Оценка состояния ценопопуляций очитков (Crassulaceae DC.) на Севере
 на Севере")
Автор: Бабак Т.В.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 2, 2010 года.
Бесплатный доступ
Обобщены итоги комплексных исследований двух представителей семейства Crassulaceae DC. на северной границе ареала в пределах таежной зоны Республики Коми. Впервые даны популяционные характеристики изучаемых видов в условиях европейского Северо-Востока и оценка состояния ценопопуляций. Приведены экологические характеристики местообитаний видов.
Состояние ценопопуляций, граница распространения, популяционные характеристики
Короткий адрес: https://sciup.org/14992382
IDR: 14992382 | УДК: 582.711.16
Estimation of state of Sedum L. (Crassulaceae DC.) coenopopulations in the North
The results of complex investigations of Sedum acre L. and Hylotelephium triphyllum (Haw) Holub., two species from Crassulaceae DC. family in the northern boundary of its distribution within taiga subzone of the Komi Republic, are generalized. Original material on population characteristics (age structure, seed production, population density and number) are obtained. Data on modern state of studied species of coenopopulations are considered. The basic age spec-trum and ecological characteristics of these species habitats are presented.
Текст научной статьи Оценка состояния ценопопуляций очитков (Crassulaceae DC.) на Севере
Семейство Crassulaceae DC. (толстянковые) представляет большой интерес с ботанической, физиологической и хозяйственной точки зрения. Виды этого семейства широко распространены по всему Земному шару, преимущественно в сухих теплых и умеренных областях. В пределах бывшего СССР встречается, по разным данным, от 9 до 13 родов, включающих от 121 до 148 видов [1, 2], из которых на территории европейского Северо-Востока России представлены только три рода: Rhodiola L., Sedum L. и Hylotelephium H. Ohba. К настоящему времени наиболее обобщены данные по роду Rhodiola [3-10]. Два других рода: Sedum и Hylotelephium остаются практически неизученными. Имеющиеся в литературе сведения по ним отрывочны, вопросам биоморфологии и биогеографии посвящено весьма незначительное число работ. Более полно представлены данные по систематике, таксономии и биохимии этой группы растений. Большая часть работ носит прикладной характер, что связано с перспективным использованием очитков как лекарственного сырья и в качестве декоративных растений. В настоящее время интерес представляет изучение свойств ценопопуляций различных видов растений и их сравнение в разных частях ареала, особое внимание привлекают подобные исследования на границе распространения вида.
Цель нашей работы – выявить приспособления природных ценопопуляций Sedum acre L. и Hy-lotelephium triphyllum (Haw.) Holub к длительному, устойчивому существованию в условиях Севера и оценить их современное состояние.
Материал и методы
Объектами исследований были два представителя семейства Crassulaceae – S. acre (очиток едкий) и H. triphyllum (очиток или очитник пурпурный, ранее S. purpureum L.) из природных ценопо-пуляций. Изучение природных ценопопуляций S.acre и H.triphyllum проводили на территории европейского Северо-Востока в пойме бассейна р.Луза (граница подзон южной и средней тайги), на песчаных обнажениях вдоль шоссе Сыктывкар – Ухта (окрестности с.Серегово), в поймах бассейна рек Сысола, Вычегда (подзона средней тайги), верховьях р.Щугор (подзона северной тайги), низовьях р.Печора и устье рек Цильма и Печорская Пижма, а также в средней части р. Печорская Пижма (подзона крайнесеверной тайги). Всего за вегетационные периоды 2000 – 2004 гг. изучено 30 ценопопуляций H. triphyllum и 10 ценопопуляций S. acre в различных сообществах и экотопах.
При изучении ценопопуляций использовали общепринятые методики [11-13]. В растительных сообществах с H.triphyllum на трансектах закладывали учетные площадки 1х1 – 2х2 м2, для изучения S.acre – 0.25х0.25 м2. Учитывали следующие признаки: численность и плотность особей, возрастной состав, способ возобновления. Для каждой ценопопу-ляции был установлен возрастной спектр. Возрастную структуру ценопопуляции определяли на основе учета численности особей по возрастным состояниям. На основании возрастных спектров ценопопуля-ций рассчитаны базовые возрастные спектры двух модельных видов, произрастающих на территории европейского Северо-Востока.
