Оценка реакции ели европейской на воздействие климатических факторов в древостоях разного породного состава

Автор: Стоноженко Л.В., Румянцев Д.Е., Найденова Е.В.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Лесоведение и лесоводство
Статья в выпуске: 4, 2018 года.
Бесплатный доступ
Методами анализа дендроклиматической информации выполнена оценка реакции ели европейской в древостоях разного породного состава на воздействие различных климатических факторов. Выявлена меньшая засухоустойчивость деревьев ели в чистом древостое по сравнению с липово-еловыми древостоями с небольшой долей ели в составе. Рекомендовано в соответствующих лесорастительных условиях Московской обл. создавать смешанные липово-еловые древостои
Дендрохронология, радиальный прирост, ель, липа, породный состав, межвидовая конкуренция
Короткий адрес: https://sciup.org/143166233
IDR: 143166233 | УДК: 630.561.24 | DOI: 10.24419/LHI.2304-3083.2018.4.03
Evaluation of response of Norway spruce on the influence of climatic factors in forest stands of different species composition
The article considers the issue of creation and formation of new plantations in forest areas, passed continuous sanitary logging. Less stability of homogeneous ecosystems is noted, in this connection the creation of pure spruce forests, especially in protective forests, seems to be impractical. In the formation of mixed stands it is necessary to select breeds that would be in a number of positions are complementary to each other and compensate for the impact of certain environmental factors. As a companion species or in the zone of coniferous-broad-leaved forests, the Linden is proposed in view of the presence of natural mechanisms of protection from dry periods, to which spruce is vulnerable.Also in the article the problem of interspecific and intraspecific competition as a factor determining the growth and development of stands is considered. In particular, the influence of this factor on the formation of the radial growth of wood was studied. The aim of this work was to assess the reaction of European spruce in stands of different species composition to the impact of different climatic factors, performed by methods of analysis of dendroclimatic information...
Текст научной статьи Оценка реакции ели европейской на воздействие климатических факторов в древостоях разного породного состава
З асуха 2010 г. и последовавшая за ней катастрофическая вспышка численности короеда-типографа привели к массовому усыханию ельников в Центрально-Европейской части России. В Московской обл., по разным оценкам, подверглись усыханию ельники на площади около 120 тыс. га. Это составляет 6,8% занятых лесными насаждениями земель и 28% всех ельников Подмосковья. Такие катастрофические последствия для лесов, выполняющих защитные функции, заставляют задуматься о необходимости повышения устойчивости лесов Московской обл. Прежде всего это касается создания и формирования новых насаждений на лесных участках, пройденных сплошными санитарными рубками.
Л. П. Рысин для условий Московской обл. считал ель коренной породой [1]. Очевидно, что несмотря на циклически повторяющиеся вспышки короеда-типографа и засухи, ель будет оставаться одной из основных лесообразующих пород подмосковных лесов. Это обусловлено ее теневыносливостью и долговечностью, благодаря чему она способна занимать доминирующее положение в насаждениях через смену пород.
Одним из постулатов в современной экологии является представление о меньшей устойчивости экосистем с пониженным биологическим разнообразием [2]. Увеличение биоразнообразия экосистемы повышает ее устойчивость, что связано с различной реакцией разных организмов на внешнее воздействие климатических, биотических и антропогенных факторов. В нашем случае из этого постулата следует, что создавать чистые ельники, особенно в защитных лесах, нецелесообразно. Однако при формировании смешанных древостоев необходимо подбирать породы, которые по ряду позиций дополняют друг друга, что компенсирует воздействие тех или иных экологических факторов. Ель очень требовательна к влагообеспеченности. В сухой период у древесных пород с глубокой корневой системой процесс транспирации нарушается меньше (или совсем не нарушается), чем у пород с поверхностной корневой системой; и они легче переносят засуху. Например, даже в зоне тайги ель, которая имеет поверхностную корневую систему и предпочитает влажный климат и влажные почвы, погибает в крайне засушливые годы или сразу после них [3]. Соответственно порода-спутник ели должна иметь природные механизмы защиты от засушливых периодов, аналогичные ели темпы роста для условий Московской обл., обладать теневыносливостью, чтобы переносить конкуренцию с елью или временное затенение породами-пионерами и травянистой растительностью на вырубках.
