северных оленей ненецкой породы в зависимости от пола
Table 6
Characterisation of DNA microsatellite frequencies in reindeer of the Nenets breed depending on sex
|
Аллель |
q |
Аллель |
q |
Аллель |
q |
|||
|
Хоры |
Важенки |
Хоры |
Важенки |
Хоры |
Важенки |
|||
|
NVHRT76 |
RT30 |
NV21 |
||||||
|
91 |
0,000 |
0,006 |
188 |
0,663 |
0,267*** |
158 |
0,051 |
0,022 |
|
95 |
0,000 |
0,022 |
190 |
0,000 |
0,320*** |
160 |
0,122 |
0,128 |
|
97 |
0,031 |
0,134* |
192 |
0,020 |
0,008 |
162 |
0,051 |
0,016 |
|
99 |
0,000 |
0,006 |
194 |
0,000 |
0,010 |
164 |
0,082 |
0,049 |
|
101 |
0,000 |
0,006 |
198 |
0,041 |
0,014 |
166 |
0,306 |
0,211 |
|
103 |
0,000 |
0,002 |
200 |
0,204 |
0,144 |
168 |
0,265 |
0,251 |
|
105 |
0,194 |
0,099 |
202 |
0,020 |
0,168*** |
170 |
0,020 |
0,215*** |
|
107 |
0,510 |
0,375 |
204 |
0,000 |
0,004 |
172 |
0,102 |
0,103 |
|
109 |
0,255 |
0,294 |
208 |
0,010 |
0,020 |
174 |
0,000 |
0,004 |
|
111 |
0,010 |
0,055 |
210 |
0,041 |
0,040 |
RT7 |
||
|
RT6 |
212 |
0,000 |
0,006 |
116 |
0,000 |
0,002 |
||
|
101 |
0,010 |
0,022 |
NVHRT24 |
216 |
0,031 |
0,024 |
||
|
103 |
0,082 |
0,103 |
147 |
0,000 |
0,004 |
218 |
0,020 |
0,069 |
|
105 |
0,276 |
0,235 |
149 |
0,041 |
0,069 |
220 |
0,500 |
0,385 |
|
107 |
0,031 |
0,042 |
151 |
0,276 |
0,209 |
222 |
0,082 |
0,146 |
|
109 |
0,071 |
0,043 |
153 |
0,500 |
0,496 |
224 |
0,031 |
0,053 |
|
111 |
0,051 |
0,061 |
155 |
0,000 |
0,016 |
226 |
0,020 |
0,016 |
|
113 |
0,051 |
0,059 |
157 |
0,184 |
0,206 |
228 |
0,153 |
0,136 |
|
115 |
0,388 |
0,397 |
RT1 |
230 |
0,153 |
0,132 |
||
|
117 |
0,041 |
0,036 |
223 |
0,082 |
0,103 |
232 |
0,010 |
0,032 |
|
119 |
0,000 |
0,002 |
225 |
0,163 |
0,119 |
234 |
0,000 |
0,004 |
|
RT9 |
227 |
0,163 |
0,111 |
RT27 |
||||
|
116 |
0,000 |
0,020 |
229 |
0,122 |
0,166 |
133 |
0,000 |
0,049* |
|
118 |
0,010 |
0,069 |
231 |
0,010 |
0,026 |
135 |
0,367 |
0,403 |
|
120 |
0,010 |
0,071* |
233 |
0,000 |
0,008 |
137 |
0,000 |
0,006 |
|
122 |
0,010 |
0,059 |
235 |
0,153 |
0,134 |
139 |
0,296 |
0,247 |
|
124 |
0,000 |
0,020 |
237 |
0,051 |
0,105 |
141 |
0,010 |
0,034 |
|
126 |
0,969 |
0,628*** |
239 |
0,092 |
0,091 |
143 |
0,000 |
0,004 |
|
128 |
0,000 |
0,085*** |
241 |
0,143 |
0,113 |
145 |
0,102 |
0,059 |
|
130 |
0,000 |
0,026 |
243 |
0,010 |
0,004 |
147 |
0,143 |
0,125 |
|
132 |
0,000 |
0,022 |
245 |
0,010 |
0,022 |
149 |
0,082 |
0,073 |
Примечание. Различия достоверны: *р ≤ 0,05; ***р ≤ 0,001 по отношению к хорам.
