Оценка и отбор исходного материала для селекции солеустойчивых сортов риса в Республике Казахстан

Автор: Батаева Д.С., Усенбеков Б.Н., Рысбекова А.Б., Мухина Ж.М., Казкеев Д.Т., Жанбырбаев Е.А., Сартбаева И.А., Гаркуша С.В., Волкова С.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Зерновые: генетический потенциал, селекционное улучшение, агробиотехнологии
Статья в выпуске: 3 т.52, 2017 года.
Бесплатный доступ
Около 25 % всей поверхности суши представлены засоленными почвами, а в отдельных регионах Средней Азии, Закавказья, Украины и Казахстана - до 90 % всей орошаемой площади. Область рисосеяния Республики Казахстан также находится в зоне с повышенной засоленностью. Одна из проблем рисоводства Казахстана - ухудшения гумусного и мелиоративного состояния почв, усиление их вторичного засоления и деградации. В Кызылординской области загрязнение солевыми остатками поверхностных (до 3-5 г/л) и подземных (до 6-7 г/л) вод доходит до критической отметки. При этом по химическому составу преобладает сульфатно-хлоридно-нат-риевый тип засоления, который особо токсичен для сельскохозяйственных культур. Акдалинский и Каратальский массивы орошения в Алматинской области тоже находятся в пределах провинций сульфатно-содового и борного биогеохимического засоления почв. По данным Казахского НИИ рисоводства, в последние годы резко снизилась урожайность риса (с 50 до 35-48 ц/га) и качество производимой продукции (выход крупы - с 65 до 45-50 %). В этой связи селекция риса на солеустойчивость - важнейшая задача обеспечения продовольственной безопасности республики. Целью нашего исследования были оценка и отбор перспективного исходного материала для селекции риса на устойчивость к разным типам засоления. Объектом исследований служили сорта и сортообразцы (включая потомства гибридных комбинаций разных поколений) риса ( Oryza sativa L.) российской, казахстанской и филиппинской селекции (всего 34 генотипа). Для первоначальной оценки на стадии проростков был проведен лабораторный скрининг устойчивости к различным типам засоления: хлоридному, сульфатному и карбонатному. Солевой стресс оказывал негативное влияние на прирост общей биомассы проростков у исследованных образцов. Выявлено, что наиболее токсичен для растений риса карбонатный тип засоления; меньший эффект оказывали хлоридное и сульфатное. Гибрид F2 сорт Ханкайский 429 × коллекционный образец 4-09, а также сорта Маржан и Мадина накапливали наибольшую (в процентном соотношении) биомассу при всех трех типах засоления. Следовательно, указанные генотипы представляют ценность как исходный материал в схемах селекции на солеустойчивость. Молекулярный скрининг хромосомных участков ДНК, сцепленных с QTL солеустойчивости ( Saltol QTL), проводили с помощью ПЦР-анализа. Применяли микросателлитные маркеры RM 493 и AP 3206, тесно сцепленные с целевыми хромосомными регионами. Маркер RM 493 оказался наиболее информативным для ранжирования генетической плазмы риса по признаку солеустойчивости, выявляя полиморфизм между генотипами, контрастными по изучаемому признаку.
Хлоридное засоление, сульфатное засоление, карбонатное засоление, молекулярно-генетический скрининг, солеустойчивость, селекционные образцы
Короткий адрес: https://sciup.org/142214050
IDR: 142214050 | УДК: 633.18:631.522/.524:575.2:58.051(574) | DOI: 10.15389/agrobiology.2017.3.544ru
Estimation and selection of parental forms for breeding Kazakhstan salt tolerant rice varieties
About 25 % of the entire land surface is represented by saline soils, and up to 90 % of the total irrigated area - in some regions of Central Asia and the Caucasus, Ukraine and Kazakhstan. Rice-growing region of the Republic of Kazakhstan is also located in the area with high salinity. A challenge for rice growing in Kazakhstan is deteriorating humus and reclamation of soil, rise in soil secondary salinity and degradation. In Kyzylorda region, pollution of surface (up to 3-5 g/l) and ground water (up to 6-7 g/l) by salt residues reaches a critical point. The dominating sulfate-chloride-sodium type of salinity is especially toxic for crops. Akdalinski and Karatal zones of irrigation in the Almaty region are also located within the provinces of sulfate-soda and boric biogeochemical soil salinity. According the data of Kazakh Rice Research Institute, in recent years the rice yields and yield quality have sharply decreased - from 50 to 35-48 centners per hectare, and from 65 to 45-50 % of the groats output, respectively. In this regard, the rice breeding for salt tolerance is the most important to ensure food security in Kazakhstan. The objective of our study was the evaluation and selection a promising starting material of rice resistant