Исходя из литературных источников, такой породой в зоне хвойно-широколиственных лесов является липа. Согласно шкале М. К. Турского липа занимает одно из последних мест по требовательности к свету, уступая только пихте, буку, ели и грабу [4]. По Е. С. Мурахтанову, липа имеет глубокий стержневой и сильно развитые боковые корни, поэтому эта порода устойчива к ветровалу. Благодаря глубокой и хорошо развитой корневой системе и наличию в листьях малого количества устьиц липа легко переносит периодические засухи, а при очень сильных засухах только снижает прирост. При этом на плодородных почвах она более устойчива к засухе. Кроме того, липа ценится как почвоулучшающая порода. Это качество определяет ее преимущество перед другими породами [5].
Исследования Н. П. Ремезова (1959) по распределению корневой системы в почвенном профиле показали, что липа осваивает верхний (до 0,5 м) слой почвенной толщи, но часть корней (20–30%) доходит до суглинистой почвы. Такое строение корневой системы липы обеспечивает не только ее высокую устойчивость, но и способность в полной мере использовать ресурсы почвы – влагу и элементы питания [1, 6]. К аналогичным выводам пришел в своих исследованиях Ю. П. Хлонов [7, 8].
М. И. Гордиенко отмечает, что в засушливый период листья липы желтеют и опадают раньше, чем у других пород, уменьшая общий расход влаги из почвы [9, 10].
Проблема межвидовой и внутривидовой конкуренции как фактора, определяющего рост и развитие древостоев, рассматривалась в лите- ратуре неоднократно [3, 11]. В частности, изучался такой аспект данной проблемы, как влияние этого фактора на формирование радиального прироста древесины [12, 13].
Цель данной работы – оценка реакции ели европейской в древостоях разного породного состава на воздействие различных климатических факторов с помощью методов анализа дендроклима-тической информации. Исследования проведены в 2016 г. на территории Щелковского учебноопытного лесхоза Мытищинского филиала МГТУ им. Н. Э. Баумана (ЩУОЛХ). С каждого учетного дерева на высоте 1,3 м отбирали по одному керну древесины с помощью бурава Преслера. Число кернов, отобранных с каждой постоянной пробной площади (ППП), варьировалось от 7 до 13. Таксационная характеристика насаждений на постоянных пробных площадях приведена в табл. 1.
В дендрохронологической лаборатории Научно-образовательного экспертно-аналитического центра исследований древесных растений Мытищинского филиала МГТУ им. Н. Э. Баумана с помощью прибора Lintab и специализирован-
Таблица 1. Таксационная характеристика насаждений на пробных площадях ЩУОЛХ
|
№ |
Год перечета |
Породный состав |
Средние |
Сумма площадей |
Пол- |
Возраст, лет |
Запас, |
Класс бонитета |
Подрост, тыс. шт./га (2013–2016 гг.) |
||
|
ППП |
по ярусам |
диаметр, см |
высота, м |
сечения, м2/га |
нота |
м3/га |
|||||
|
10 |
2013 |
8Е1Ос1Б+Д |
19,2 |
15,5 |
23,3 |
0,54 |
58 |
181 |
II |
5Д4Ос1Лп (2,66) |
|
|
2016 |
