Оценкa потенциала секвестрации углерода у быстрорастущих тополей

Автор: Евлаков П.М., Гродецкая Т.А., Евтушенко Н.А.
Журнал: Биология в сельском хозяйстве @biology-in-agriculture
Рубрика: Актуальные вопросы зоотехнии и ветеринарии
Статья в выпуске: 4 (41), 2023 года.
Бесплатный доступ
Изменение климата затрагивает жизнедеятельность растений, что обусловлено воздействием абиотического стресса, который значительным образом влияет на важнейшие биохимические процессы, включая фотосинтез. В связи с этим необходимо подобрать комплекс методов для оценки параметров фотосинтеза у культурных растений. В статье проведена оценка потенциала секвестрации углерода на карбоновом полигоне Воронежской области у перспективных селекционно-ценных форм и гибридов тополей ( Populus L.) 'Ведуга', 'Э.с.-38', 'ПОК' . В качестве контроля использовался эндемик Воронежской области тополь черный (осокорь). Исследованы морфометрические параметры растений, проведен анализ площади отдельного листа и суммарной ассимиляционной поверхности тополя разных форм и гибридов с применением портативного лазерного измерителя площади листа CI-202 (CID Bioscience, США). Для определения параметров фотосинтеза и транспирации использовали портативную систему измерения газообмена растений CI-340 (CID Bioscience, США). Показано, что суммарная площадь листьев на 1 растение у тополя сорта 'Э.с.-38' значительно (в 5,6 раз) превышала аналогичный показатель у контрольного варианта. Вторым по данному значению выступал тополь белый 'Ведуга', опережающий контроль в 3,3 раза. Значительное преимущество по высоте (на 33% по сравнению с контролем) и площади листьев определило большие в 4,6 раз показатели поглощения CO2 у 'Э.с.-38' за один вегетационный период. Для тополя 'Ведуга' этот показатель составил 2,8 раз. Полученные результаты позволяют характеризовать тополь 'Э.с.-38' в качестве перспективной формы и рекомендовать его для закладки быстрорастущих плантаций с высоким потенциалом секвестрации углерода.
Тополь, изменение климата, абиотический стресс, фотосинтез, депонирование co2, продуктивность
Короткий адрес: https://sciup.org/147242871
IDR: 147242871 | УДК: 577.3+581.1
Assessment of the carbon sequestration potential in fast growing poplars
Climate change affects plant life due to the impact of abiotic stress, which significantly affects the most important biochemical processes, including photosynthesis. In this regard, it is necessary to select a set of methods for assessing photosynthesis parameters in cultivated plants. The article assesses the potential for carbon sequestration at the carbon test site in the Voronezh region for promising selection-valuable forms and hybrids of poplars ( Populus L.) 'Veduga', 'E.s.-38', 'POK'. The endemic black poplar (sedge) of the Voronezh region was used as a control. The morphometric parameters of plants were studied, the area of an individual leaf and the total assimilation surface of poplar of different forms and hybrids were analyzed using a portable laser leaf area meter CI-202 (CID Bioscience, USA). To determine the parameters of photosynthesis and transpiration, a portable system for measuring plant gas exchange CI-340 (CID Bioscience, USA) was used. It was shown that the total leaf area per 1 plant of the poplar variety 'E.s.-38' was significantly (5.6 times) higher than that of the control variant. The second in terms of this value was the white poplar 'Veduga', outstripping the control by 3.3 times. A significant advantage in height (by 33% compared to the control) and leaf area determined the 4.6 times greater CO2 absorption rates of 'E.s.-38' during one growing season. For poplar 'Veduga' this figure was 2.8 times. The results obtained make it possible to characterize poplar 'E.s.-38' as a promising form and recommend it for establishing fast-growing plantations with a high potential for carbon sequestration.
Текст научной статьи Оценкa потенциала секвестрации углерода у быстрорастущих тополей
Введение. Увеличение углекислого газа в атмосфере – глобальный процесс, затрагивающий жизнь растений по всей планете. Накопление CO2 способствует изменению климата в сторону повышения температур и провоцирует возникновение стресса. Засу- ха, засоление, холод, жара, ультрафиолетовое излучение и тяжелые металлы значительным образом усложняют жизнедеятельность сельскохозяйственных и лесных растений. По некоторым оценкам, воздей- ствие стресса может снижать урожайность культурных растений до 50% [1].