Note. Differences are reliable at * р ≤ 0.05; *** р ≤ 0.001 for males.
новлено, что в общей сложности среди исследуемого массива животных выделено 24 коротких тандемных повтора, которые были характерны только для самок. Приват-ал-лели присутствовали по всем девяти анализируемым локусам со средней частотой 0,073. Максимальное различие сумм частот наблюдалось по RT30 (0,340) и RT9 (0,173), а минимальное – по RT6 (0,002).
Полокусный анализ соответствия наблюдаемых и ожидаемых генотипов, согласно закону Харди-Вайнберга, показал, что наибольшее сходство фактического и теоретического распределения генотипов у обоих полов наблюдалось по локусу NVHRT24, максимальное расхождение у хоров – по RT6, а у важенок – по RT30. Несоответствие частот генов у самцов и самок является одной из
Таблица 7
Приват-аллели по девяти анализируемым микросателлитным локусам, встречающиеся среди важенок и не обнаруженные у хоров
Table 7
Private alleles for nine analysed microsatellite loci found among reindeer females and not found among males
В табл. 8 дана характеристика основных генетических параметров анализируемой популяции северных оленей. Установлено, что наименьшей полиморфностью среди важенок характеризуется локус NVHRT24 (шесть вариантов микросателлитов), а среди хоров NVHRT24 и RT9 (четыре варианта микросателлитов). Наиболее полиморфным яв-
Таблица 8
Полокусная характеристика основных генетических показателей северных оленей ненецкой породы в зависимости от половой принадлежности
Table 8
Per locus characterisation of main genetic parameters of reindeer of the Nenets breed depending on sex affiliation
|
Локус |
Пол |
N A |
NA q ≥ 10% |
N E |
NA/ N E |
I |
H O |
H E |
FIS |
|
NVHRT76 |
Хоры |
7 |
3 |
2,75 |
2,55 |
1,163 |
0,286 |
0,636 |
0,550 |
|
Важенки |
10 |
3 |
3,86 |
2,59 |
1,573 |
0,462 |
0,741 |
0,377 |
|
|
RT30 |
Хоры |
8 |
2 |
2,06 |
3,88 |
1,063 |
0,551 |
0,514 |
-0,072 |
|
Важенки |
11 |
3 |
4,44 |
2,48 |
1,697 |
0,565 |
0,775 |
0,271 |
|
|
NV21 |
Хоры |
8 |
4 |
4,96 |
1,61 |
1,792 |
0,245 |
0,798 |
0,693 |
|
Важенки |
9 |
5 |
5,42 |
1,66 |
1,823 |
0,399 |
0,816 |
0,511 |
|
|
RT6 |
Хоры |
9 |
2 |
4,07 |
2,21 |
1,703 |
0,959 |
0,754 |
-0,272 |
|
Важенки |
10 |
3 |
4,23 |
2,36 |
1,762 |
0,779 |
0,764 |
-0,020 |
|
|
NVHRT24 |
Хоры |
4 |
3 |
2,77 |
1,44 |
1,144 |
0,714 |
0,639 |
-0,117 |
|
Важенки |
6 |
3 |
2,97 |
2,02 |
1,273 |
0,605 |
0,663 |
0,087 |
|
|
RT7 |
Хоры |
9 |
3 |
3,26 |
2,76 |
1,545 |
0,449 |
0,694 |
0,353 |
|
Важенки |
11 |
4 |
4,64 |
2,37 |
1,827 |
0,688 |
0,785 |
0,124 |
|
|
RT9 |
Хоры |
4 |
1 |
1,06 |
3,77 |
0,170 |
0,061 |
0,060 |
-0,017 |
|
Важенки |
9 |
1 |
2,40 |
3,75 |
1,374 |
0,352 |
0,583 |
0,396 |
|
|
RT1 |
Хоры |
11 |
5 |
7,68 |
1,43 |
2,130 |
0,816 |
0,870 |
0,062 |
|
Важенки |
12 |
7 |
8,65 |
1,39 |
2,236 |
0,838 |
0,884 |
0,052 |
|
|
RT27 |
Хоры |
6 |
4 |
3,84 |
1,56 |
1,490 |
0,673 |
0,740 |
0,091 |
|
Важенки |
9 |
3 |
3,98 |
2,26 |
1,645 |
0,419 |
0,748 |
0,440 |
|
|
M±m |
Хоры |
7,33± 0,78 |
3,00± 0,41 |
3,61± 0,64 |
2,36± 0,32 |
1,356± 0,188 |
0,528± 0,018 |
0,634± 0,080 |
0,141± 0,108 |
|
Важенки |
9,57± 0,58* |
3,56± 0,56 |
4,51± 0,60 |
2,32± 0,22 |
1,690± 0,093 |
0,567± 0,010 |
0,751± 0,029 |
0,249 ±0,064 |
|
Примечание. Различия достоверны: *р ≤ 0,05 по отношению к хорам.