8Е1Ос1Б+Д |
20,2 |
16,9 |
24,7 |
0,54 |
61 |
206 |
II |
|||
|
100 |
2001 |
7Е1Ос1Б1Лп+С |
20,3 |
21,4 |
33,3 |
0,65 |
47 |
341 |
Ia |
9Лп1Д (4,29) |
|
|
2013 |
7Е1Ос1Б1Лп+С |
23,9 |
24,5 |
32,8 |
0,6 |
60 |
379 |
||||
|
106 |
2002 |
10Е |
14,2 |
15,2 |
40,6 |
0,96 |
44 |
310 |
I |
8Лп2Е+Д (2,67) |
|
|
2013 |
10Е |
17,1 |
15,8 |
52,2 |
1,0 |
55 |
412 |
II |
|||
|
119 |
2013 |
I |
5Лп3Ос1Б1Е |
26,1 |
25,0 |
32,5 |
0,9 |
55–65 |
364 |
Ia |
7Лп2Е1Ос+Д (5,7) |
|
II |
9Лп1Е+Б |
11,6 |
16,7 |
4,2 |
0,1 |
40–45 |
33 |
||||
|
120 |
2013 |
I |
9Е1Б+Ос |
21,5 |
20,4 |
25,9 |
0,72 |
127 |
242 |
III |
6Д4Е (0,65) |
|
II |
8Е2Б+Д,Ос |
12,7 |
16,5 |
10,0 |
0,29 |
78 |
|||||
|
2016 |
I |
9Е1Б+Ос |
22,4 |
23,1 |
29,2 |
0,66 |
130 |
273 |
III |
||
|
II |
8Е1Б1Д+Ос |
12,7 |
18,3 |
10,5 |
0,27 |
80 |
|||||
|
121 |
2013 |
9Е1Б+Д |
19,8 |
15,4 |
38,6 |
0,89 |
55 |
298 |
II |
10Д (0,12) |
|
|
2016 |
9Е1Б+Д |
21,1 |
17,7 |
38,0 |
0,82 |
58 |
330 |
||||
|
122 |
2013 |
I |
5Ос3Лп1Б1Е |
24,5 |
23,0 |
35,6 |
1,0 |
60–75 (122*) |
370 |
I |
9Лп1Е (7,31) |
|
II |
7Лп3Е |
12,7 |
14,0 |
7,7 |
0,3 |
35–50 |
52 |
||||
|
123 |
2014 |
6Лп3Е1Б + С, Д |
32,2 |
28,7 |
40,3 |
1,0 |
75–90 |
511 |
Ia |
9Лп1Е+Д, Кл(4,25) |
|
|
127 |
2014 |
I |
7Лп2Е1Б+Д |
23,6 |
25,0 |
29,5 |
0,9 |
65–70 (125*) |
330 |
Ia |
7Е3Лп+Д (5,74) |
|
II |
8Лп2Е+Б, Д |
10,4 |
13,4 |
3,8 |
0,1 |
30–45 |
25 |
||||
|
129 |
2014 |
I |
7Б3Лп+Ос, Е |
16,1 |
23,3 |
12,7 |
0,5 |
30–35(65*) |
134 |
Ia |
10Лп+Ос, Д, Е (4,14) |
|
II |
5Лп5Б+Ос, Е |
9,6 |
16,1 |
3,7 |
0,1 |
20–35 |
28 |
||||
|
131 |
2014 |
I |
5С5Б+Е, Лп |
26,1 |
27,3 |
31,3 |
0,7 |
65–80 |
379 |
Ia |
9Лп1Е+Д (6,8) |
|
II |
5Е4Лп1Б+С, Кл, Д |
11,2 |
13,7 |
8,3 |
0,3 |
30–55 |
55 |
||||
|
135 |
2015 |
4Лп3Е3Б+Д, Ос |
20,1 |
18,7 |
37,5 |
1,0 |
75 |
342 |
Ia |
5Лп3Е1Б 1Ос+Д (3,83) |
|
|
140 |
2015 |
9Е1Б+Д |
21,4 |
24,8 |
45,7 |
0,83 |
66 |
534 |
I |
10Д (0,23) |
|
* Возраст единичных деревьев ной программы Tsap-win были построены индивидуальные древесно-кольцевые хронологии для каждого учетного дерева. Затем эти хронологии индексировали путем деления годичного радиального прироста на средний радиальный прирост за последние 5 лет. На основе индексированных хронологий построили усредненные хронологии для каждой пробной площади. Далее отобрали пробные площади в двух группах древостоев: с участием ели в составе древостоя более 6 единиц и с участием липы более 6 единиц. Сравнение древостоев с преобладанием липы и ели неслучайно. Благодаря способности произрастать под пологом ельников и обильному порослевому возобновлению, быстрому росту, устойчивости к заморозкам и болезням, а также теневыносливости липа в благоприятных для нее почвенных условиях становится серьезным конкурентом хвойных и лиственных пород. При этом в хвойно-широколиственных лесах, участвуя в первом ярусе, липа является главнейшим компонентом леса и почти повсеместно достигает размеров дерева первой величины [5].
