Оценка разнообразия цианобактерий и водорослей эпилитона р. Сылвы (Пермский край) морфологическими и молекулярными методами
 морфологическими и молекулярными методами")
Автор: Саралов А.И., Беляева П.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 1, 2018 года.
Бесплатный доступ
Исследовано разнообразие альгоценозов эпилитона среднего течения р. Сылвы. Применен ком-плекс морфоцитологических методов альгологии и современной молекулярной экологии. Проведе-на полимеразная цепная реакция с использованием олигонуклеотидных праймеров для избира-тельной амплификации, клонирования и секвенирования фрагментов генов 16S рРНК цианобакте-рий и пластид. В конце аномально теплого лета 2016 г. в эпилитоне р. Сылвы выявлено 98 видов водорослей из трех отделов: Bacillariophyta (80% по биомассе - В; 40% по численности -N), Chlorophyta (15% В; 10% N) и Cyanophyta/Cyanobacteria (5% В; 50% N). По результатам филогене-тического анализа генов 16S рРНК 142 клонов, 80 отнесено к диатомовым и 2 к зеленым водорос-лям, 60 - к цианобактериям. В фрагментах пластидных геномов Bacillariophyta эпилитона выявле-но две филогенетически обособленных ветви клонов, родственные представителям с разным строением панциря и отношением к солености воды. Причем 89% видов диатомовых были детек-тированы в эпилитоне и морфологическими, и молекулярными методами. Результаты исследова-ния генов 16S рРНК хлоропластов Bacillariophyta эпилитона хорошо согласовались с их естествен-ной филогенетической системой, основанной на морфологии и детальной структуре кремневого панциря. Напротив, морфотаксономические признаки цианобактерий эпилитона слабо согласова-лись с данными молекулярного анализа на основе сравнения нуклеотидных последовательностей генов 16S рРНК.
Эпилитон, цианобактерии, 16s ррнк, диатомовые водоросли, зеленые водоросли, таксономия
Короткий адрес: https://sciup.org/147204873
IDR: 147204873 | УДК: 577.29:574.586 | DOI: 10.17072/1994-9952-2018-1-97-104.
Diversity assessment of cyanobacteria and algae of Sylva river epiliton (Perm kray) by morfological and molecular methods
Algocenosis diversity of the epilithon to the middle reach of submountain Sylva river (Perm kray) was studied. A complex of morphological and cytological methods of algology and modern molecular ecology was applied. Polymerase chain reaction (PCR) was conducted using oligonucleotide primers for selective amplification, cloning and sequencing of 16S rRNA genes' fragments of cyanobacteria and plastids, but not for bacteria and archaea. By the end of anomalous hot summer 2016, 98 algal species from three groups within the epilithon of the Sylva river - Bacillariophyta (80% of biomass - B; 40% of number - N), Chlorophyta (15% B; 10% N) and Cyanophyta/Cyanobacteria (5% B; 50%N) - were identified. 142 clones from the PCR-products were selected and detected to the phylogenetic analysis: 80 species were belonged to diatoms, 2 - to green algae and 60 - to Cyanobacteria. In 16S rRNA genes' fragments, local-ized in chloroplast genomes of Bacillariophyta from the epilithon, two phylogenetically distinct groups of clones were revealed that have different frustule structure and sensitivity to water salinity. The analysis of 16S rRNA genes' fragments, localized in Bacillariophyta's chloroplast genomes well corresponded to natural phylogenetic system based on morphology and detailed structure of silica frustule. Notably, 89% of diatom species were detected within the epilithon using morphological and molecular methods. Mor-phological characteristics of cyanobacteria from the epilithon poorly corresponded to their phylogeny based on the comparison of 16S rRNA genes' nucleotide sequences.
Текст научной статьи Оценка разнообразия цианобактерий и водорослей эпилитона р. Сылвы (Пермский край) морфологическими и молекулярными методами
ной экологии [Nübel et al., 2000; O'Sullivan, Weightmann, Fry, 2002; Bricheux et al., 2013; Lindemann et al., 2013; Семейкина и др., 2015].