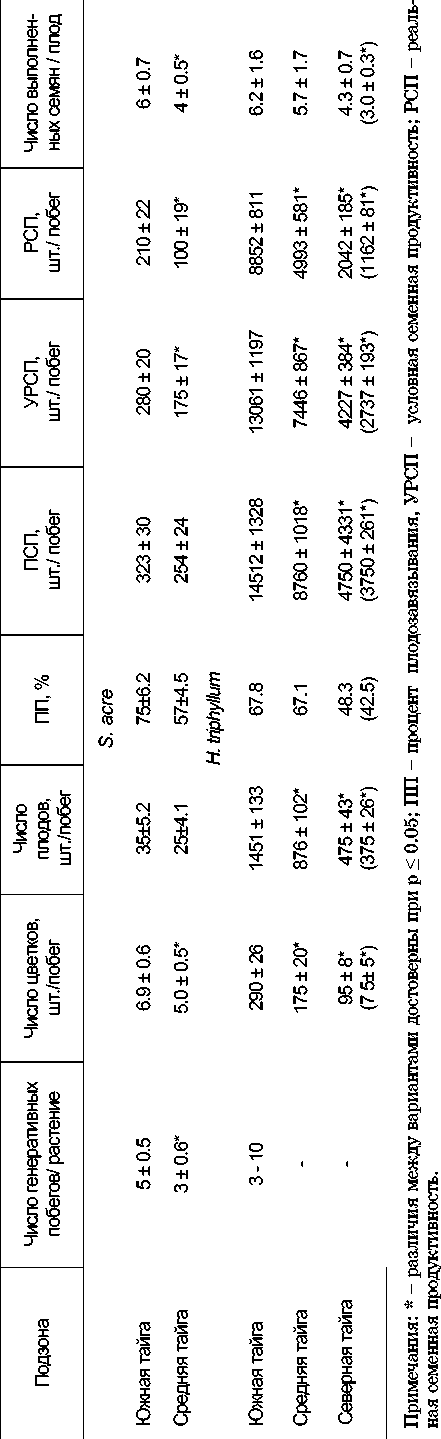
При определении семенной продуктивности в ценопопуляциях был проведен расчет числа генеративных побегов на особь, числа цветков, плодов, семязачатков и выполненных семян на побег, а также рассчитаны величины потенциальной семенной продуктивности (ПСП), реальной семенной продуктивности (РСП), процент плодозавязывания [14, 15], проведен подсчет условно-реальной семенной продуктивности (УРСП) [16].
Описание растительных сообществ выполнено по методике Л.Г.Раменского [17]. Список видов уточнен по сводке [2]. С помощью фитоиндикаци-онных экологических шкал [18, 19] определены экологические особенности и региональные амплитуды изучаемых видов на исследованной территории.
Результаты и обсуждение
На территории европейского Северо-Востока проходит крайняя северо-восточная граница распространения средиземноморско-европейского вида – S.acre . Это сукку-лентнолистовое летнезимнезеленое, многолетнее травянистое растение. По классификации жизненных форм растений К. Раункиера – хамефит. Имеет двухтипные укореняющиеся ползучие и приподнимающиеся неодревесневающие побеги, которые при разрастании образуют куртины или «клоны-латки».
Большинство ценопопуляций S.acre, находящихся на территории европейского Северо-Востока, имеют неполночленные возрастные спектры. Для всех спектров характерно отсутствие проростков, в некоторых отсутствуют генеративные и сенильные растения, в большинстве ценопопуляций доминирует имматурная возрастная группа, ее доля составляет от 35 до 42%. По данным исследований ценопопуляций S.acre в южной и средней частях подзоны средней тайги был составлен базовый возрастной спектр (рис. 1), отражающий общие закономерности развития вида на границе распространения под влиянием эколого-географических факторов. Выявлено, что доминирование имма-турной (im) и виргинильной (v) возрастных групп обусловлено преобладанием вегетативного размножения, а также замедленной сменой поколений растений семенного происхождения в неблагоприятных для развития условиях.
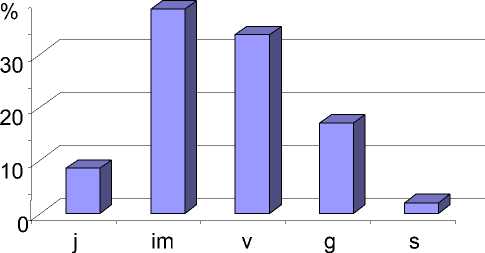
Рис. 1. Базовый возрастной спектр S. acre на северо-восточной границе распространения.