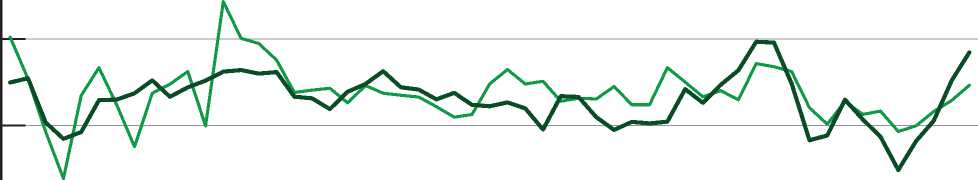
Графики, отражающие усредненные хронологии ели для групп древостоев с участием в их составе ели более 6 единиц и с участием липы более 6 единиц, представлены на рис. 1.
Хронологии характеризуются средней корреляцией (коэффициент корреляции равен 0,54), что свидетельствует о влиянии состава древостоя на характер изменчивости радиального прироста под действием климатических факторов. Для выявления конкретных форм модификаций проведен дендроклиматический анализ.
Для расчета коэффициентов корреляции между индексами прироста и рядами метеопараметров использованы данные метеостанций МГУ. Результаты расчета приведены в табл. 2. Достоверные значения коэффициентов корреляции выделены в таблицах жирным шрифтом. При числе степеней свободы 53 и уровне доверительной вероятности 0,05 достоверными считаются значения коэффициента корреляции от 0,27 и выше [14].
Расчет коэффициента корреляции необходимо осуществлять прежде всего между временными рядами метеопараметров (сумма осадков по месяцам, средняя температура по месяцам), сопоставленными с рядами индексов радиального прироста по принципу «год в год». Такое сопоставление дает представление о влиянии метеоусловий текущего вегетационного периода на формирование прироста древесины. Кроме того, необходим анализ влияния на прирост метеоусловий, предшествовавших началу вегетационного периода, а также метеоусловий прошлого вегетационного периода [15, 16]. Это достигается расчетом коэффициента корреляции между сдвину-
Индекс прироста
1,5
1,2
мт41пчмо»опмтяп>ок«0№Опмт.Т1гиекео»опмт»1пчмо№0 ююююююююм».м».м».м».м».ооооооооооооооооооооохохохохохохохохохохе ^^ ^N ^N ^N ^N ^N ^N ^N ^N ^N ^N ^N ^N ^N ^N ^N ^N ^N O^ O^ O^ O^ O^ O^ O^ O^ O^ O^ ^^ ^N ^N ^N ^N ^N ^N ^N ^N ^N C1
0,6
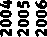
Год
Хронология ели в древостоях с участием липы более 6 единиц в породном составе
Хронология ели в древостоях с участием ели более 6 единиц в породном составе
Рис.
-
1. Динамика индексов радиального прироста ели по годам в насаждениях разного породного состава
тыми на один год относительно друг друга временными рядами.
Как видно из данных табл. 2, между приростом ели и отдельными метеопараметрами фиксируется некоторая слабая флуктурирующая связь: максимальные значения коэффициентов корреляции не превышают 0,38. Такая же связь установлена при сопоставлении чистого древостоя ели 10Е+С, Б и смешанного с участием 2 единиц ели в составе (4С2Е2Б), которое было выполнено в процессе аналогичного исследования на территории ЩУОЛХ [17]. Авторы объясняли с точки зрения классических представлений экологии эту связь тем, что Московская обл. лежит в той части ареала ели европейской, где ежегодные дозы климатических факторов близки дозам, составляющим зону оптимума жизнедеятельности организма [17]. Сильные отклонения от этих доз в большую или меньшую сторону отрицательно сказываются на продуктивности ели.