Фито- и бактериоперифитону водотоков Камского бассейна, в частности, в наиболее изученной нами предгорной р. Сылве Пермского края, принадлежит ведущее положение в самоочищении и биопродуктивности, азотфиксации и денитрификации [Беляева, 2004; Беляева и др., 2007; Саралов и др., 2010]. В настоящее время согласно морфоцитологическим данным в перифитоне р. Сылвы отмечена четкая тенденция увеличения численности и разнообразия Cyanophyta/Cyanobacteria за счет представителей родов Snowella, Merismopedia, Gloeotrichia, Oscillatoria и сокращение Bacillario-phyta родов Navicula, Synedra, Cymbella [Беляева, 2014]. В связи с этим возникает необходимость в оценке биоразнообразия и состава альго-бактериальных сообществ перифитона предгорной реки с использованием комплекса альгологических и молекулярных методов.
Цель работы – с применением молекулярнобиологических и общепринятых в альгологии морфоцитологических методов охарактеризовать разнообразие цианобактерий, диатомовых и зеленых водорослей в образцах эпилитона р. Сылвы.
Материалы и методы
Река Сылва – предгорная река Среднего Урала, впадает в Чусовской залив Камского водохранилища. Каменистые субстраты среднего течения р. Сылвы занимают около 70% площади, состоят из песка (<0.1 см), гравия, гальки и валунов (диаметром 0.1–1.5 м). Пробы эпилитона отбирали в середине августа 2016 г. выше г. Кунгура в районе учебно-научной базы «Предуралье» Пермского университета. Лето 2016 г. в Пермском крае по характеру погоды отличалось преобладанием очень теплой погоды и значительным дефицитом осадков. Средняя температура воздуха за лето 2016 г. превысила норму на 2.6°C. Самым жарким днем лета на большей части края стало 15 августа, до 35.6°С [Климатические…, 2016].
Для более полного анализа разнообразия водорослей и цианобактерий альгологические пробы отбирали в августе, при максимальном числе видов и биомассе альгоценозов [Беляева, 2014]. Пробы обрабатывали по общепринятым методам, описанным нами ранее [Беляева, 2004; Беляева и др.., 2007]. Таксономическую принадлежность водорослей устанавливали по отечественным и зарубежным определителям и справочникам пресноводных водорослей [Забелина и др., 1951; Голлер-бах, Косинская, Полянский, 1953; Паламарь-Мордвинцева, 1982; Куликовский и др., 2016; Komàrek, Fott, 1983; Krammer, Lange-Bertalot, 1986–1991; Komàrek, Anagnostidis, 1999]. Назва- ния таксонов приведены согласно классификации, принятой в серии «Определитель пресноводных водорослей СССР» [Забелина и др., 1951; Голлер-бах, Косинская, Полянский, 1953; Паламарь-Мордвинцева, 1982].
ДНК выделяли по методике, основанной на модифицированном методе щелочного выделения ДНК Бирнбойма-Доли и Wizard-технологии фирмы Promega (США) [Булыгина и др., 2002]. Очищенную ДНК хранили при -20 ° С.
Амплификацию фрагментов генов, кодирующих 16S рРНК, проводили с помощью системы праймеров CYA106F (5’ - CGG ACG GGT GAG TAA CGC GTG – 3’), CYA781R(a) (5’ - GAC TAC TGG GGT ATC TAA TCC CAT T – 3’) и CYA781R(b) (5’ - GAC TAC AGG GGT ATC TAA TCC CTT T – 3’) [Nübel, 1997]. Полученные ПЦР-продукты очищали от посторонних примесей и неспецифичных продуктов реакции при помощи электрофореза в легкоплавкой агарозе с применением набора Wizard PCR Preps (“Promega”, США). Очищенные ПЦР-продукты лигировали в вектор pGEM-T Easy System (“Promega”, США), согласно рекомендациям производителя, и клонировали в штамме E. coli DH10B.
Секвенирование ПЦР–продуктов и клонированных фрагментов проводили на секвенаторе ABI3730 DNA Analyzer (“Applied Biosystems”, США) с использованием набора реактивов Big Dye Terminator v 3.1 Cycle Sequencing Kit (“Applied Biosystems”, США), согласно инструкции производителя.