to different types of salinity. To this end, we studied the rice ( Oryza sativa L.) varieties, collection samples and their hybrid combinations of different generations derived from Russia, Kazakhstan and Philippine (34 genotypes in total). For the initial assessment at the seedling stage we used the laboratory screening for tolerance to various types of salinity, i.e. chloride, sulphate and carbonate. Saline stress negatively affected seedling growth and total weight in the studied samples. It was revealed that the carbonate type of salinity is the most toxic for rice plants; the chloride and sulfate types were less adverse. The hybrid collection sample F2 Khankai 429 × 4-09 and as well as varieties Marzhan and Madina accumulated the highest percentage of biomass compared to the control at the salinity of all three types. Therefore, these genotypes are valuable in selection for salt tolerance. Molecular screening of chromosomal DNA regions linked to quantitative trait loci of salt tolerance ( Saltol QTL) by PCR with microsatellite markers closely linked to the target chromosomal regions, RM 493 and AP 3206, showed RM 493 to produce polymorphism which allowed to distinguish the studied genotypes contrasting in salt tolerance. Thus RM 493 is informative to rank the rice genetic plasma by salt tolerance
Текст научной статьи Оценка и отбор исходного материала для селекции солеустойчивых сортов риса в Республике Казахстан
Около 950 млн га земель сельскохозяйственного назначения в мире засолены и 77 млн га орошаются соленой водой. По прогнозу, к 2050 году засоление затронет более 50 % возделываемых территорий (1). В России засоленные почвы занимают 15 млн га, в том числе в Краснодарском крае — 196,4 тыс га, из них 76,4 тыс га находятся на орошаемых участках, причем более 85 % — в рисовых оросительных системах.
Засоление почвы выше 0,35 % приводит к гибели растений риса. Одна из серьезных проблем рисоводства в Республике Казахстан — ухудшение гумусного и мелиоративного состояния почв, усиление их вторичного засоления и деградация. Площадь засоленных и солонцовых почв в настоящее время составляет 111,6 млн га, или 41,0 % от всей территории республики. В Кызылординской области загрязнение солевыми остатками поверхностных (до 3-5 г/л) и подземных (до 6-7 г/л) вод доходит до критической отметки. На большей части (30 %) орошаемых рисосеющих территорий минерализация грунтовых вод достигает 5-10 г/л. Как следствие, урожайность культур здесь падает на 20-50 % (2). При этом по химиче-544
скому составу преобладает сульфатно-хлоридно-натриевый тип засоления, который особо токсичен для сельскохозяйственных растений. Акдалин-ский и Каратальский массивы орошения другого важного рисосеющего региона Казахстана — Алматинской области тоже находятся в пределах провинций сульфатно-содового и борного биогеохимического засоления почв. Избыток водорастворимых солей в почве вызывает изреживание всходов, тормозит рост и развитие растений. По данным Казахского НИИ рисоводства, в последние годы это стало причиной резкого снижения урожайности риса (с 50 до 35-48 ц/га) и качества производимой продукции (выход крупы уменьшился с 65 до 45-50 %) (3).
Основные возделываемые сорта риса в Казахстане уже не способны соответствовать возросшим запросам производства перерабатывающей и пищевой промышленности в связи с расширением площадей вторично засоленных почв. Снижение урожайности на засоленных почвах можно преодолеть посредством их рекультивации и повышения толерантности риса к засолению. Селекция риса для основных рисосеющих регионов Республики Казахстан — Кызылординской и Алматинской областей должна быть направлена на создание сортов, устойчивых к засолению, болезням и вредителям, с высокой продуктивностью и качеством крупы. Внедрение солеустойчивых сортов в производственную практику — экономически эффективный прием и основная задача современного рисосеяния в Казахстане.
Способность растений выживать и осуществлять полный цикл развития в среде с неблагоприятными для большинства других видов концентрациями солей принято называть солеустойчивостью. По Б.П. Строгонову, следует различать биологическую и агрономическую солеустойчивость (4, 5). Солеустойчивость — полигенный признак, который проявляется в комплексе разнообразных физиологических реакций, затрагивающих практически весь метаболизм растения и реализующихся на разных уровнях организации (6). Известны прямые и косвенные методы определения со-леустойчивости растений, предусматривающие оценку урожайности и продуктивности, скорости прорастания семян, интенсивности плазмолиза клеток, скорости выцветания хлорофилла в листьях проростков, помещенных в солевые растворы (7).