Наличие одинаковой реакции прироста на разнонаправленные экстремальные отклонения дозы факторов от среднего в совокупности со сменой лимитирующего фактора из года в год затрудняет использование корреляционного анализа при выявлении зависимостей продуктивности ели от воздействий экологических факторов. Тем не менее даже в таких условиях нередко фиксируются достоверные корреляции между изменчивостью радиального прироста и отдельных метеопараметров по годам. Так, в нашей работе, как и в предыдущем исследовании [17], зафиксирована достоверная отрицательная корреляция между радиальным приростом ели и увеличением среднемесячной температуры в июне текущего года в насаждениях с участием ели более 6 единиц в составе (значения коэффициентов корреляции составляют -0,38 и -0,33 соответственно), а также в августе в год, предшествовавший году формирования годичного кольца, в насаждениях c участием ели в составе 3 единицы и менее (-0,37 и -0,38 соответственно). Фенологически август соответствует периоду заложения вегетативных и генеративных почек. Вероятнее всего высокие температу-
Таблица 2. Результаты расчета коэффициентов корреляции индексов прироста с метеопараметрами текущего (числитель) и прошлого (знаменатель) годов за 1961–2016 гг.
|
Месяц |
Хронология ели в насаждениях с участием |
|
|
ели более 6 единиц в составе |
липы более 6 единиц в составе |
|
Осадки
|
Январь |
0,14/0,17 |
0,03/0,12 |
|
Февраль |
-0,08/-0,10 |
0,00/0,18 |
|
Март |
-0,28/-0,20 |
0,04/0,07 |
|
Апрель |
0,12/0,13 |
-0,08/0,15 |
|
Май |
0,08/0,19 |
0,10/0,18 |
|
Июнь |
0,23/0,07 |
0,04/0,14 |
|
Июль |
0,20/0,24 |
0,10/0,30 |
|
Август |
-0,09/0,02 |
-0,04/0,22 |
|
Сентябрь |
-0,16/-0,05 |
-0,31/-0,09 |
|
Октябрь |
-0,08/-0,03 |
0,06/0,09 |
|
Ноябрь |
0,03/-0,05 |
0,09/0,18 |
|
Декабрь |
0,03/0,06 |
-0,09/0,22 |
Температура
-
Х. Хеттонен [18] предложила другой вариант механизма редукции годичного кольца. Ссылаясь на ряд малодоступных в России источников, автор отмечает, что высокие летние температуры в год, предшествовавший году формирования
годичного кольца, определяют заложение у ели большого числа генеративных почек, что ведет к снижению прироста на следующий год за счет перевода значительного числа продуктов ассимиляции на формирование семян.
Следует отметить положительное влияние повышенных температур февраля и марта на формирование прироста древесины в древостоях с небольшой долей ели в составе. По-видимому, в смешанных древостоях повышенные температуры могут способствовать более быстрому таянию снега, скорейшему прогреванию почвы и раннему началу вегетационного периода [19].
Рассмотрим динамику индексов радиального прироста ели за последние 15 лет (в исследуемых хронологиях) (рис. 2). Выбранный период соответствует периоду постоянных наблюдений на исследуемых пробных площадях.
В течение рассматриваемого периода отчетливо выраженная засуха наблюдалась в 2010 г. Анализ влияния этой засухи на прирост ели в древостоях, резко отличающихся по породному составу, представляет интерес в связи с ее максимальной приближенностью по времени к периоду выполнения полевых таксационных исследований. Это важно, потому что состав древостоя – динамический показатель, на который большое влияние оказывает временной фактор.