Длина анализируемых последовательностей 16S рРНК составляла 470 нуклеотидов. Первичный сравнительный анализ полученных последовательностей проводили с помощью сервиса NCBI BLAST . Последовательности были проверены на наличие химер с помощью сервиса Bellerophon [Huber et al., 2004]. Для построения филогенетических деревьев использовали программу MEGA 6.0 [Tamura et al., 2013].
Последовательности 16S рРНК были депонированы в GenBank под номерами KY274302– KY274361(цианобактериальные) и KY282959-KY283040 (пластидные).
Результаты и их обсуждение
В период с 2003 по 2016 гг. в р. Сылве в летнем эпилитоне наблюдалось значительное (в 10 раз) увеличение численности и биомассы цианобактерий родов Snowella, Merismopedia и Gloeotrichia echinulata P.G. Richter (таблица).
За этот же промежуток времени численность диатомовых сократилась в 2 раза (особенно Navicula cryptocephala Kütz. и Synedra ulna (Nitzsch) Ehrenb.). Однако они продолжают доми- нировать по биомассе, в частности, за счет получивших массовое распространение крупноклеточных Amphora ovalis (Kütz.) Kütz. и Cocconeis placentula Ehrenb.
Зеленые водоросли в многолетней динамике по данным за 2003–2016 гг. продолжают поддерживать и численность (10–15%), и биомассу (10– 15%) на сравнительно стабильном уровне, в частности, благодаря широко распространенным в планктоне и обрастаниях Cosmarium botrytis Meneg. ex Ralfs, Pediastrum boryanum (Turp.) Meneg. и Closterium spp. В конце аномально теплого лета 2016 г. в эпилитоне р. Сылвы выявлено 98 таксонов водорослей рангом ниже рода из трех основных отделов: Bacillariophyta (80% B; 40% N), Chlorophyta (15% B; 10% N) и Cyanophyta (5% B; 50% N).
Многолетняя динамика структуры эпилитона р. Сылвы (2003–2016 гг.)
|
Показатели |
2003–2005 гг. |
2012 г. |
2014 г. |
2016 г. |
|
Численность, млрд.кл/ м 2 |
8.9±3.6 |
10.8±4.1 |
5.0±1.2 |
4.6±0.4 |
|
Биомасса, г/м2 |
12.7±5.8 |
12.5±4.4 |
28.0±4.8 |
20.9±3.5 |
|
Количество видов |
127 |
122 |
102 |
98 |
|
з m g £ § О S S U p § § is" 5 S 8 I s c s 3 X |
Bacillariophyta : 80%N; 88%В Navicula cryptocephala 15%N; 25%B, Cocconeis placentula 20%B, Synedra ulna 15%B, Diatoma vulgaris 15%N; 5%B, Achnanthes spp. 15%N Chlorophyta : 15%N; 11.5%В Cosmarium botrytis 5%B, Pediastrum boryanum 2%N; 4%B, Closterium spp. 2%B Cyanophyta / Cyanobacteria : 5%N; 0.5%В Planctolyngbya lim-netica , Merismopedia spp. |
Bacillariophyta : 80% N; 85%В Cocconeis placentula 15%N; 50%B, Diatoma vulgaris 10%B, Epithemia sorex 8%B, Achnanthes spp. 10–25%N Chlorophyta : 15%N; 14.5%В Cosmarium spp. 8%B, Pediastrum boryanum 6%B, Crucigenia fenestrata 8%N Cyanophyta/ Cyanobacteria : 5%N; 0.5%В Snowella rosea , Merismopedia spp., Anabaena contorta , Gloeotrichia echinulata |