Note. Differences are reliable at * р ≤ 0.05 for males.
ляется локус RT1, по которому у самцов выявлено 11 аллелей, а у самок – 12. Среднее количество вариантов микросателлитов у важенок было на 2,24 больше (8 ≤ 0,05), чем у хоров. Число эффективных аллелей у обоих полов было минимальным по локусу RT1 (хоры – 1,06; важенки – 2,40), а максимальные значения установлены по RT9 (хоры – 7,68; важенки – 8,65). Существенную разницу между числом наблюдаемых и эффективных аллелей у самцов установили по локусу RT30 (в 3,88 раза), у самок – RT9 (в 3,75 раза), а наибольшее соответствие у важенок и хоров по аллелям локуса RT1. Оценка индекса Шеннона показала, что у самцов и самок наиболее полиморфным являлся локус RT1 (2,130 у хоров и 2,236 у важенок). Среди важенок минимальное генетическое разнообразие установлено по локусу NVHRT24 (1,273), а среди хоров – по локусу RT9 (0,170).
Максимальной наблюдаемой гетерозиготностью среди самцов характеризовался локус RT6 (0,959), а минимальной – RT9 (0,061). Среди важенок наибольшая фактическая гетерозиготность установлена по локусу RT1 (0,838), а наименьшая – по RT9 (0,352). Оценка ожидаемой гетерозиготности показала, что локус RT9 также характеризуется минимальными значениями (хоры – 0,060; важенки – 0,583), а максимальные значения, независимо от половой принадлежности, установлены по локусу RT1 (хоры – 0,870; важенки – 0,884). Полокусный анализ уровня инбредно-сти показал, что наибольшим соответствием наблюдаемой и ожидаемой гетерозиготности среди хоров характеризуется локус RT9 (-0,017), среди важенок – RT6 (-0,020). Заметная редукция гетерозигот, независимо от пола животных, прослеживалась по локусу NV21 (хоры – 0,693; важенки – 0,511), тогда как выраженный избыток гетерозигот наблюдался у самцов по локусу RT6 (-0,272).
Анализ генетического сходства свидетельствует (табл. 9, рис. 1), что максимально близкой структурой между важенками и хорами характеризуется локус RT6 (DN=0,006), а наибольшим генетическим различием – локусы RT9 (DN=0,152) и RT30 (DN=0,299). При этом необходимо отметить, что снижение генетического сходства по указанным локусам произошло по причине сокращения генетического разнообразия у хоров. В целом, по частотам встречаемости аллелей девять проанализированных STR-локусов, величину генетической дивергенции между самцами и самками можно характеризовать как невысокую (DN=0,073).
Проведенный анализ микросателлитного профиля северных оленей ненецкой породы свидетельствует, что важенки характеризуются более высоким генетическим разнообразием по сравнению с хорами. Отчасти это, по-видимому, объясняется разными размерами (численностями) выборок, представляющих две половые группы. Однако с учетом биологии размножения северных оленей, а именно участия в спаривании на 8-12 самок одного самца, можно заключить, что
Table 9
Characterisation of genetic divergence for individual STR loci between reindeer females and males of the Nenets breed
|
Локус |
I N |
D N |
|
NVHRT76 |
0,938 |
0,064 |
|
RT9 |
0,859 |
0,152 |
|
NVHRT24 |
0,986 |
0,014 |
|
RT30 |
0,742 |
0,299 |
|
RT1 |
0,980 |
0,020 |
|
RT6 |
0,994 |
0,006 |
|
RT27 |
0,962 |
0,039 |
|
NV21 |
0,932 |
0,071 |
|
RT7 |
0,976 |
0,024 |
|
В среднем |
0,930 |
0,073 |
Характеристика генетической дивергенции по отдельным STR-локусам между важенками и хорами ненецкой породы северных оленей
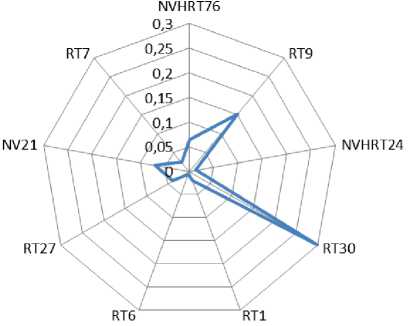
Рисунок 1. Визуализация генетической дистанции между важенками и хорами ненецкой породы северных оленей по отдельным микросател-литным локусам.