При сравнении ценопопуляции S.acre в зональном плане (табл. 1) выявлено, что количество особей и рамет S.acre в южной части среднетаежной подзоны в три раза, а количество побегов ветвления в четыре раза больше, чем в средней. По направлению на север наблюдается также уменьшение средней плотности ценопопуляции в 2.6 раз и изменение структуры ценопопуляции: от сплошного размещения особей – моногруппировок – до ценопопуляций, представленных отдельными небольшими куртинами.
Исследования показали, что растения S.acre развивали в среднем от трех (в центральной части подзоны средней тайги) до пяти генеративных побегов на растение (в южной части)(табл. 2). При этом
Таблица 1
Характеристика ценопопуляций Sedum acre в подзонах южной (1) и средней тайги (2)
|
Показатель |
1 |
] 2 |
|
Число особей, шт / ЦП |
836 ± 26 |
279 ± 10 * |
|
Число побегов, шт / ЦП |
5933 ± 245 |
1520 ± 68 * |
|
Площадь ЦП, кв.м |
10 ± 1.6 |
10.1 ± 0.07 |
|
Плотность ЦП, шт./кв.м: |
||
|
-средняя |
55.7 ± 3.9 |
21.5 ± 2.5 * |
|
-экологическая |
6 2 ± 8.5 |
44 ± 3.4 |
|
ПП, % |
78 |
31.5 |
Примечания: * – различия между вариантами достоверны при Р ≤ 0,05; ЦП – ценопопуляция, ПП – проективное покрытие.
число цветков и плодов на генеративный побег с юга на север уменьшалось в 1.4 раза. Показатель УРСП для S.acre варьировал от 175 (в средней части) до 280 шт./побег (в южной части); РСП – соответственно от 100 до 210 шт./побег. Для установления качества семян нами была определена их выполненность. Такие семена составляли от 57 до 75% от общего числа семян, при этом в средней части подзоны их процент был в 1.3 раза меньше, чем в южной.
Основную роль в размножении растений S.acre играет – вегетативное, которое осуществляется путем образования рамет, плагиотропных вегетативных побегов (длиной до 10 см). Появление новых особей у S.acre происходит, главным образом, за счет повреждения материнских растений и обособления дочерних рамет. Приоритет вегетативного размножения S.acre на границе ареала мож-
Таблица 2
но объяснить несколькими причинами: во-первых – повышенной жизнестойкостью отделенных побегов за счет их суккулент-ности и способностью в короткий промежуток времени при благоприятных условиях образовывать придаточные корни, во-вторых – большей конкурентоспособностью и выживаемостью по сравнению со всходами, и в-третьих, постепенным захватом территории – превращением отдельных куртин в моногруппировки за счет активного роста парциальных побегов и интенсивного образования партикул. Согласно нашим наблюдениям и исследованиям [20], скорость разрастания клонов связана не только с длиной многолетнего участка плагиотропной части материнских побегов, но и с отхождением от него побегов ветвления и образованием отделившихся дочерних особей.
S.acre – стресс-толерантный вид [21], приуроченный к сообществам с разреженным напочвенным покровом, в сложении которых значительное участие принимают однолетники, стержнекорневые и реже дерновинные травянистые многолетники. Типичными местами произрастания на обследуемой территории являются суходольные луга, окраины сухих сосновых боров и населенные пункты. В соответствующих экологических условиях участвует в зарастании оголенных участков, но по мере формирования сомкнутого растительного покрова постепенно исчезает. Часто поселяется в нарушенных человеком местообитаниях, вдоль дорог, на паровых полях и сбитых сухих лугах. Однако слабо устойчив к вытаптыванию [19].
Применение метода фитоиндикаци-онных экологических шкал позволило нам дать общую экологическую характеристику местообитаний S.acre . Для наглядного изображения экологических ареалов изучаемого вида нами была использована графическая модель – полиграмма с диапазонами экологических шкал [18] (рис.2). Анализ местообитаний S.acre на территории Республики Коми показал, что наименьшие доли экологического пространства в экологическом ареале вида наблюдаются по общему терморежиму, континентальности и морозности климата, а также увлажнению и богатству почв (составляют от 22 до 30%). По влажности климата и кислотности почв размеры занимаемого видом экологического пространства приближаются к 50%, а по переменности увлажнения почв и по режиму затенения доли экологического пространства составляют 60 и 100% от экологического ареала вида.