Bacillariophyta : 70%N; 85%В Cocconeis placentula 10%N; 20%B, Synedra ulna 15%B, Navicula cryptocephala 10%N; 15%B, Cymbella lanceolata 10%B, C. ventricosa 5%N, Achnanthes spp. 15%N Chlorophyta : 15%N; 10%В Pediastrum tetras 4%B, Pediastrum boryanum 2%N; 5%B Cyanophyta / Cyanobacteria : 15%N; 5%В Merismopedia spp. 10%N, Planctolyngbya limnetica , Snowella rosea , Anabaena spp. |
Bacillariophyta : 40%N; 80%В Amphora ovalis 40%B, Cocconeis placentula 10%N; 25%B, Diatoma vulgaris 6%B, Achnanthes spp. 20%N, Cymbella ventricosa , Melosira varians , Nitzschia amphibia Chlorophyta : 10%N; 15%В Scenedesmus spp. 6%N, Pediastrum boryanum 2%N; 5%B, Closterium spp. 5%B, Cosmarium botrytis 3%B Cyanophyta / Cyanobacteria : 50%N; 5%В Snowella rosea 20%N, Merismopedia spp. 13%N, Gloeotrichia echinulata 8%N, Oscillatoria spp. 5%N, Calothrix spp. |
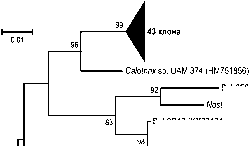
Филогенетический анализ разнообразия альго-бактериальных сообществ эпилитона р. Сылвы по результатам сравнения последовательностей генов 16S рРНК из библиотеки клонированных ПЦР-фрагментов, был проведен достаточно эффективно, идентифицировано 142 клона, 80 отнесено к диатомовым и 2 к зеленым водорослям, 60 – к цианобактериям (рис. 1, 2). На филогенетическом древе клоны с высокой степенью достоверности формировали обособленные кластеры. Основу аль-гофлоры эпилитона р. Сылвы составили широко распространенные пресноводные виды и космополиты (более 80% видового разнообразия), как правило, многочисленные в 16S рРНК-клонотеках пресноводных и морских экосистем [Nübel, Garcia-Pichel, Muyzer, 1997; Nübel et al., 2000; O´Sullivan, Weightmann, Fry, 2002; Bricheux et al., 2013; Lindemann et al., 2013; Komárek et al., 2014].
С использованием «специфичных» праймеров для 16S рРНК цианобактерий на дереве филогении были выявлены последовательности генов 16S рРНК фрагментов пластидных геномов пресноводных зеленых водорослей порядка Desmidiales рода Cosmarium ( C. botrytis ) (рис. 1). Для них характерно наличие примитивных архепластид, чрезвычайное разнообразие типов строения клеток и видов рода Cosmarium (более 200), широкое распространение в обрастаниях макрофитов и камней, в планктоне и бентосе [Паламарь-Мордвинцева, 1982; Keeling, 2013]. Последовательности генов 16S рРНК фрагментов хлоропластных геномов Chlorophyta, однако, филогенетически удалены (как и ядерные геномы) от неродственных Bacillariophyta, у которых пластиды были образованы в высоко развитых формах с кремневым панцирем лишь при третичном эндосимбиозе [Bhattacharya, Medlin, 1995; Keeling, 2013].
В фрагментах генов 16S рРНК, локализованных в хлоропластных геномах Bacillariophyta в эпилитоне р. Сылвы, выявлено две филогенетически обособленные ветви клонов представителей диатомей с разным строением панциря и отношением к солености воды (рис. 1). Единичные представители гетерогенной ветви из 5 родов принадлежат к древнему классу Centricae с панцирем радиального строения (20 клонов: 18 родственны минорным солоноватоводным-морским, эвригалинным Melosira varians и Aulacoseira (Melosira) granulata).