Figure 1. Visualisation of the genetic distance between females and males of the Nenets reindeer breed by individual microsatellite loci.
именно хоры будут оказывать существенное влияние на генетическую структуру популяции. Поэтому наиболее вероятным прогнозом, при отсутствии завоза производителей из других хозяйств, в данном стаде будет являться «генетическая эрозия», что в целом окажет негативное воздействие на генетическое разнообразие анализируемой популяции.
Численность и показатели вероятностей кластерной принадлежности особей
Table 10
Number and cluster affiliation likelihood of individuals
В результате деления на кластеры при k=5 (табл. 10), были сформированы группы с численностью от 16 до 25 животных, с минимальной вероятностью принадлежности к кластеру от 0,213 (k2) и максимальной – до 0,610 (k2). Средние значения Q находились в пределах 0,257–0,366.
Проведенная в программе GenAlEx генетическая оценка показала (табл. 11), что в исследованной выборке присутствует 119 аллелей, или в среднем по 7,438 на локус. Максимальное количество наблюдаемых микросателлитов пришлось на k5 (6,188), минимальное – на k3 (5,188). Число эффективных аллелей также было наибольшим в k5 (3,903), а наименьшим – в k1 (3,110). Одновременно первый кластер по значению индекса Шеннона характеризовался меньшим генетическим разнообразием (I=1,202), пятый, напротив, максимальным (I=1,414). В целом, уровень аллельного разнообразия исследуемой популяции составил 1,413. Наблюдаемая средняя гетерозиготность особей в k5 была выше на 0,083, чем в k1, а ожидаемая – на 0,074. Разница несмещенного показателя ожидаемой гетерозиготности между первым и пятым кластерами составила 0,051.
Оценка F-статистики показала, что по k1, k4 и k5 присутствовал незначительный избыток гетерозигот (-0,0610,029), по k2 и k3 установили соответствие наблюдаемой и ожидаемой гетерозиготности. В целом по Pop отмечена небольшая редукция гетерозигот.
Внутрипопуляционная генетическая дифференциация стада северных оленей Ямальской опытной станции
На следующем этапе исследовательской работы была проведена оценка внутрипопуляционной генетической дифференциации стада северных оленей ненецкой породы кластерным методом. Ана-
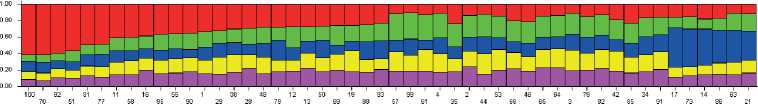
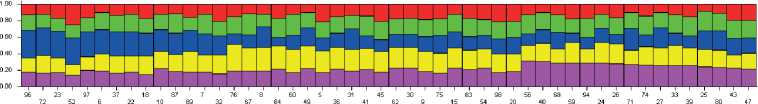
Рисунок 2. Визуализация генетической гетерогенности исследуемой выборки северных оленей, разделенной на пять кластеров (по оси Х – списочные номера животных, по оси Y – вероятности членства в кластерах: k1 – красный, k2 – зеленый, k3 – синий, k4 – желтый, k5 – сиреневый).
Figure 2. Visualisation of the genetic heterogeneity of the studied sample collection of reindeer divided into five clusters (X-axis – list numbers of animals, Y-axis – probabilities of cluster membership: k1 – red, k2 – green, k3 – blue, k4 – yellow, k5 – lilac).