Второй изученный нами вид – H.tri-phyllum является евроазиатским бореальным, имеет достаточно широкое распространение по территории европейского Се-веро-Востока.Это сукклентнолистовой ве- сенне-летне-осеннезеле-ный травянистый многолетник с прямостоячими побегами; относится к без-розеточным криптофитам, корнеклубневым геофитам.
Общая характеристика ценопопуляции H.tri-phyllum на территории европейского Северо-Востока показала, что возрастные спектры данного вида представлены как полночленными (характерно для ценопопуляций, находящихся преимущественно вблизи леса), так и факультативно неполночленными. Отсутствие одной или нескольких возрастных групп в спектрах, вероятно, было обусловлено особенностями онтогенеза вида в районе исследований, либо нерегулярностью семенного размножения.
Обобщение результатов анализа возрастных спектров H.triphyllum по обследованным ценозам и экотопам позволило выделить четыре типа онтогенетического спектра (рис. 3): нормальный полночленный и неполночленный зрелый – на пойменных разнотравнозлаковых лугах в ассоциации с пойменными кустарниками, где абсолютный максимум приходится на средневозрастные генеративные особи (1); инвазионно-регрессивный – под пологом леса и в зарослях крупного ивняка, где максимум приходится либо на взрослые генеративные и сенильные (2), либо на ювенильные и имматурные группы (3) и инвазионный – на обрывистых берегах и в оврагах, где преобладают виргинильные особи (4). В луговых ценозах с наиболее типичным для H.triphyllum местообитанием, как правило, это окраина луга вблизи зарослей кустарников (ива или шиповник), средних и максимальных значений в возрастных спектрах ценопопуляций достигали средневозрастные генеративные особи. Анализ возрастных спектров H.triphyllum в зональном градиенте, где было проведено сравнение состояния модельных популяций из трех разных точек ареала в пределах Республики Коми, показал, что в двух популяциях (южная и средняя точки) происходило активное семенное возобновление: ювенильные особи в них составляли до 14%, доля генеративных – до 37%.
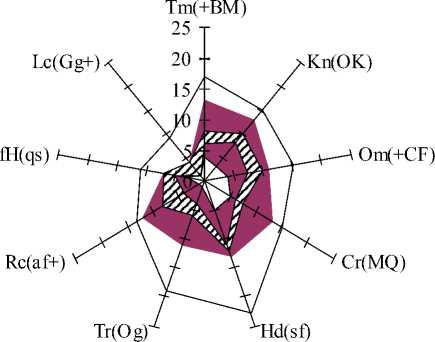
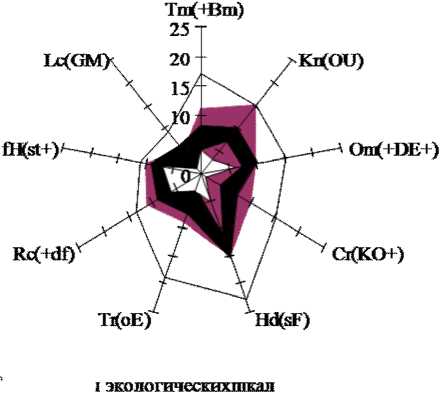
□ диапазоны
■ max - min границы экологических ареалов вида
■ max - min границы на изучаемой территории
Рис. 2. Экологические факторы в ценопопуляциях S. acre (А) и H. triphyllum (Б) (согласно экологическим шкалам Д.Н. Цыганова, 1983). Примечания : Tm – терморежим; Kn – континентальность; Om – омбро-режим; Cr – криорежим; Hd – гидрорежим; Tr – трофность почв; Rc – кислотность почв; fH – переменность увлажнения; Lc – освещенность в ценозе.
Третий спектр (северная точка) отличался от предыдущих доминированием виргинильных и генеративных партикул и малой долей молодых растений.
По мере продвижения на север общее число растений и побегов в це-нопопуляциях H.tri-phyllum значительно уменьшалось (табл.3), при этом количество побегов на растении достоверно не изменялось на всем протяжении от южной до северной точки распространения; происходило сокращение площади цено-популяций в 1.5 – 3 раза, что, вероятно, послужило причиной снижения общей численности данного вида. Достоверное изменение числа побегов (шт./м2) было отмечено при сравнении южной части средней и северной подзоны тайги, а также подзон северной и крайнесеверной тайги. Изменения числа растений (шт./м2) варьировали в пределах ошибки.