Солеустойчивость целесообразно оценивать по агрономической устойчивости, то есть по способности растений сохранить урожайность, определенную на незасоленном фоне, при выращивании на засоленной почве. Устойчивость к засолению почвы изменяется по фазам вегетации; при этом в онтогенезе у растений риса отмечены две критические фазы: в период прорастания—образования всходов и в фазу цветения (8-11).
В настоящей работе мы впервые провели скрининг казахстанских линий риса на солеустойчивость в раннюю фазу вегетации и молекулярнобиологический анализ с использованием микросателлитных маркеров.
Цель нашего исследования заключалась в оценке и отборе перспективного исходного материала для селекции риса на устойчивость к разным типам засоления.
Методика. Объектом исследований служили сорта и сортообразцы (включая потомства гибридных комбинаций разных поколений) риса (Ory-za sativa L.): российской селекции — Лиман, Регул, Дарий 23, Соната, ВНИИР10173, Славянец, Ханкайский 429, Фишт, Янтарь, Рапан, Кубань 3, Атлант, Серпантин, Курчанка и коллекционные образцы 4-09 и 49-09; казахстанской селекции — сорта Мадина, Аналог II, Баканасский, Маржан, коллекционный образце 34-09, гибридные линии F2 Соната ½ Лиман, F2 Регул ½ Курчанка, F2 Дарий 23 ½ коллекционный образец 49-09, F2 Хан- кайский 429 ½ коллекционный образец 4-09, F2 Маржан ½ Курчанка, F2 Баканасский ½ Аналог II, F2 Дарий 23 ½ Аналог II, F2 Кубань 3 ½ коллекционный образец 34-09, F2 Ханкайский 429 ½ Курчанка; филиппинской селекции — сорта FL 478 HB 9093, BINA dhan 8 HB 9106, BRRI dhan 47 HB 9114. Во всех экспериментах в качестве положительного контроля использовали сорта Маржан (казахстанский солеустойчивый стандарт), Курчанка (российский солеустойчивый стандарт), FL 478 НВ9093, BINA dhan 8 НВ9106, BRRI dhan 47 НВ9114 (солеустойчивые образцы, полученные из International Rice Research Institute, IRRI, Филиппины).
Фенологические наблюдения проводили по П.С. Ерыгину (12).
В лабораторных условиях осуществляли скрининг сортов, сортооб-разцов и гибридных линий на солеустойчивость в фазу прорастания по методике Всероссийского НИИ риса (13). Для этого семена проращивали в термостате при температуре 29 °С в течение 4 сут в условиях солевого стресса (0,75 % NaCl, 0,75 % Na2SO4, 0,075 % Na2CO3) (в 3-кратной повторности). На 5-е сут отбирали 20 типичных проростков из каждого варианта и определяли их массу с точностью до 0,01 г.
Молекулярный скрининг хромосомных регионов ДНК, сцепленных с QTL (quantitative trait loci) солеустойчивости ( Saltol QTL), проводили с помощью ПЦР-анализа в гибридных комбинациях F3 и F4. Экстракцию ДНК осуществляли СТАВ-методом, используя бесхлорофилльные 7-суточные проростки (14). Применяли микросателлитные маркеры, тесно сцепленные с целевыми хромосомными регионами, — RM 493 и AP 3206 (15). ПЦР поводили с 40-50 нг ДНК в конечном объеме 25 мкл. Состав реакционной смеси: 0,05 мМ dNTPs, по 0,3 мМ каждого праймера, 25 мM KCl, 60 мM Трис-HCl (pH 8,5), 0,1 % Тритон Х-100, 10 мМ 2-меркапто-этанол, 1,5 мМ MgCl2, 1 ед. Taq-полимеразы («New England Biolabs, Inc.», США). ДНК-амплификацию осуществляли в следующем режиме: начальная денатурация 4 мин при 94 °С; денатурация 1 мин при 94 °С, отжиг праймеров 1 мин при 55 °С, элонгация 1 мин при 72 °С (30 циклов); последний цикл синтеза 5 мин при 72 °С. Праймерные пары для указанных маркеров были синтезированы компанией ЗАО «Синтол» (г. Москва). Нуклеотидные последовательности праймерных пар для RM493 — TAGC-TCCAACAGGATCGACC (5'^3'), GTACG-TAAACGCGGAAGGTG (5'^3'); для AP3206 — GGAGGAGGAGAGGA-AGAAG (5'^3'), GCAAGAATTAAT-CCATGTGAAAGA (5'^3'). ПЦР проводили в ДНК-амплификаторе Терцик (НПО «ДНК-Технология», Россия). Для электрофоретического разделения продуктов ПЦР использовали 8 % полиакриламидный гель на основе 1½ Трис-боратного буфера (Трис-HCl — 0,09 М, борная кислота — 0,09 М, EDTA — 2 мМ, рН 8,2). Визуализацию проводили в УФ-свете после окрашивания гелей бромистым этидием.