Таблица 11 Характеристика популяции северных оленей, подразделенных на кластеры по основным генетическим показателям
Table 11
Characteristics of the reindeer population subdivided into clusters by the main genetic indicators
|
k |
Na |
Ne |
I |
Ho |
He |
uHe |
F is |
|
|
1 (n=25) |
X |
5,625 |
3,110 |
1,202 |
0,608 |
0,591 |
0,603 |
-0,029 |
|
Sx |
0,625 |
0,347 |
0,135 |
0,066 |
0,058 |
0,060 |
0,043 |
|
|
2 (n=19) |
X |
6,250 |
3,715 |
1,356 |
0,635 |
0,631 |
0,648 |
0,002 |
|
Sx |
0,574 |
0,474 |
0,142 |
0,066 |
0,060 |
0,062 |
0,033 |
|
|
3 (n=20) |
X |
5,188 |
3,436 |
1,269 |
0,638 |
0,632 |
0,648 |
0,001 |
|
Sx |
0,614 |
0,364 |
0,138 |
0,064 |
0,055 |
0,056 |
0,040 |
|
|
4 (n=20) |
X |
5,875 |
3,453 |
1,303 |
0,656 |
0,621 |
0,637 |
-0,061 |
|
Sx |
0,515 |
0,390 |
0,135 |
0,065 |
0,061 |
0,062 |
0,024 |
|
|
5 (n=16) |
X |
6,188 |
3,903 |
1,414 |
0,691 |
0,665 |
0,686 |
-0,032 |
|
Sx |
0,702 |
0,463 |
0,141 |
0,061 |
0,053 |
0,055 |
0,037 |
|
|
Pop (n=100) |
X |
7,438 |
3,827 |
1,413 |
0,642 |
0,651 |
0,654 |
0,017 |
|
Sx |
0,713 |
0,453 |
0,143 |
0,059 |
0,057 |
0,057 |
0,010 |
|
Таблица 12
Значимость (P) различий нарушения равновесия по Харди–Вайнбергу (HWE) для каждой комбинации «выборка-локус»
Table 12
Significance (P) of differences in Hardy-Weinberg equilibrium (HWE) disturbance for each sample-locus combination
|
Локус |
Кластеры |
Pop |
||||
|
1 |
2 |
3 |
4 |
5 |
||
|
BMS1788 |
0,976 |
0,741 |
0,733 |
0,080 |
0,271 |
0,327 |
|
RT30 |
0,141 |
0,948 |
0,438 |
0,844 |
1,000 |
0,999 |
|
RT1 |
0,982 |
0,360 |
0,902 |
0,065 |
0,390 |
0,517 |
|
RT9 |
0,918 |
0,462 |
0,515 |
0,194 |
0,711 |
0,587 |
|
C143 |
0,545 |
0,967 |
0,831 |
0,939 |
0,460 |
0,474 |
|
RT7 |
0,284 |
0,980 |
0,723 |
0,577 |
0,985 |
0,985 |
|
OHEQ |
0,828 |
0,969 |
0,954 |
0,974 |
0,957 |
0,979 |
|
FCB193 |
0,811 |
0,440 |
0,535 |
0,709 |
0,581 |
0,695 |
|
RT6 |
0,928 |
0,887 |
0,789 |
0,996 |
0,111 |
0,027 |
|
RT24 |
0,179 |
0,734 |
0,926 |
0,649 |
0,013 |
0,000 |
|
BMS745 |
0,379 |
0,142 |
0,128 |
0,987 |
0,262 |
0,311 |
|
NVHRT16 |
0,091 |
0,603 |
0,523 |
0,804 |
0,381 |
0,492 |
|
T40 |
0,652 |
0,578 |
0,876 |
0,771 |
0,650 |
0,598 |
|
C276 |
0,999 |
0,011 |
0,954 |
0,996 |
0,463 |
0,095 |
|
C217 |
0,853 |
0,934 |
0,274 |
0,285 |
0,019 |
0,023 |
|
C32 |
- |
0,906 |
- |
- |
0,920 |
0,916 |
Полокусный анализ соответствия наблюдаемого распределения частот генотипов ожидаемому по закону Харди-Вайнберга показал (табл. 12), что за исключением локуса C276 k2 (р ≤ 0,05), по всем кластерам отсутствуют значимые отклонения. В целом в Pop установили статистически значимое отклонение наблюдаемого распределения генотипов от ожидаемого по локусам RT6 (р ≤ 0,05), RT24 (р ≤ 0,001) и C217 (р ≤ 0,05).