Растения H.triphyl-lum имели в среднем от трех до десяти генеративных побегов (табл.2). Величина этого показателя не зависела от зональной локализации местообитания. Число цветков и плодов на генеративном побеге в природных ценопопуляциях с юга на север снизилось в три-четыре раза, происходило также уменьшение диаметра и разветвленности соцветия. Величина УРСП для данного вида составляла от 2.7 тыс. семян на побег (в подзоне северной тайги) до 13.1 тыс. шт./ побег (в южной части среднетаежной подзоны). Величина РСП изменялась от 1.2 тыс. (подзона северной тайги) до 8.8 тыс. шт./ побег (южная граница средней подзоны). Доля выполненных семян варьировала от 42.5 до 67.8% от общего числа семян. При этом на крайней северной точке процент распространения их был в 1.6 раза меньше, чем в южной. Следовательно, по направлению на север (по подзонам) снижается потенциальная продуктивность на элемент в год, увели-
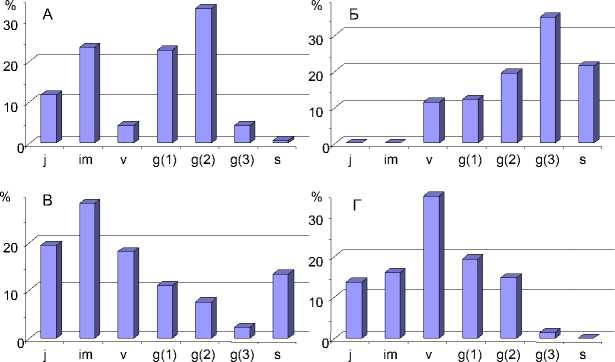
Рис. 3. Онтогенетические спектры, характерные для ценопопуля-ций H. triphyllum в разных растительных сообществах: А – нормальный полночленный и неполночленный зрелый (I тип); Б – инвазионно-регрессивный с доминированием среднегенеративных и сенильных групп (II тип); В – инвазионно-регрессивный с преобладанием ювенильных и имматурных групп (III тип); Г – инвазионный (IV тип).
бей вегетативного происхождения, а при слабой сомкнутости травостоя це-нопопуляции представлены в основном особями семенного происхождения.
Исследования ценопопуляций H.triphyllum в пределах таежной зоны европейского Северо-Востока и изучение гербарного материала показали, что вид имеет широкую экологическую амплитуду и достаточно обширно распространен по данной территории. При этом ценотическая приуроченность его несколько уже: основное число ценопопуляций H.tri-phyllum обнаружено по поймам рек, преимущественно в зарослях пойменных кустарников ( Salix sp. и Rosa acicularis Lindl. или R.majalis Herrm), где вид находит более благоприятные условия – хорошо прогреваемые места с умеренным затенением и увлажнением. Анализ местообитаний H.triphyllum на территории европейского Северо-Вос-
Таблица 3 тока показал, что наименьшие доли эколо-
|
Численность и плотность ценопопуляций Hylotelephium triphyllum |
||||||
|
Параметр |
Подзона южной тайги |
Подзона средней тайги |
Подзона северной тайги |
Подзона крайнесеверной тайги |
||
|
1 |
2 |
1 |
2 |
|||
|
Число побегов, шт./ЦП |
846 ± 28 |
530 ± 22* |
120 ± 9 |
264 ± 10* |
550 ± 21 |
84 ± 6* |
|
Число растений, шт./ЦП |
141 ± 22 |
90 ± 18 |
63 ± 3 |
88 ± 7* |
98 ± 6 |
28 ± 2* |
|
Число побегов, шт./ растение |
6.0 ± 2.1 |
5.9 ± 1.7 |
2.1 ± 0.7 |
3.0 ± 0.7 |
5.8 ± 1.1 |
3.0 ± 0.5 |
|
Число побегов, шт./ кв.м |
28.2 ± 4.1 |
21.2 ± 2.1 |
12 ± 1.2 |
13.2 ± 1.6* |
20.3 ± 3.1 |
8.4 ± 1.2* |
|
Число растений, шт./ кв.м |
6.0 ± 2.1 |
3.7 ± 1.0 |
6.7 ± 1.1 |
5.0 ± 0.9 |
4.0 ± 1.2 |
3.0 ± 0.7 |
|
Площадь ЦП, кв.м |
30 ± 4.4 |
25 ± 4.1 |
10 ± 2.1 |
20 ± 1.2* |
27 ± 1.6 |
10 ± 1.5* |
|
ОПП, % |
40 - 90 |
45 - 90 |
10 - 15 |
50 - 100 |
20 - 30 |
45 - 65 |
Примечания: * – различия между вариантами достоверно при Р ≤ 0,05; 1 – на пойменном лугу; 2 – под пологом леса; ОПП – общее проективное покрытие.