Полученные данные обрабатывали с использованием компьютерной программы Microsoft Excel. В таблице представлены средние арифметические ( M ) и ошибки средней ( m ).
Результаты. Солевой стресс негативно влиял на прирост общей биомассы проростков у исследованных образцов (табл.). Наибольшим (в процентном отношении к контролю) накоплением биомассы в условиях хлоридного засоления характеризовались сорта казахстанской селекции Баканасский и Маржан; средними значениями — сорта российской селекции Рапан, Янтарь, Фишт, Соната и Ханкайский 429. Наименьшее накопление общей биомассы отмечали у сортов из IRRI: BINA dhan 8HB9106, FL 478 HB9093, BRRI dhan 47 HB9114, хотя они считаются стандартами солеустойчивости. Возможно, солеустойчивость у этих образцов проявля-546
ется на более поздних этапах онтогенеза.
Скрининг образцов риса (Oryza sativa L.) по общей биомассе проростков относительно контроля (%) при разных типах засоления (лабораторный опыт, M±m)
|
Сорт, образец, гибрид |
Засоление |
||
|
хлоридное (0,75 %) |
сульфатное (0,75 %) |
карбонатное (0,075 %) |
|
|
BINA dhan 8 HB9106 |
45,45±2,73 |
87,61±16,15 |
61,50±10,45 |
|
BRRI dhan 47 HB9114 |
46,19±4,89 |
87,01±14,19 |
74,43±8,15 |
|
FL 478 HB9093 |
47,75±4,86 |
79,47±5,77 |
61,97±6,66 |
|
Акдала |
74,98±8,25 |
66,31±6,22 |
59,65±6,46 |
|
Аналог II |
73,37±6,01 |
97,02±19,46 |
61,78±8,39 |
|
Атлант |
75,26±4,91 |
75,36±11,62 |
66,90±10,71 |
|
Баканасский |
81,97±16,42 |
65,39±8,75 |
75,30±15,28 |
|
ВНИИР10173 |
73,24±9,69 |
69,71±8,85 |
63,00±32,50 |
|
Дарий 23 |
71,72±12,26 |
82,33±13,51 |
62,10±5,61 |
|
Коллекционный образец 34-09 |
75,86±6,33 |
67,25±7,94 |
73,68±5,76 |
|
Коллекционный образец 4-09 |
61,93±4,31 |
87,47±22,14 |
54,80±9,58 |
|
Коллекционный образец 49-09 |
70,50±3,09 |
71,11±5,56 |
74,31±6,07 |
|
Кубань 3 |
79,29±6,50 |
82,67±14,44 |
68,86±9,06 |
|
Курчанка |
69,79±4,59 |
74,02±12,02 |
67,16±4,46 |
|
Лиман |
65,03±8,19 |
70,50±7,49 |
45,11±5,37 |
|
Мадина |
85,64±5,02 |
78,41±10,95 |
74,29±7,11 |
|
Маржан |
80,90±15,55 |
78,90±7,37 |
76,04±6,65 |
|
Рапан |
71,15±14,91 |
69,10±20,49 |
73,03±17,24 |
|
Регул |
70,36±12,61 |
98,36±10,36 |
75,15±13,77 |
|
Серпантин |
81,42±5,14 |
79,67±8,15 |
60,15±7,11 |
|
Славянец |
80,77±11,44 |
65,10±3,52 |
58,44±8,88 |
|
Соната |
70,23±6,56 |
79,50±7,02 |
55,62±3,15 |
|
Фишт |
77,40±8,23 |
83,97±8,76 |
72,83±3,84 |
|
Ханкайский 429 |
66,13±6,17 |
75,39±4,12 |
58,25±6,59 |
|
Янтарь |
72,97±12,66 |
78,07±16,28 |
57,29±4,06 |
|
F2 Баканасский ½ Аналог II |
78,01±8,05 |
71,90±6,78 |
70,23±10,10 |
|
F2 Дарий ½ Аналог II |
73,08±9,68 |
74,00±6,78 |
62,84±3,97 |
|
F2 Дарий 23 ½ коллекционный образец 49-09 |
67,34±6,05 |
81,03±8,43 |
77,06±7,07 |
|
F2 Кубань 3 ½ коллекционный образец 34-09 |
89,48±7,44 |
71,73±7,04 |
72,26±4,63 |
|
F2 Маржан ½ Курчанка |
68,37±11,37 |
71,37±6,41 |
67,76±10,91 |
|
F2 Регул ½ Курчанка |
70,63±7,46 |
58,79±5,36 |
69,03±4,33 |
|
F2 Соната ½ Лиман |
69,78±7,75 |
77,56±6,05 |
69,64±8,21 |
|