Анализ специфичных для каждого кластера аллелей показал (табл. 13), что по k1 и k2 выявлено по два при-ват-аллеля: по локусам Rt6 и C276 с суммарной частотой 0,060; Rt7240 и FCB193138 с общей частотой 0,079 соответственно. Встречаемость трех приват-аллелей в k4 соста- вила 0,100, которые находились в локусах Rt1, BMS745, NVHRT16. Пятый кластер характеризовался максимальным значением частот специфичных микросателлитов (Σq=0,219), находящихся в пяти локусах: BMS1788, Rt30, Rt7, OheQ, Rt24. В отличие от других кластеров, k3 не имел приват-микросателлитов.
Оценка генетического сходства между сформированными группами показала, что минимальная генетическая дистанция, рассчитанная по Нею (табл. 14, рис. 3), установлена между k2 и k4 (0,043) и максимальная – между k2 и k3 (0,119). Вариация несмещенных оценок генетической дивергенции, по Нею, была ниже некорректированных, а между k2 и k4 вовсе отсутствовала. Попарные оценки генетических дистанций между группами (кластерами), рассчитанные разными методами, дали сходные результаты ранжирования величин генетического различия.
Сравнение аллельных характеристик между отдельными кластерами и популяцией в целом, с использованием различных статистических подходов, выявило некоторые различия. Так, стандартная генетическая дистанция Нея характеризовалась наибольшим значением при сравнении Pop с k3 (DN=0,049), а наименьшим – Pop
Таблица 13
Приват-аллели, выявленные у генотипированных особей, разделенных на кластеры
Table 13
Private alleles identified in genotyped individuals divided into clusters
|
Кластер |
Локусы |
Микросателлита |
q |
Σq |
|
1 |
Rt6 |
186 |
0,020 |
0,060 |
|
C276 |
259 |
0,040 |
||
|
2 |
Rt7 |
240 |
0,053 |
0,079 |
|
FCB193 |
138 |
0,026 |
||
|
4 |
Rt1 |
267 |
0,050 |
0,100 |
|
BMS745 |
330 |
0,025 |
||
|
NVHRT16 |
142 |
0,025 |
||
|
5 |
BMS1788 |
150 |
0,031 |
0,219 |
|
Rt30 |
223 |
0,031 |
||
|
Rt7 |
254 |
0,063 |
||
|
OheQ |
299 |
0,031 |
||
|
Rt24 |
248 |
0,063 |
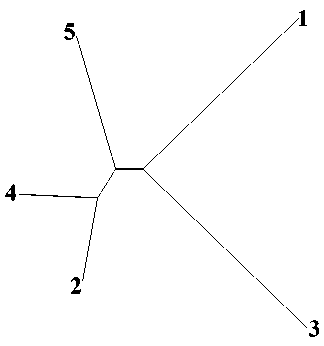
Рисунок 3. Визуализация генетических дистанций (DN) между группами, сформированными в кластерах.
Figure 3. Visualisation of genetic distances (DN) between groups formed in clusters.
Таблица 14 Результаты оценки генетических дистанций между группами (кластерами)
Table 14
Evaluation results of genetic distances between groups (clusters)
Таким образом, деление популяции с помощью программы Structure 2.3.4 на пять кластеров позволило с невысокой вероятностью членства в собственном кластере сформировать генетически дифференцированные группы особей, из которых наибольшим аллельным разнообразием обладали животные, отнесенные к k5, а наименьшим – к k1. Кластеризация дала возможность выявить приват-аллели, наибольшее количество которых было характерно для k5. Используя различные способы оценки генетического различия, наиболее дистанцированными по отношению к аллелофонду исследованной популяции можно считать k1 и k3. По стаду северных оленей ненецкой породы Ямальской опытной станции в целом наблюдали незначительную редукцию гетерозигот, минимизировать которую можно путем целенаправленного отбора и подбора особей в зависимости от их индивидуальных генотипов или принадлежности группы особей к кластеру, установленных по результатам кластерного анализа.
В целом проведенные исследования полиморфизма микросателлитов показали, что генотипированная выборка северных оленей ненецкой породы представляет генетически слабо дифференцированный массив.