чивается число невыполненных семян и уменьшается процент плодозавязывания.
H.triphyllum относится к вегетативно малоподвижным видам, размножающимся, как правило, семенами. С продвижением на север наблюдается увеличение роли вегетативного размножения, происходящего за счет травматической партикуляции у среднегенеративных и стареющих особей. Согласно Т.Ю.Рогожиной [22], в культуре данный вид, как правило, размножают черенками либо искусственной партикуляцией. Отмечено также, что в ценозах с большим проективным покрытием и обострением конкурентных отношений увеличивается доля осо- гического пространства в экологическом ареале вида наблюдаются только по континентальности климата (составляют 31%) (рис.2); одновременно охват свыше 50% экологического ареала по таким факторам, как общий терморежим, влажность и морозность климата, а также по увлажнению почв свидетельствует о несколько меньшей стено-топности и большей патиентности вида к этим факторам. Однако значительную патиентность вид проявляет по отношению к богатству, кислотности и переменности увлажнения почв, а также к режиму затенения, где доли экологического пространства ценопопуляций на территории европейского Северо-Востока приближены либо превышают 100% от экологического ареала вида.
Заключение
Изучение состояния ценопопуляций очитков – S.acre и H.triphyllum , представителей семейства Crassulaceae на европейском Севере, обнаружило некоторые характерные для данных видов особенности, способствующие их существованию в условиях холодного климата.
Исследование эколого-ценотических характеристик S.acre на европейском Северо-Востоке выявило, что распространение данного вида в пределах изучаемой территории весьма ограничено. В средней подзоне тайги проходит северо-восточная граница его ареала, а основная часть ценопопуля-ций S.acre встречается в подзоне южной тайги и на границе южной и средней. Основу популяций S.acre в данных точках составляют парциальные побеги вегетативного происхождения. Все возрастные спектры изученных ценопопуляций S.acre являлись неполночленными, при этом в большинстве из них доминировала имматурная возрастная группа. Выявлено, что для S.acre характерно сплошное или групповое размещение растений по площади цено-популяций, что является следствием активного вегетативного размножения. При сравнении ценопо-пуляций S.acre в зональном плане показано, что по направлению с юга на север уменьшается плотность ценопопуляций и происходит изменение их структуры. На территории европейского Северо-Востока наблюдается снижение числа генеративных побегов, цветков и выполненных семян. Как следствие, на границе ареала в два раза сокращается реальная семенная продуктивность S.acre . В результате анализа эколого-ценотических условий местообитаний S.acre выявлено, что, несмотря на относительно широкие границы экологических факторов для вида в целом, в пределах изучаемой нами территории ценопопуляции S.acre по большей части экологических факторов встречаются лишь в узком диапазоне условий. Это определяет высокую степень мозаичности в распределении S.acre в экосистемах на границе ареала вида.
H.triphyllum достаточно широко распространен в пределах европейского Северо-Востока, характеризуется широкой экологической амплитудой и приурочен преимущественно к местам среднего уровня увлажнения и освещенности. H.triphyllum относится к экологической группе мезофитов, по оптимуму богатства почвы является мезотрофом. К условному экологическому оптимуму H.triphyllum наиболее близки пойменные луга средней подзоны тайги. Выявлено, что в возрастных спектрах ценопо-пуляций H.triphyllum доминируют молодые и генеративные растения. В северном направлении в цено-популяциях отмечено значительное уменьшение общего числа растений и побегов. В зависимости от типа местообитания и растительного сообщества выделено четыре вида онтогенетического спектра H.triphyllum . Отмечено, что для большей части цено-популяций H.triphyllum характерно преимущество семенного возобновления. С продвижением на север увеличивается доля вегетативного размножения за счет травматической партикуляции у средневозрастных и стареющих особей.
Список литературы Оценка состояния ценопопуляций очитков (Crassulaceae DC.) на Севере
- Борисова А.Г. Семейство Толстянковые -Crassulaceae DC.//Флора СССР. М.-Л.: Изд-во АН СССР, 1939. Т.9. С.8-134.
- Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья, 1995. 992 с.
- Фролов Ю.М., Полетаева И.И. Возрастной состав ценопопуляций родиолы розовой на Приполярном Урале. Сыктывкар, 1995. С.13-26. (Труды Коми НЦ УрО РАН; №141).
- Фролов Ю.М., Полетаева И.И. Специфика возрастного состава ценопопуляций родиолы розовой на Приполярном Урале в постсеменной год. Сыктывкар, 1996. С.128-139. (Труды Коми НЦ УрО РАН; №149).
- Фролов Ю.М., Полетаева И.И. Родиола розовая на европейском Северо-Востоке. Екатеринбург: УрО РАН, 1998. 192 с.
- Фролов Ю.М. Особенности возрастной структуры ценопопуляций Rhodiola rosea L. на юге Приполярого Урала в 1995 г. Сыктывкар, 2001. С. 50-61. (Труды Коми НЦ УрО РАН; №165).
- Лавриненко О.В., Сивков М.Д., Назаров С.К. СО2 -газообмен Rhodiola rosea L. в естественных условиях обитания и при интродукции в таежную зону Республики Коми//Дыхание растений: физиологические и экологические аспекты: Тезисы Международного совещания. Сыктывкар, 1995. С. 86-87.
- Лавриненко И.А., Ткаченко К.Г., Елсаков В.В. Популяционная и межвидовая изменчивость двух видов рода Rhodiola (Crassulaceae) в условиях Арктики//Ботанический журнал, 1998. Т. 83 №. 9. С.63-70.
- Далькэ И.В., Головко Т.К., Бачаров Д.С. Роль покоя почек в адаптации Rhodiola rosea L. в холодном климате//Материалы Всероссийской научно-практической конференции. Ярославль: Яросл. гос. ун-т, 2003. С.80-81.
- Морфофизиологические и биохимические характеристики Rhodiola rosea L. из разных ценопопуляций на европейском Севере/И.В.Далькэ, Т.К.Головко, И.Г.Захожий, Г.Н.Табаленкова//Методы популяционной биологии: Сборник материалов докладов VII Всероссийского популяционного семинара. 16-21 февраля 2004 г. (часть 1). Сыктывкар, 2004. С.47-49.
- Ценопопуляции растений (Основные понятия и структура)/Под ред. О.В.Смирновой, Л.Б. Заугольновой, И.М.Ермаковой и др. М.: Наука, 1976. 216 с.
- Ценопопуляции растений (Очерки популяционной биологии)/Под ред. Л.Б. Заугольновой, Л.А. Жуковой, А.С. Комарова и др. М.: Наука, 1988. 184 с.
- Мониторинг фитопопуляций/Л.Б.Заугольнова, О.В.Смирнова, А.С.Комаров, П.Г.Ханина//Успехи совр. биологии, 1993. Т.113. Вып.4. С.402-414.
- Вайнагий И.В. Методика статистической обработки материала по семенной продуктивности растений на примере Potentilla aurea L.//Растит. ресурсы, 1973. Т.9. Вып.2. С.287-296.
- Левина Р.Е. Репродуктивная биология семенных растений. Обзор проблемы. М.: Наука, 1981. 96 с.
- Ходачек Е.А. Семенная продуктивность растений Западного Таймыра//Структура и функции биогеоценозов таймырской тундры. Л., 1978. С.167-198.
- Раменский Л.Г. Проблемы и методика изучения растительного покрова. Избр. работы. Л.: Наука, 1971. 334 с.
- Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М., 1983. 197 с.
- Экологическая оценка кормовых угодий по растительному покрову/Л.Г.Раменский, И.А.Цаценкин, О.Н.Чижиков, Н.А.Антипин//М., 1956. 470 с. 31
- Горбачевская О.А., Жмылев П.Ю., Шинковская К.А. Очиток едкий: номенклатура, систематическое положение и изменчивость//Биологическая флора Московской области. М., 2000. Вып.14. С.101-111.
- Grime J.P., Hodgson J.G., Hunt R. Comparative plant ecology. London, 1988. 742 p.
- Рогожина Т.Ю. Комплексная оценка устойчивости декоративных травянистых многолетников//Ботанические сады -центры изучения и сохранения биоразнообразия. Якутск: Изд-во ЯГУ, 2003. С.93-97.