F2 Ханкайский 429 ½ Курчанка |
80,56±10,36 |
72,60±6,25 |
67,88±7,63 |
|
F2 Ханкайский 429 ½ коллекционный образец 4-09 |
70,71±6,69 |
96,74±31,91 |
80,04±4,73 |
По устойчивости к сульфатному засолению испытанные генотипы были разделены на три группы: высокоустойчивые (76-100 %), среднеустойчивые (70-75 %) слабоустойчивые (0-69 %). Высокую устойчивость проявили сорта казахстанской селекции Маржан (78 %), Мадина (77 %); сорта из IRRI FL 478 HB 9093 (79 %), BRRI dhan 47 HB 9114 (85 %), BINA dhan 8 HB 9106 (87 %); сорта из России Кубань 3 (82 %), Фишт (82 %), Соната (80 %), Атлант (93 %), Регул (97 %), Дарий 23 ½ коллекционный образец 49-09 (80 %), Ханкайский 429 ½ коллекционный образец 4-09 (91 %). Слабоустойчивыми к этому селективному фактору оказались проростки сортов и гибридов Баканасский, Рапан, Славянец, Акдала, ВНИ-ИР10173, F2 Регул ½ Курчанка, коллекционного образца 34-09.
Содовое засоление отличается более значительным повреждающим влиянием на поглотительную способность корней риса по сравнению с хлоридным и сульфатным (16). В наших экспериментах самую высокую чувствительность к карбонатному засолению проявили сорта селекции ВНИИ риса — Лиман, Соната, Янтарь, Славянец, Ханкайский 429, ВНИИР10173, Серпантин. Наибольшим накоплением биомассы характеризовались Маржан, Регул, Мадина, F2 Дарий 23 ½ коллекционный образец 49-09 и F2 Ханкайский 429 ½ коллекционный образец 4-09.
Имеются различные теории, объясняющие угнетение растений в условиях засоления. Согласно одной из них, это явление обусловлено ос- мотическим влиянием растворов солей, в соответствии с другой — подавление роста и развития растений происходит вследствие токсического воздействия поглощенных ионов на физиолого-биохимические процессы (17). В наших опытах наиболее токсичным было карбонатное засоление, меньший эффект оказывали хлоридное и сульфатное.
А
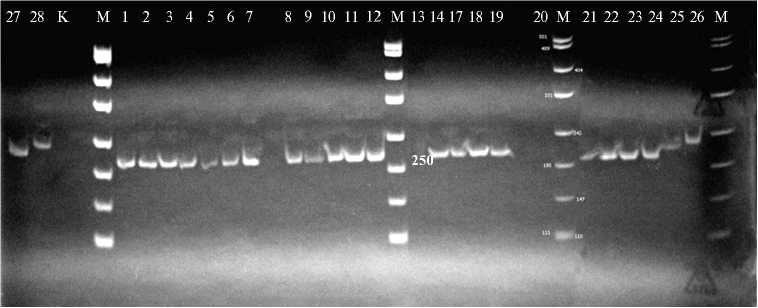
Б
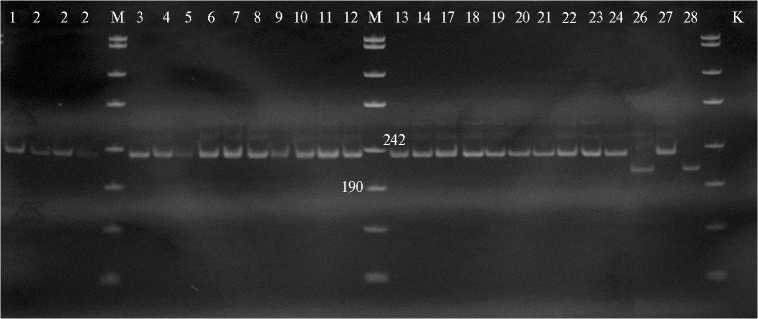
Микросателлитные профили у образцов риса ( Oryza sativa L.) российской, казахстанской и мировой селекции в локусах RM 493 (А) и AP 3206 (Б) : 1 — Маржан, 2 — Курчанка, 3 — F 4 Маржан ½ Курчанка var. dichroa ; 4 — F 4 Маржан ½ Курчанка var. zeravschanica , 5 — F 4 Маржан ½ Курчанка var. italica , 6 — F 4 Маржан ½ Курчанка var. vulgaris , 7 — Регул, 8 — F 3 Регул ½ Курчанка var. italica , 9 — F 3 Регул ½ Курчанка var. zeravschanica , 10 — F 3 Регул ½ Кур-чанка var. dichroa , 11 — Соната, 12 — Лиман, 13 — F 4 Соната ½ Лиман var. italica , 14 — F 4 Соната ½ Лиман var. nigroapiculata , 17 — F 3 Кубань 3 ½ коллекционный образец 34-09 var. zeravschanica , 18 — Ханкайский 429, 19 — F 4 Ханкайский 429 ½ Курчанка var. dichroa , 20 — Баканасский, 21 — Аналог II, 22 — F 4 Баканасский ½ Аналог II var. nigroapiculata , 23 — Дарий 23, 24 — F 3 Дарий 23 ½ Аналог II var. italica , 25 — FL 478 HB9093, 26 — BINA dhan8 HB9106, 27 — BRRI dhan47 HB9114, 28 — Мадина, К — контроль (Н 2 О); M — маркер молекулярных масс (п.н.) 100 bp (OOO «Синтол». Г. Москва).