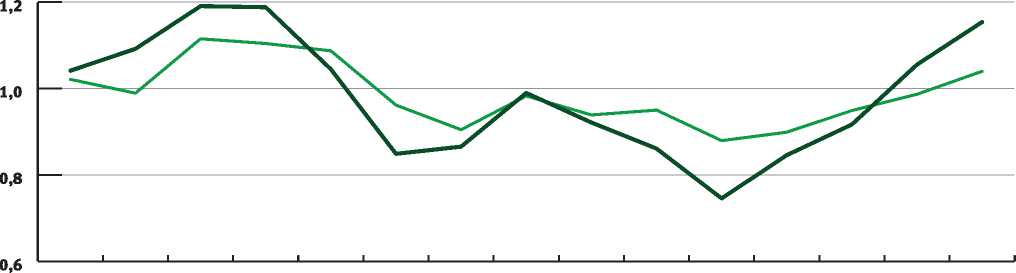
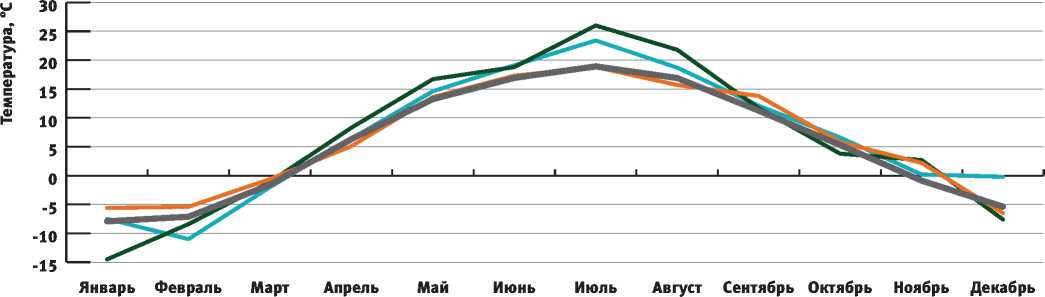
Рассмотрим подробнее погодный режим 2010 г. и сравним его со средними многолетними показателями, а также погодными условиями 2009 и 2011 гг. Данный анализ может быть выполнен методом климаграмм. Климаграммы, отражающие специфику распределения осадков и среднемесячных температур по месяцам в 2009, 2010 и 2011 гг. по сравнению со средними многолетними данными, приведены на рис. 3 и 4.
На рис. 3 и 4 видно, что 2010 г. характеризуется засушливым погодным режимом. В июле количество осадков было на 86,1% ниже средней многолетней нормы, а температуры превышали средние многолетние показатели на 37,5%.
В 2010 г. индекс радиального прироста ели для рассматриваемых групп был одинаковым (рис. 2), однако в 2011 г. этот показатель в группе «чистые ельники» снизился заметно сильнее, чем в древостоях с преобладанием липы. Феномен запаздывания реакции прироста ели на засуху хорошо известен и многократно описан [17]. В 2012–2014 гг. индекс радиального прироста ели в древостоях с преобладанием липы продолжал превышать аналогичный показатель для группы «чистые ельники». Из этого следует, что засуха 2010 г. оказала более сильное отрицательное влияние на рост ели в чистых еловых древостоях по сравнению с древостоями ели с преобладанием липы в составе.
Таким образом, и методом корреляционного анализа, и методом сопряженного анализа хронологий нам удалось выявить меньшую засухоустой-
Индекс прироста
2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 2014 2015 2016
Год
Хронология ели в древостоях с участием липы более 6 единиц в породном составе
Хронология ели в древостоях с участием ели более 6 единиц в породном составе
2. Динамика индексов радиального прироста ели в насаждениях разного породного состава за последние 15 лет
Рис.
2011 2009
2010 Среднее значение
Рис. 3. Сумма осадков по месяцам в 2009, 2010 и 2011 гг. в сравнении со средними многолетними значениями
Среднее значение
Рис. 4. Среднемесячные температуры в 2009, 2010 и 2011 гг. в сравнении со средними многолетними значениями
чивость деревьев ели в чистом древостое по сравнению с древостоями с небольшой долей ели в составе. То есть величина прироста ели в составе насаждения с преобладанием липы оказалась менее зависима от влияния климатических факторов, обусловливающих формирование водного дефицита в организме растений.