—lAmp/ior^o^/s^ крона)
0.02
I Syl-3A05 (KY282978)
Syl-3B06 (KY282987) 7Л- Cvmatooleura solea (E
721- C^^urasofea (EU580477)
j-Syl-3D06 (KY283002)
pSyl-2E07 (KY282969)
5i|jSyl-3B05 (KY282986)
1 * Pseudo-nitzschia seriata (FJ002178)
— Cymbella subturqidul9 №0519^
Syl-3B01 (KY282983)
Syl-2D01 (KY282967)
" |—Syl-3G12 (KY283033)
. Syl-3B04 (KY282985)
Syl-3F06 (KY283020)
- Didymosphenia geminate (KC509523)
Lsyl-3E12 (KY283016)
-Syl-3E01 (KY283008)
■ Gyrosigma fasciola (AF514847)
1-Syl-2A04 (KY282960)
J Nitzschia frustulum (AY221721) l-Sy|-1E07(KY282959)
-Syl-3F05 (KY283019)
— Nitzschia amphibia (FJ002224)
— Pseudo-nitzschia seriata (FJ002177)
-
—◄ 5 клонов
-
- Synedra acus (JQ088178)
____rSyl-2F10 (KY282972)
о
Л й c
c
Q
!98*— Synedra hyperborea (FJ002235)
I------- Fragilaria zeilleri (EU580508)
53jLsyl-3G08 (KY283030)
^|Lsyl-3F03 (KY283018)
I— Fra*niic№fe (FJ002188)
I----- Achnanthes brevipes (FJ002237)
И—Syl-2A10 (KY282961)
1i-------- Asterionellopsis glacialis (AJ536455)
|rSyl-2H08 (KY282974)
8ol-Syl-3GO1 (KY283025)
— Stephanopyxis nipponica (AJ536465) Syl-3H02 (KY283034)
■ Man(2£l№ (AY221720)
j Syl-3H06 (KY283037)
■ Cyclotella cryplica (FJ002189)
■ Aulacoseira (Melosira} aranulata (FJ002181)
И 4 клона
Closterium baillyanum (KU646494)
I— Cosmarium botrvtis (KU646492) hSyl-3B08(KY282988) ■ Syl-3C02 (KY282993)
0 й CO
и й
c и 0
Я
a 0
"л
E
V
Q
Рис. 1 . Дендрограмма сравнения последовательностей генов 16S рРНК пластид диатомовых и стрептофитовых водорослей эпилитона р. Сылвы
Дендрограмма построена путем сравнения последовательностей длиной 470 нуклеотидов на основании анализа 1000 альтернативных деревьев, алгоритм построения Neighbour-Joining. Цифры указывают достоверность ветвления на основании bootstrap – анализа. Масштабная линейка – 2 нуклеотидные замены на каждые 100 нуклеотидов. Подчеркнуты виды, выявленные и молекулярными, и морфологическими методами
Большинство нуклеотидных последовательностей 16S рРНК на дереве филогении отнесено к классу Pennatae (панцирь моносимметричного строения с перистой структурой). Из 60 фрагментов пластидных геномов диатомей у 34 выявлено сходство с доминантными пресноводными крупноклеточной Amphora ovalis , 6 – субдоминантными пресноводными Cymbella subturgidula / Didymos-phenia geminata , 8 – минорными эвригалинными Nitzschia amphibia / N. frustulum . Около 80% видов диатомовых были детектированы в эпилитоне и морфологическими, и молекулярными методами.
Cyanophyta/Cyanobacteria эпилитона р. Сылвы характеризуются морфологическим многообразием пресноводных видов. Морфологические признаки цианобактерий эпилитона слабо согласовались с их филогенией на основе сравнения нуклеотидных последовательностей генов 16S рРНК. У этих наиболее древних оксигенных фототрофов таксономически значимый ключевой фермент фотоассимиляции углекислоты (RubisCO) несовершенен, обладает очень низким сродством к CO 2 , локализуется в специфичных включениях цитоплазмы – кар-боксисомах α- и β-типа, соответственно у α- и β-цианобактерий [Badger, Hanson, Price, 2002; Espie, Kimber, 2011; Rae et al., 2013].
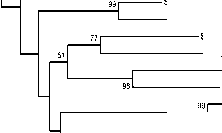
Согласно филогенетическому анализу последовательностей генов 16S рРНК, клоны цианобактерий вошли в две гетерогенные ветви, состоящие из нескольких довольно обособленных кластеров (рис. 2). Большинство клонов первой ветви (75%) проявили высокий уровень сходства с β-цианобактериями (с карбоксисомами β-типа) класса Hormogoneae порядка Nostocales с варьирующими значениями бутстреп-поддержки (92–99%). Только представителям Hormogoneae присущи плазмодесмы – тончайшие протоплазматические тяжи, соединяющие соседние клетки в единое целое в форме многоклеточного нитчатого трихома [Голлербах, Косинская, Полянски й , 1953; Вассер и др., 1989; Komárek et al., 2014].