Основным жизненно важным процессом, обеспечивающим растения энергией и углеродом, необходимым для синтеза органических соединений, является фотосинтез. Напряженность факторов окружающей среды негативно влияют на эффективность процессов фотосинтеза [2, 3]. Поскольку жизнеспособность и урожайность культурных растений в том числе, обусловлены особенностями их фотосинтетической деятельности, возникает необходимость подробного изучения фотосинтеза для конкретных пород растений, подверженных определенным воздействиям окружающей среды. Понимание особенностей фотосинтеза при влиянии абиотического стресса открывает перспективу для его регуляции и интенсификации, что поможет значительно улучшить продуктивность растений.
Для оценки фотосинтеза необходимо рассмотреть ряд параметров, одним из которых является флуоресценция хлорофилла. Этот индикатор позволяет анализировать фотохимическое состояние фотосистемы II при повышении уровня углекислого газа в атмосфере, воздействия экстремальных температур или изменения содержания воды в листьях.
Рост концентрации CO2 может способствовать увеличению скорости его фиксации, увеличению биомассы и площади листьев, что обусловлено активностью рибулозобисфосфаткарбоксилазы (Рубиско), фермента, обеспечивающего присоединение углекислого газа к рибулозо-1,5-бисфосфату на первой стадии восстановительного пентозофосфатного цикла [4]. Акклиматизация в виде увеличения скорости фотосинтеза в условиях повышенного содержания углекислого газа происходит за счет изменения скорости функционирования Рубиско или её концентрации. Стимуляция ростовых процессов в условиях повышенной концентрации CO2 происходит благодаря оп- тимизации расхода воды фотосинтезирующими органами растений, что отражается в повышении показателя WUE (water-use efficiency), определяемого как отношение фотосинтеза к транспирации [5].
Воздействие неблагоприятных условий окружающей среды провоцирует снижение устьичной проводимости, развитие окислительного стресса и снижение активности Рубиско, увеличивает нефотохимическое гашение (NPQ). Показатель NPQ характеризует количество энергии, рассеивающимся в виде тепла относительно линейного переноса электронов через фотосистемы. Повышенные концентрации углекислого газа могут способствовать снижению NPQ, стимуляции фотосинтеза и улучшению биохимии листьев.
Учитывая вышеизложенное, при оценке фотосинтетического ответа у растений в условиях неблагоприятного воздействия окружающей среды, необходимо поводить анализ параметров морфометрии, анатомии и биохимии листа, измерения активности ключевых ферментов фотосинтеза листа, ферментов антиоксидантного комплекса. Флюоресценция хлорофилла и газообмен листа являются важнейшими параметрами, необходимыми для полноценной характеристики фотосинтетической деятельности растений.
Цель исследований
Целью данного исследования была оценка потенциала углерододепонирующей способности быстрорастущих тополей ( Populus L.).
Условия, материалы и методы
Объект исследования. В качестве объекта исследования использовали селекционно-ценные быстрорастущие сорта и гибриды тополей второго года произрастания (таблица 1, рисунок 1) [6, 7].
Таблица 1 - Характеристика перспективных биотипов тополей
|
№ п/п |
Наименование тополя |
Инв. № |
Происхождение |
Автор гибрида |
|
Белые тополя с пирамидальной формой кроны |
||||
|
1 |
'Ведуга' |
26-07 |
P. alba L. × P. bolleana Laurche |
Царев А.П. |
|
Межсекционные гибриды |
||||
|
2 |
'Э.с.-38'* |
49 |
P. deltoides Marsh. × P. bal-samifera L. |
Вересин М.М. |
|
Черные тополя с пирамидальной формой кроны |
||||
|
3 |
'ПОК' ** |
91 |
P. piramidalis Ros. × P. nigra L. |
Альбенский А.В. |
'Э.с.-38'* – 'Элитный сеянец-38'
'ПОК' ** - 'Пирамидально-осокоревыйКамышинский'

!V:^S- с.-38' второго года вегетации
Рис. 1 – Внешний вид тополя сорта 'Э.