Для ПЦР-анализа ДНК (рис.) мы использовали микросателлитные маркеры RM 493 и AP 3206, картированные внутри Saltol региона на 1-й хромосоме. Сравнение микросателлитных профилей у исследованных образцов позволяет сделать вывод о наличии в локусе RM 493 трех аллелей: первый несли генотипы BINA dhan8 HB9106 (солеустойчивый стандарт, IRRI) и сорт Мадина казахстанской селекции; второй — генотипы FL 478 HB9093 и BRRI dhan47 HB9114 (солеустойчивые стандарты, IRRI); третий — все остальные образцы (см. рис.). Интересно отметить, что в число последних попали и солеустойчивые стандарты: сорт Маржан казахстанской селекции и сорт Курчанка российской селекции. Солеустойчивость указанных российских и казахстанских генотипов риса, скорее всего, была обусловлена другими QTL.
Использование AP 3206 также позволило выявить полиморфизм, но этот микросателлитный маркер оказался менее информативным по сравнению RM 493 (см. рис.). Профили ПЦР-продуктов были одинаковы почти у всех исследуемых образцов, кроме двух — BINA dhan8 HB9106 (дорожка 26) и Мадина (дорожка 28), что свидетельствует о существовании двух аллелей солеустойчивости по локусу AP 3206.
Маркер-опосредованная селекция (MAS, marker-assisted selection), широко применяемая в настоящее время (18), в том числе для повышения солеустойчивости растений, позволила добиться значительного прогресса по сравнению с традиционными методами (19-22). С ее помощью можно существенно сокращать время, необходимое для создания новых генотипов, оценивать чистоту сортового материала и сортовую идентичность, изучать генетическое разнообразие современных сортов, картировать гены и локусы количественных признаков (QTL), выявлять маркеры, тесно сцепленные с генами и QTL целевых признаков, контролировать типы скрещиваний и интрогрессию генов и QTL в различных схемах MAS, выполнять пирамидирование генов (23, 24).
У риса солеустойчивость контролируется несколькими независимыми локусами, ответственными за формирование признака на разных стадиях онтогенеза. Одними из ключевых служат SKC1 и Saltol . Saltol относится к QTL и был картирован у солеустойчивого сорта риса Pokkali на 1-й хромосоме (25, 26). Технология молекулярно-генетического скрининга солеустойчивости позволяет оперативно отбирать формы растений с целевыми генами и создавать сорта с заданными свойствами без использования провокационных фонов (27, 28). По данным литературы, введение Saltol QTL в высокопродуктивные сорта стабилизирует урожайность у риса на засоленных почвах (29). В настоящее время с целью интрогрессии Saltol QTL в казахстанские сорта риса нами уже проведена гибридизация между сортами Баканасский, Маржан, Мадина, Кубань 3 и донорами солеустой-чивости BINA dhan 8HB 9106, FL 478 HB 9093, BRRI dhan 47 HB 9114 (данные не представлены). Следующими этапами в селекционной схеме будет серия беккроссов с казахстанскими сортами в качестве реципиентных форм с последующим испытанием полученных линий в полевых условиях, ДНК-идентификация Saltol -локусов у этих линий и создание на их основе солеустойчивых казахстанских сортов риса.