Основной кластер (45 клонов) примыкает к азотфиксирующим минорным видам эпилитона порядка Nostocales ( Calothrix , Nostoc, Tolypothrix ), формирующих асимметричные трихомы с гетероцистами. Представители этих родов широко распространены на водорослях и макрофитах, камнях и скалах [Вассер и др., 1989].
К основному кластеру порядка Nostocales примыкает обособленный кластер из 5 клонов, родственных доминантным в эпилитоне р. Сылвы видам Snowella класса Chroococcaceae порядка Chroococ-cales . У видов рода Snowella ( S. rosea, S. litoralis ), хотя плазмодесмы отсутствуют, но шаровидные клетки диаметром 3–4 мкм в слизистых колониях располагаются на тонких радиальных разветвлениях–ножках [Голлербах, Косинская, Полянский,
1953]. На периферии первой группы клонов выявлен клон, родственный недостаточно описанному штамму Oscillatoria sp. KNUA009 порядка Oscilla-toriales , объединяющего безгетероцистных представителей класса Hormogoneae [Komárek et al., 2014].
Syl-2F06 (KY274354)
toe insulare SAG 54.79 (KM019927)
Syl-2B12 (KY274342)
j— Calothrix marchica AUS-JR/MT/NT-093 (KX670258)
7^- Tolypothrix sp. PCC 7504 (FJ661002)
---- Snowella rosea 1LM40S01 (AJ781042)
____________________ j—Syl-2D03 (KY274347)
99 П Syl-2A06 (KY274337)
88 —Syl-1D01 (KY274314) — Syl-1D02 (KY274315) Syl-1A07 (KY274304)
Snowella litoralis 1ES42S2 (DQ264220)
।--------------Syl-2C05 (KY274344)
Я---------------- Oscillatoria sp. KNUA009 (HQ201392)
991— Chamaesiphon minutus РОС 6605T (NR_102459)
I—Syl-2B02 (KY274338)
-
99, Cyanobium sp. 0ES35S4 (DQ264240)
99 1 L Syl-2D07 (KY274349)
П_____, Syl-2E02 (KY274351)
I9sL Cyanobium gracile PCC 6307T (NR_102447)
Syl-2F09 (KY274355)
----Pleurocapsa concharum VP3-02 (FR798927)
Syl-2H07 (KY274359)
— Nodosilinea sp. NTRI45 (KP030748)
■ Syl-2D09 (KY274350)
I
z
Hormogoneae
Oscillatoriales
Chamaesiphoneae Pleurocapsales
Pseudanabaena limnetica PUPCCC (KM376980) 8 ■ Limnococcus limneticus Svet06 (GQ375048) J — Syl-1H12 (KY274336)
-----Oscillatoria prolifera UTEX В 1270 (AB075993)£
___1— Syl-2B06 (KY274340)|
"93I— Syl-3A12 (KY274361)“
Рис. 2 . Дендрограмма сравнения последовательностей генов 16S рРНК цианобактерий эпилитона р. Сылвы
Дендрограмма построена путем сравнения последовательностей длиной 470 нуклеотидов на основании анализа 1000 альтернативных деревьев, алгоритм построения Neighbour-Joining. Цифры указывают достоверность ветвления на основании bootstrap – анализа. Масштабная линейка – 2 нуклеотидные замены на каждые 100 нуклеотидов. Подчеркнуты виды, выявленные и молекулярными, и морфологическими методами
Другие клоны эпилитона, близкие α-циано-бактериям (с карбоксисомами α-типа) рода Oscilla-toria, включая холодноводную O. prolifera, вошли во вторую малочисленную группу. Тем не менее, в нее включены морфологически, метаболически и таксономически разнообразные представители классов Hormogoneae, Chroococcaceae, Chamaesi-phoneae, порядков Oscillatoriales, Synechococcales, Pleurocapsales и 5 родов (Oscillatoria, Limnococcus (Chroococcus), Pleurocapsa, Cyanobium, Chamaesi-phon (Sphaerogonium) с высоким значением бутст-реп-поддержки – 99%). Молекулярные исследования последних лет, секвенирование генов большо- го числа видов, полное секвенирование геномов позволило внести существенные коррективы в предшествующие филогенетические системы цианобактерий, в расстановку большого числа видов внутри других родов, семейств, порядков и даже классов [Komárek et al., 2014].