Исследование уровня поглощения углекислого газа у перспективных тополей проводилось на экспериментальном полигоне, заложенном в Воронеже (51.660781, 39.200296) в 2021 году. Контрольная группа состояла из представителей эндемичного для Воронежской области вида тополя черного (осокоря) P. nigra L.
У опытных и контрольных образцов проводили оценку морфометрических параметров, площади ли- ста и ассимиляционной поверхности с использованием лазерного измерителя CI-202 (CID Bioscience, США).
Для измерения скорости поглощения углекислого газа листом in vivo использовали инфракрасный газовый анализатор CI-340 (CID Bioscience, США), позволяющий оценить ряд фотосинтетических параметров в режиме реального времени (Таблица 2).
Таблица 2 – Интерпретация параметров фотосинтеза,
|
анализируемых при помощи инфракрасного газового анализатора |
||
|
Обозначение параметра |
Единица измерения |
Интерпретация параметра |
|
CO 2 in |
мл/м3 |
Количество CO на входе в анализатор |
|
CO 2 out |
мл/м3 |
Количество CO на выходе из листовой камеры |
|
H 2 O in |
кПа |
Количество H 2 O на входе в анализатор |
|
H 2 O out |
кПа |
Количество H 2 O на выходе из анализатора |
|
PAR (ФАР) |
мкмоль/м2/с |
Фотосинтетически активная радиация |
|
W |
моль/м2/с |
Молярный поток (массовый расход на площадь листа) |
|
E |
ммоль/м2/с |
Скорость транспирации |
|
T воздуха |
°С |
Температура окружающего воздуха в листовой камере |
|
T листа |
°С |
Температура листа, измеренная инфракрасным датчиком температуры |
|
ATM |
кПа |
Атмосферное давление |
|
Pn |
мкмоль/м2/с |
Чистая скорость фотосинтеза (нетто) |
Параметры газообмена и транспирации были измерены у растений второго года жизни на листьях, которые находились в верхнем ярусе и были хорошо освещены. Листья не отделяли от растения перед проведением измерений. Повторность анализа для каждой точки была 3-5 кратной. Расчет нетто фотосинтеза прибором осуществляли при помощи встроенного программного обеспечения по следующей формуле:
v x p
Pn = -tv X (Co - G) = -200S39 X ^—7 X (C, - Cj)
где Pn – скорость наблюдаемого фотосинтеза для открытых систем (мкмоль м-2 с-1);
W – массовый расход на площадь листа (моль м-² с-1) для открытых систем;
C 0 ( C i ) – концентрация CO 2 на выходе (входе) в газоанализатор (мл м-3 или мкмоль моль-1);
V – скорость потока газа через кювету (л мин-1);
P – атмосферное давление (бар);
T a – температура воздуха (K);
A – площадь листа (см2).
Статистическую обработку проводили общепринятыми методами [8] с использованием программы Statistica и MS Excel.
Результаты и обсуждение
Показатели фотосинтетического усвоения углекислоты у исследованных генотипов тополей находились примерно на одном уровне и варьировались от 25,1 у 'ПОК' до 30,7 у осокоря (рисунок 2).
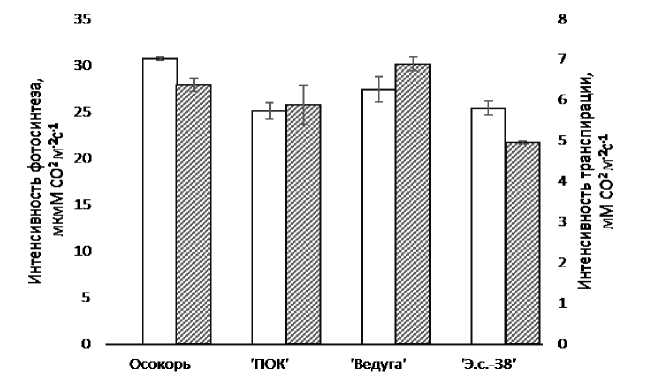
Рис. 2 – Фиксация CO2 в ходе фотосинтеза у селекционно-ценных сортов и гибридов тополей.