Сообщалось, что при молекулярно-генетическом анализе целевых регионов ДНК наиболее перспективны полиморфные SSR маркеры, сцепленные с QTL солеустойчивости, такие как RM 490, RM 1287, RM 10694, AP 3206, AP 3206f, RM 8094, RM 3412b, RM 10748, RM 493, RM 10793, SalT 1, RM 562, RM 7075 (30). В наших исследованиях для ранжирования генетической плазмы риса на наличие локуса солеустойчивости Saltol информативным оказался микросателлитный маркер RM 493.
Таким образом, изученные нами генотипы риса разного происхождения при скрининге в фазу проростков показали неодинаковую устойчивость к карбонатному, хлоридному и сульфатному засолению (наиболее токсичным оказалось карбонатное). При молекулярно-генетическом анализе контрастных по изучаемому признаку образцов из двух использованных микросателлитных маркеров полиморфизм проявил RM 493, картированный внутри Saltol региона. У исследуемых генотипов обнаружены три аллеля этого микросателлитного локуса, из которых один характерен для большинства изученных генотипов, а из двух других один обнаружен у двух филиппинских сортов, другой — у филиппинского и казахского.
Итак, в результате скрининга 34 генотипов риса (сорта, сортообраз-цы, коллекционные образцы российской, казахстанской и филиппинской селекции и их разные гибридные комбинации) выявлены наиболее перспективные солеустойчивые сорта и гибриды, которые можно выращивать на засоленных почвах, а также использовать как исходной материал для селекции более солетолерантных форм. Гибрид F2 Ханкайский 429 ½ коллекционный образец 4-09, а также сорта Маржан и Мадина накапливали максимальную в процентном соотношении биомассу при трех типах засоления (хлоридном, сульфатном и карбонатном). Следовательно, эти генотипы представляют ценность как исходный материал в селекционных программах по повышению солеустойчивости. Для контроля интрогрессии локуса Saltol может использоваться полиморфный микросателлитный маркер RM 493, который оказался наиболее информативным при анализе контрастных по изучаемому признаку генотипов.
Список литературы Оценка и отбор исходного материала для селекции солеустойчивых сортов риса в Республике Казахстан
- Ткаченко Ю.А., Досеева О.А. Содержание натрия, калия и кальция в органах растений риса в разные фазы их вегетации в условиях хлоридного засоления. Рисоводство, 2007, 10: 59-66.
- Benduhn F., Renard P. A dynamic model of the Aral Sea water and salt balance. J. Marine Syst., 2004, 47: 35-50.
- Умирзаков С.И. Инновационный путь развития рисоводства Казахстана: проблемы и перспективы. Мат. Межд. науч.-практ. конф. «Научно-инновационные основы развития рисоводства в Казахстане и странах зарубежья». Кызылорда, 2012: 17.
- Cтроганов Б.П., Клышев Л.К., Азимов Р.А. Проблемы солеустойчивости растений. Ташкент, 1989.
- Hosseini S.J., Tahmasebi S.Z., Pirdashti H. Screening of rice (Oryza sativa L.) genotypes for NaCl tolerance at early seedling stage. International Journal of Agronomy and Plant Production, 2012, 3(8): 274-283.
- Abbas M.K., Ali A.S., Hasan H.H., Ghal R.H. Salt tolerance study of six cultivars of rice (Oryza sativa L.) during germination and early seedling growth. J. Agr. Sci., 2013, 5(1): 250-258 ( ) DOI: 10.5539/jas.v5n1p250
- Bhowmik S.K., Titov S., Islam M.M., Siddika A., Sultana S., Shahidul Haque M.D. Phenotypic and genotypic screening of rice genotypes at seedling stage for salt tolerance. Afr. J. Biotechnol., 2009, 8(23): 6490-6494.
- Белецкий Ю.Д., Шевякова Н.И., Карнаухова Т.Б. Пластиды и адаптация растений к засолению. Ростов-на-Дону, 1990.
- Tiwari S., Krishnamurthy S.L., Kumar V., Singh B., Rao A.R., Mithra A., Rai V., Singh A.K., Singh N.K. Mapping QTLs for salt tolerance in rice (Oryza sativa L.) by bulked segregant analysis of recombinant inbred lines using 50K SNP chip. PLoS ONE, 2016, 11(4): e0153610 ( ) DOI: 10.1371/journal.pone.0153610
- Удовенко Г.В. Особенности различных методов оценки солеустойчивости расте-ний. В сб.: Методы оценки устойчивости растений к неблагоприятным факторам среды. Л., 1976: 228-238.
- Akbar M., Yabuno T., Nakao S. Breeding for saline resistant varieties of rice. I. Variability for salt tolerance among some rice varieties. Jpn. J. Breed., 1977, 22: 277-284.