Заключение
Впервые в эпилитоне предгорной р. Сылвы осуществлена параллельная оценка биоразнообразия альго-бактериальных сообществ с использованием современных методов молекулярной экологии и общепринятых в альгологии морфоцитологических методов.
Согласно филогенетическому анализу последовательностей генов 16S рРНК, большинство клонов Cyanobacteria (75%) проявили высокий уровень сходства с представителями порядка Nostocales класса Hormogoneae, с азотфиксирую-щими видами эпилитона, относящихся к β-цианобактериям (с карбоксисомами β-типа). Морфо-таксономические признаки Cyanobacteria слабо согласовались с их филогенией на основе сравнения последовательностей генов 16S рРНК.
Среди Bacillariophyta в эпилитоне р. Сылвы явно преобладает пресноводная крупноклеточная Amphora ovalis (класс Pennatae – с панцирями мо-носимметричного строения). Представители второй гетерогенной ветви принадлежат к древнему классу Centricae с панцирем радиального строения, родственных минорным солоноватоводным-морским, эвригалинным Melosira varians и Aulacoseira ( Melosira ) granulata . Установлено, что диатомовые водоросли сравнительно полно детектируются и современными молекулярными in situ методами и традиционными морфологическими методами водной альгологии. Результаты исследования фрагментов генов 16S рРНК, локализованных в хлоропластных геномах Bacillariophyta, хорошо согласовались с их естественной филогенетической системой, основанной на морфологии и детальной структуре кремневого панциря.
Секвенирование ПЦР–продуктов и клонированных фрагментов выполнено на оборудовании ФГБУН Центр «Биоинженерия» РАН, Москва.
Список литературы Оценка разнообразия цианобактерий и водорослей эпилитона р. Сылвы (Пермский край) морфологическими и молекулярными методами
- Беляева П.Г. Фитоперифитон предгорной реки Сылва (бассейн Камы)//Ботанический журнал. 2004. Т. 89, № 3. С. 435-449
- Беляева П.Г. Состав и структура фитоперифитона реки Сылва (Пермский край)//Ботанический журнал. 2014. Т. 99, № 8. С. 903-916
- Беляева П.Г. и др. Функциональная роль перифитона предгорной реки Сылва (бассейн р. Камы) 102 А. И. Саралов, П. Г. Беляева//Биология внутренних вод. 2007. № 3. С. 32-40
- Булыгина Е.С. и др. Изучение нуклеотидных последовательностей nifH генов у представителей метанотрофных бактерий//Микробиология. 2002. Т. 71, № 4. С. 425-432
- Вассер С.П. и др. Водоросли. Киев, 1989. 608 с
- Голлербах М.М., Косинская Е.К., Полянский В.И. Определитель пресноводных водорослей СССР. Синезеленые водоросли. М., 1953. Вып. 2. 652 с
- Забелина М.М. и др. Определитель пресноводных водорослей СССР. Диатомовые водоросли. М., 1951. 619 с
- Климатические особенности лета 2016 г. в Пермском крае URL: http://accident.perm.ru/index.php/novosti/854summer-2016. (дата обращения: 31.08.2016)
- Куликовский М.С.и др. Определитель диатомовых водорослей России. Ярославль: Филигрань, 2016. 804 с
- Паламарь-Мордвинцева Г.М. Определитель пресноводных водорослей СССР. Зеленые водоросли. Класс Конъюгаты. Порядок Десмидиевые. М., 1982. Вып. 11, ч. 2. 620 с
- Саралов А.И. и др. Азотфиксация и денитрификация в планктоне и перифитоне водотоков Камского бассейна//Биология внутренних вод. 2010. № 2. С. 13-19