Белые столбцы - интенсивность фотосинтеза в мкМм-2 с1, заштрихованные столбцы - интенсивность транспирации в мМм- 2 с-1 (n=30, M±SEM)
Промежуточное положение по уровню результирующего углеродного газообмена занимали 'Эс-38' и белый тополь сорта 'Ведуга'. По имеющимся данным [9], максимальные показатели интенсивности фотосинтеза не превышали 25 мкмМ CO2 м”2 с”1, однако в проведенном исследовании отмечены значительно превышающие этот порог значения. Скорость поглощения углекислоты на одно растение является результатом произведения суммарной площади листа одного растения и интенсивности фотосинтеза. Данный показатель у 'Э.с.-38' был на 37% выше, чем у белого тополя 'Ведуга' и в 4 и более раз превышал значение поглощения CO2 у генотипов 'ПОК' и осокорь. Высокие показатели фотосинтетической ассимиляции CO2, как правило, обусловлены увеличенной активностью комплекса Рубиско. Однако, по мнению ряда исследователей [10], чистая ассимиляция углекислого газа в процессе фотосинтеза у C3 растений не является простым балансом между фиксацией CO2 посредством карбоксилирования и производством CO2 при фотодыхании, но включает реакции «вспомогательного метаболизма». То есть, утилизация углерода и азота путями, отличными от цикла Кальвина и фотоды-хательного цикла, а также быстрые сигнальные события могут влиять на наблюдаемую скорость чистого фотосинтеза и требуют дополнительного изучения.
Было показано, что межсекционный гибрид 'Эс-38' имел наибольшую суммарную площадь листьев среди всех проанализированных тополей, с результатом 7,5 м2 (рисунок 3).
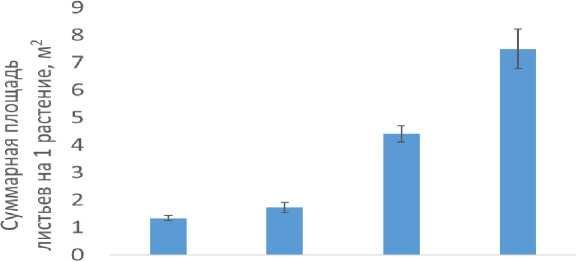
Осокорь 'ПОК' 'Ведуга' 'Э.С.-38'
Рис. 3 – Значения суммарной ассимиляционной поверхности у исследованных генотипов тополей
Сорт 'Ведуга' занимает второе место по площади с показателем 4,4 м2. При этом 'ПОК' и осокорь сформировали наименьшие суммарные ассимиляционные поверхности на одно растение. В результате проведенного анализа было установлено, что в ювенильном возрасте межсекционный гибрид 'Э.с.-38' превосходит другие исследованные сорта тополей по показателям роста. Кроме того, у этого гибрида формируется наибольшая ассимиляционная поверхность на одно растение.
Морфометрические параметры гибрида тополя 'Э.с.-38', первоначально названного М.М. Вересиным «Воронежским гигантом», значительно превышали аналогичные показатели других исследованных генотипов тополей. Его высота (399.1 см) превосходила контрольные растения осокоря на 33%, а диаметр корневой шейки составлял 52,4 мм, что на 38-49% больше по сравнению с другими формами тополей. Площадь отдельного листа у гибрида 'Эс-38' также была вдвое больше, чем у других исследованных генотипов, и составляла в среднем 103,3 см2. Получен- ные результаты подтверждают выводы проведенных ранее исследований [11].
Густота закладки анализируемой плантации соответствовала 2500 саженцев с ЗКС (закрытой корневой системой) на гектар, соответственно стандарту, что учитывалось при расчете депонирующей способности исследуемых растений. Согласно данным Цельникер Ю.Л. за три летних месяца среднее суточное время фотосинтеза составляло 14,6 часов, а темное время – 9,4 часов [12]. В нашем случае, при длительности среднесуточного фотосинтетического светового периода в 12 часов и продолжительности вегетационного периода в 5 месяцев, расчетная максимальная депонирующая способность плантационных насаждений тополя 'Э.с.-38' показала максимальное значение, составив 138,7 тонн на гектар (рисунок 4).