- Ерыгин П.С., Красноок Н.П. Основы биологии риса. М., 1965: 15-33.
- Скаженник М.А., Воробьев Н.В., Досеева О.А. Методы физиологических исследований в рисоводстве. Краснодар, 2009: 23.
- Singh R.K., Gregorio G.B., Ali S., Arceta M., Mohammadi R., Vispo N.A., Amas J., Thomson M., Gautam R., Brar D.S., Ismail A. Molecular screening and diversity of salt tolerance in rice genotypes. FSC Brief No 9. Food Security Center, Germany, 2011.
- Murray M.G., Thompson W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res., 1980, 8(19): 4321-4325 ( ) DOI: 10.1093/nar/8.19.4321
- Huyen L.T.N., Cuc L.M., Ismail A.M., Ham L.H. Introgression the salinity tolerance QTLs Saltol into AS996, the elite rice variety of Vietnam. American Journal of Plant Sciences, 2012, 3: 981-987.
- Амирова С. Влияние различных типов засоления на проницаемость мембран клеток корней риса. Тез. докл. IV Всес. симп. «Физиологические и биохимические основы солеустойчивости растений». Ташкент, 1986: 76.
- Строганов Б.П. Физиологические основы солеустойчивости растений. М., 1962.
- Леонова И.Н. Молекулярные маркеры: использование в селекции зерновых культур для идентификации, интрогрессии и пирамидирования генов. Вавиловский журнал генетики и селекции, 2013, 17(2): 314-325.
- Gregorio G.B., Senadhira D., Mendoza R.D., Manigbas N.L., Roxas J.P., Guerta C.Q. Progress in breeding for salinity tolerance and associated abiotic stresses in rice. Field Crops Res., 2002, 76: 91-101 ( ) DOI: 10.1016/S0378-4290(02)00031-X
- Yamaguchi Y., Blumwald E. Developing salt-tolerant crop plants: challenges and opportunities. Trends Plant Sci., 2005, 10: 615-620 ( ) DOI: 10.1016/j.tplants.2005.10.002
- Ismail A.M., Heuer S., Thomson M.J., Wissuwa M. Genetic and genomic approaches to develop rice germplasm for problem soils. Plant Mol. Biol., 2007, 65: 547-570 ( ) DOI: 10.1007/s11103-007-9215-2
- Hu S., Tao H., Qian Q., Guo L. Genetic and molecular breeding for salt-tolerance in rice. Rice Genomics and Genetics, 2012, 3(7): 39-49 ( ) DOI: 10.5376/rgg.2012.03.0007
- Blumwald E., Grover A. Salt tolerance. In: Plant biotechnology: current and future uses of genetically modified crops/N.G. Halford (ed.). UK, John Wiley & Sons, Ltd, 2006: 206-224 ( ) DOI: 10.1002/0470021837.ch11
- Linh L.H., Linh T.H, Xuan T.D., Ham L.H., Ismail A.M., Khanh T.D. Molecular breeding to improve salt tolerance of rice (Oryza sativa L.) in the Red River Delta of Vietnam. International Journal of Plant Genomics, 2012: Article ID 949038 ( ) DOI: 10.1155/2012/949038
- Waziri A., Kumar P., Purty R.S. Saltol QTL and their role in salinity tolerance in rice. Austin J. Biotechnol. Bioeng., 2016, 3(3): 1067.
- Islam M.R., Salam M.A., Hassan L. QTL mapping for salinity tolerance at seedling stage in rice. J. Agric. Food Technol., 2011, 23: 137-146.
- Sabouri H., Rezai A.M., Moumeni A. QTLs mapping of physiological traits related to salt tolerance in young rice seedlings. Biologia Plantarum, 2009, 53: 657-662 ( ) DOI: 10.1007/s10535-009-0119-7
- Hien Thi Thu Vu, Duc Duy Le, Abdelbagi M. Ismail, Ham Huy Le. Marker-assisted backcrossing (MABC) for improved salinity tolerance in rice (Oryza sativa L.) to cope with climate change in Vietnam. Australian Journal of Crop Science, 2012, 6(12): 1649-1654.
- Thomson M.J., de Ocampo M., Egdane J., Rahman M.A., Sajise A.G., Adorada D.L., Tumimbang-Raiz E., Blumwald E., Seraj Z.I., Singh R.K., Gregorio G.B., Ismail A.M. Characterizing the Saltol quantitative trait locus for salinity tolerance in Rice. Rice, 2010, 3: 148-160 ( ) DOI: 10.1007/s12284-010-9053-8