- Семейкина П.И. и др. Состав альгобактериального сообщества эпилитона предгорной реки Сылва//Российский иммунологический журнал. 2015. Т. 9 (18), вып. 2(1). С. 606-608
- Badger M.R., Hanson D., Price G.D. Evolution and diversity of CO 2 concentrating mechanisms in cyanobacteria//Functional Plant Biology Journal. 2002. Vol. 29(2-3). P. 161-173
- Bhattacharya D., Medlin L. The phylogeny of plastids: a review based on comparisons of smallsubunit ribosomal RNA coding regions//Journal of Phycology. 1995. Vol. 31. P. 489-498
- Bricheux G. et al. Pyrosequencing assessment of prokaryotic and eukaryotic diversity in biofilm communities from a French river//Microbiology Open. 2013. Vol. 2 (3). P. 402-414
- Espie G.S., Kimber M.S. Carboxysomes: cyanobacterial RubisCO comes in small packages//Photosynthesis Research. 2011. Vol. 109(1-3). P. 7-20
- Huber T., Faulkner G., Hugenholtz P. Bellerophon: a program to detect chimeric sequences in multiple sequence alignments//Bioinformatics. 2004. Vol. 20(14). P. 2317-2319
- Keeling P.J. The number, speed, and impact of plastid endosimbioses in eukaryotic evolution//Annual Review of Plant Biology. 2013. Vol. 64. P. 583-607
- Komàrek J., Anagnostidis K. Cyanoprokaryota. Chroococcales. Süsswasserflora von Mitteleuropa. Jena: FicherVerland, 1999. Bd. 19. T. 1. 548 p
- Komàrek J., Fott B. Chlorophyceae (Gruunalgen), Ordnung: Chlorococcales.Stuttgard: E. Schweizerbart’scheVerlagsbuchhandlung , 1983. Bd. 16. T. 7. H. 1. 1044 S
- Krammer K., Lange-Bertalot H. Bacillariophyceae//Süßwasserflora von Mitteleuropa. Eds. by H. Ettl, J. Gerloff, H. Heying, D. Mollenhauer. Stuttgart-Jena: G. Fisher Verlag, 1986. Bd 2/1. 876 S.; 1988. Bd 2/2. 596 S.; 1991. Bd 2/3. 576 S.; 1991. Bd 2/4. 473 S
- Komárek J. et al. Taxonomic classification of Cyanoprokaryotes (Cyanobacterial genera), using a polyphasic approach//Preslia. 2014. Vol. 86. P. 295-335
- Lindemann S.R. et al. The epsomitic phototrophic microbial mat of Hot Lake, Washington: community structural responses to seasonal cycling//Frontiers in Microbiology. 2013. Vol. 4. P. 1-17
- NCBI BLAST: The National Center for Biotechnology Information. 2016. URL: http://www.ncbi. nlm.nih.gov/blast. (дата обращения: 31.08.2016)
- Nübel U. et al. Matching molecular diversity and ecophysiology of benthic cyanobacteria and diatoms in communities along a salinity gradient//Environmental Microbiology. 2000. Vol. 2. P. 217-226
- Nübel U., Garcia-Pichel F., Muyzer G. PCR primers to amplify 16S rRNA genes from Cyanobacteria//Applied and Environmental Microbiology. 1997. Vol. 63(8). P. 3327-3332
- O'Sullivan L.A., Weightmann A.J., Fry J.C. New degenerate Cytophaga-Flexibacter-Bacteroidesspecific 16S ribosomal DNA-targeted oligonucleotide probes reveal high bacterial diversity in river taff epilithon//Applied and Environmental Microbiology. 2002. Vol. 68. P. 201-210
- Rae B.D. et al. Functions, compositions, and evolution of the two types of carboxysomes: polyhedral microcompaartments that facilitate CO 2 fixation in Cyanobacteria and some Proteobacteria//Microbiology and Molecular Biology Reviews. 2013. Vol. 77(3). P. 357-379
- Tamura K. et al. MEGA6: molecular evolutionary genetics analysis version 6.0//Molecular biology and evolution. 2013. Vol. 30(12). P. 2725-2729