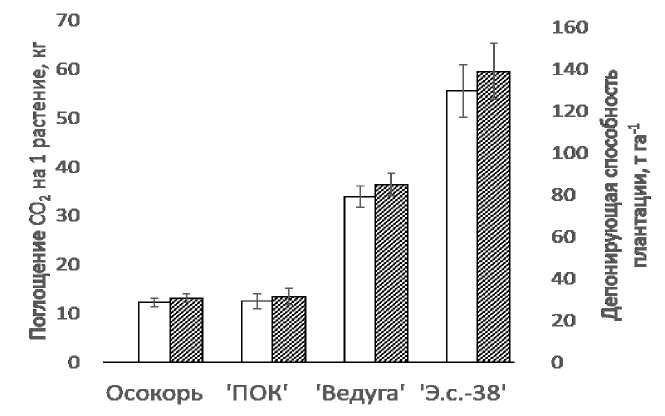
Рис. 4 – Значения депонирования CO2 у различных сортов и гибридов тополей. Белые столбцы – поглощение CO 2 на 1 растение, заштрихованные столбцы – депонирующая способность плантации, т га-1 (n=30, М±SEМ)
Тополя осокорь и 'ПОК' показали значительное отставание по результатам депонирования углекислоты со значениями 30,6 и 31,3 т га-1, соответственно.
Выводы
На основе оценки углерододепонирующей способности различных генотипов тополя в Воронежской области можно сделать вывод, что сорт 'Э.с.-38' «Воронежский гигант» является непревзойденным лиде- ром с высоким потенциалом ассимиляции углерода и может быть рекомендован для создания плантаций быстрорастущих деревьев. Этот потенциал обеспечивается скоростью роста, определяющей формирование максимальной ассимиляционной поверхности с высокой скоростью усвоения углекислого газа листовой пластинкой.
Список литературы Оценкa потенциала секвестрации углерода у быстрорастущих тополей
- Ahmadi-Lahijani M. J., Moori S. Photosynthetic Response and Adaptation of Plants in Perspective of Global Climate Change / M. J. Ahmadi-Lahijani, S. Moori – London, Изд- во IntechOpen, 2023 – 284 с.
- Mechanisms regulating the dynamics of photosynthesis under abiotic stresses / I. Muhammad [et al.] // Frontiers in plant science. 2021. V. 11. P. 615942.
- Photosynthetic response of plants under different abiotic stresses: a review / A. Sharma [et al.] // Journal of Plant Growth Regulation. 2020. V. 39. P. 509-531.
- Effects of water stress on photosynthesis, yield, and water use efficiency in winter wheat / W. Zhao [et al.] // Water. 2020. V. 12. №. 8. P. 2127.
- Iñiguez C., Aguiló-Nicolau P., Galmés J. Improving photosynthesis through the enhancement of Rubisco carboxylation capacity // Biochemical Society Transactions. 2021. V. 49. №. 5. P. 2007-2019.
- Царев А. П., Царев В. А. Биомасса тополей подрода Eupopulus Dode для производства биоэнергии // Лесной вестник/Forestry bulletin. 2015. Т. 19. №. 6. С. 57-62.
- Tsarev A. P. Aminoacids composition and fodder value of poplar leaves by minirotation cultivating //Poplars and willows: from research models to multipurpose trees for a bio-based society'. 2010. P. 135-135.
- Лакин Г.Ф. Биометрия /Г.Ф. Лакин – М.: Высшая школа, 1990. – 350 с.
- Ceulemans R., Saugier В. Photosynthesis. In: Raghavendra AS (ed) // Physiology of Trees. New York: Wiley, 1991. P. 21-50.
- Tcherkez G., Limami A. M. Net photosynthetic CO2 assimilation: more than just CO2 and O2 reduction cycles // New Phytologist. 2019. V. 223. №. 2. P. 520-529.
- Евлаков П.М., Царев А.П., Заплетин В.Ю. Изучение фотосинтетических особенностей и интенсивности транспирации у различных сортов и клонов тополя (Populus L.) // Труды Санкт-Петербургского научно-исследовательского института лесного хозяйства. 2017, №. 4. С. 4-13.
- Цельникер Ю.Л. Физиологические основы теневыносливости древесных растений / Ю.Л. Цельникер. – М: Наука, 1978. – 215 с.