Заключение
Изучение численности домашних северных оленей по федеральным округам показало, что Уральский федеральный округ занимает лидирующую позицию по количеству этих животных. В 2023 г. здесь наблюдалось снижение численности оленей на 17,58 % по сравнению с 2018 г. В рамках данного округа наибольшую долю составляют олени Ямало-Ненецкого автономного округа – 95,11 %, в то время как Ханты-Мансийский автономный округ владеет лишь 4,89 % от общего поголовья. Второе место по численности оленей занимают Дальний Восток и Северо-Западный федеральный округ. В Дальне-Восточном федеральном округе более половины оленей обитает в Республике Сахе (53,26 %), значительная доля приходится на Чукотский автономный округ (35,07 %), а оставшиеся 11,67 % находятся в Камчатском крае. В Северо-Западном федеральном округе большинство оленей сосредоточено в Ненецком автономном округе (НАО) (55,66 %), за Республикой Коми – 27,68 %, наименьшая доля наблюдается в Мурманской области (16,67 %). Наименьшее количество оленей зарегистрировано в Сибирском федеральном округе (Красноярский край).
Исследование выявило негативную динамику численности оленей в арктических регионах в 2023 г., за исключением НАО и Республики Сахи, где снижения не произошло. Максимальное сокращение отмечено в ЯНАО. В большинстве арктических регионов в 2023 г. оленеводство находится под контролем сельскохозяйственных организаций. Исключением является ЯНАО, где значительная часть оленей (63,09 %) принадлежит домохозяйствам, тогда как в Красноярском крае на К(Ф)Х и ИП приходится 51,59 %.
Анализ состава стада ЯОС показал, что в хозяйстве наблюдается нехватка самок. В частности, общее количество важенок и нетелей составляет лишь 47,6 %, что на 14,4% ниже рекомендованного уровня. Это несоответствие связано с увеличением численности быков-кастратов (на 3,7 %), третьяков (на 4,5) и самцов-телят (на 5,3 %). Такое смещение половой структуры в сторону мужского пола отрицательно влияет на количество производимого молодняка и увеличивает конкуренцию между производителями. В связи с этим, одним из способов улучшения селекционно-племенной работы в стаде является оптимизация его половозрастной структуры через плановую и своевременную выбраковку определенных особей.
Анализ фенотипических признаков исследуемой популяции оленей показал, что животные имеют типичное для уральского экотипа ненецкой породы телосложение – сравнительно невысокие с хорошо развитым в длину и глубину туловищем, несколько удлинённой грудью, хорошо развитым костяком, недлинной, сравнительно широкой головой. При оценке данных бонитировки за по- следние два года установлено, что телята, родившиеся в 2024 г., по некоторым показателям уступают признакам молодняка, появившегося в 2023 г. Так, масса телеваженок оказалась ниже на 8,7 кг (р≤0,001), а телехоров – на 5,6 кг (р≤0,001). Основной причиной такой заметной разницы, вероятно, послужили неблагоприятные климатические условия, наблюдаемые весной 2024 г.
Сравнительная оценка генетического разнообразия северных оленей ненецкой породы в зависимости от половой принадлежности показала, что среди важенок присутствуют 24 микросателлита, отсутствующие у хоров. Приват-аллели выявлены по всем девяти анализируемым локусам, средняя частота которых составила 0,073. Полокусный анализ соответствия наблюдаемых и ожидаемых генотипов показал, что наибольшее сходство, независимо от пола, наблюдается по локусу NVHRT24, а максимальное расхождение просматривалось по RT6 у хоров, и по RT30 – у важенок. Достоверное несоответствие распространенности наблюдаемых и ожидаемых генотипов у самцов было характерно для NVHRT76, RT27 и NV21 (р ≤ 0,001), а у самок – для всех девяти анализируемых локусов (р ≤ 0,05–0,001). Анализ основных генетических параметров свидетельствует, что среднее количество наблюдаемых вариантов микросателлитов среди важенок было на 2,24 больше (р ≤ 0,05) по сравнению с хорами. Наибольшим несоответствием наблюдаемых и эффективных микросателлитов у самцов характеризовался локус RT30 (разница – в 3,88 раз), у самок – RT9 (разница – в 3,75 раз). Полокусный анализ уровня инбредности показал, что наибольшим соответствием наблюдаемой и ожидаемой гетерозиготности у хоров характеризуется локус RT9 (-0,017), а у важенок – RT6 (-0,020). Заметный дефицит гетерозиготности, независимо от пола, прослеживался по NV21 (хоры – 0,693; важенки – 0,511), тогда как выраженный сдвиг в сторону избытка гетерозигот наблюдался лишь среди самцов по локусу RT6 (-0,272). Максимально близкой структурой между важенками и хорами характеризовался локус RT6 (DN=0,006), а наибольшим генетическим различием – локусы RT9 (DN=0,152) и RT30 (DN=0,299). Тем не менее средняя генетическая дивергенция между самцами и самками была незначительной (DN=0,073). Полученные данные прогнозируют дальнейшее сужение генетического разнообразия анализируемой популяции в случае отсутствия обмена хорами.