На основании вышеизложенного считаем возможным рекомендовать создание смешанных липово-еловых древостоев для Московской обл. в соответствующих лесорастительных условиях. Создание таких насаждений позволит повысить устойчивость лесов Подмосковья и обеспечит лучшее выполнение ими защитных функций.
Список литературы Оценка реакции ели европейской на воздействие климатических факторов в древостоях разного породного состава
- Рысин, Л. П. Леса Восточного Подмосковья: моногр./Л. П. Рысин; отв. ред. С. Ф. Курнаев. -М.: Наука,1979. -184 с.
- Коротков, С. А. Теоретические проблемы устойчивости леса/С. А. Коротков//Лесной вестник. -Т. 19. -2015. -№ 4. -С. 26-32.
- Мелехов, И. С. Лесоведение: учеб. для вузов/И. С. Мелехов. -М.: Лесн. пром-сть, 1980. -408 с.
- Турский, М. К. Лесоводство: науч. изд.: изд. 10-е/М. К. Турский. -М.: МГУЛ, 2010. -425 с.
- Мурахтанов, Е. С. Липа/Е. С. Мурахтанов. -М.: Лесн. пром-сть, 1981. -80 с.
- Ремезов, Н. П. Потребление и круговорот азота и зольных элементов в лесах Европейской части СССР/Н. П. Ремезов, Л. Н. Быкова, К. М. Смирнова. -М.: изд-во МГУ, 1959. -284 с.
- Хлонов, Ю. П. Липы и липняки Западной Сибири/Ю. П. Хлонов. -Новосибирск: СО АН СССР, 1965. -155 с.
- Рысин, Л. П. Липовые леса Русской равнины/Л. П. Рысин. -М.: Тов-во науч. изд. КМК, 2012. -195 с.
- Гордиенко, М. И. Взаимодействие дуба черешчатого и липы мелколистной/М. И. Гордиенко//Науч. тр. Укр. с.-х. акад. -Вып. 94. -1973. -С. 27-31.
- Гордиенко, М. И. Лесоводственная оценка липы мелколистной, клена остролистного и граба обыкновенного/М. И. Гордиенко//Лесоведение. -1979. -№ 1. -С. 59-66.
- Сукачев, В. Н. Растительные сообщества (введение в фитосоциологию): 4-е изд./В. Н. Сукачев. -М.-Л.: Книга, 1928. -232 с.
- Крамер, П. Д. Физиология древесных растений/П. Д. Крамер, Т. Т. Козловский. -М.: Лесн. пром-сть, 1983. -464 с.
- Усманов, И. Ю. Экологическая физиология растений/И. Ю. Усманов, З. Ф. Рахманкулова, А. Ю. Кулагин. -М.: Логос, 2001. -224 с.
- Лакин, Г. Ф. Биометрия: учеб. пособ. для универ. и педагог. институтов/Г. Ф. Лакин. -М.: Высшая школа, 1973. -343 с.
- Fritts, H. C. Tree rings and climate/Н. С. Fritts. -London-New-York, San Francisco: Academic press, 1976. -576 p.
- Засухи Восточно-Европейской равнины по гидрометеорологическим и дендрохронологическим данным/под ред. О. Н. Соломиной. -С.-Пб.: Нестор-История, 2017. -360 с.
- Дендрохронологическая информация в лесоводственных исследованиях: моногр./под ред. В. А. Липаткина, Д. Е. Румянцева. -М.: МГУЛ, 2007. -137 с.
- Hettonen, H. The dependence of annual rings indices on some climatic factors/H. Hettonen//Actaforestaliafennica. -1984. -Vol. 186. -38 p.
- Бех, И. А. Лес и жизнь: моногр./И. А. Бех, А. М. Калинин, И. В. Таран. -Кемерово: Книжное изд-во, 1986. -160 с.