Кластерный анализ, проведенный в программе Structure 2.3.4. генотипированных по 16 микросателлит-ным маркерам животных, показал, что наивысший уровень дифференциации наблюдается при делении выборки на пять кластеров. При этом максимальное количество наблюдаемых и эффективных STR-аллелей установили в k5 (6,188 и 3,903 соответственно). По величине индекса Шеннона минимальным генетическим разнообразием (I=1,202) характеризовался первый кластер, пятый – напротив максимальным (I=1,414). По k1, k4 и k5 установлен незначительный избыток гетерозигот (-0,061-0,029), а в целом по популяции отмечалась их небольшая редукция (Fis=0,017). У особей k1 выявлены приват-аллели Rt6186 и C276259 с суммарной частотой 0,060, характерные для k2 микросателлиты Rt7240 и FCB193138 встречались с общей частотой 0,079, в k4 суммарная частота встречаемости приват-ал- лелей Rt1267, BMS745330, NVHRT16142 составила 0,100, в k5 приват-аллели BMS1788150, Rt30223, Rt7254, OheQ299, Rt24248 обнаружили у 21,9 % животных. Генетическая дистанция (DN) была минимальной – между k2 и k4 (0,043), максимальной между k2 и k3 (0,119). По отношению к популяции в целом максимальная генетическая дистанция прослеживалась у k3 (DN=0,049), а наименьшая – у k2 (DN=0,025). Самая высокая оценка по Dest-статистике получена при сравнении пар Pop-k1 и Pop-k3 (Dest=0,021) и отсутствовала между Pop-k2 и Pop-k5 (Dest=-0,004-0,002).
В целом, в анализируемой популяции северных оленей ненецкой породы было зафиксировано легкое уменьшение числа гетерозигот. Эта редукция может быть сокращена с помощью целенаправленного отбора и подбора животных на основе их индивидуальных генотипов или принадлежности к определенной группе, выделенной в результате кластерного анализа. Исследования, проведенные по полиморфизму микросателлитов, продемонстрировали, что отобранная для генотипирования выборка северных оленей ненецкой породы составляет генетически однородный массив с низкой дифференциацией.
Таким образом, по результатам исследований, проведенных в 2024 г., получены экспериментальные данные для создания технологий направленных селекционных процессов в северном оленеводстве.
Автор заявляет об отсутствии конфликта интересов.
Список литературы Оценка генетического потенциала оленей ненецкой породы с целью создания технологий направленных селекционных процессов в северном оленеводстве
- Забродин, В. А. Результаты изучения инфекционных и инвазионных заболеваний в Таймырской популяции диких северных оленей / В. А. Забродин, К. А. Лайшев, В. П. Печин // Северный олень в России 1982-2002. - М.: Триада-Фарм, 2003. - С. 261-272. EDN: WMXQKV
- Южаков, А. А. Новые знания, методы и модели в разведении, экологии и эпизоотологии северных оленей / А. А. Южаков, Т. М. Романенко, К. А. Лайшев. - СПб. - Пушкин, 2018. EDN: UXXMIY
- Романенко, Т. М. Экстерьерные особенности северных оленей острова Колгуев и Малоземельской тундры в сравнительном аспекте / Т. М. Романенко, А. С. Митюков // Известия Санкт-Петербургского государственного аграрного университета. - 2011. - № 24. - С. 113-118. EDN: PGFVRF
- Романенко, Т. М. Краниологическая характеристика северных оленей (Rangifer tarandus) ненецкой породы Тиманской и Большеземельской тундр Ненецкого АО / Т. М. Романенко, Ю. П. Вылко // Международный журнал прикладных и фундаментальных исследований. - 2019. - № 12. - С. 110-114. EDN: YAXGZK
- Андреев, В. Н. Северные олени / В. Н. Андреев, И. М. Голосов, Б. В. Преображенский. - Красноярск: Кн. изд-во, 1972. - 190 с.
