Оценка состояния ценопопуляций редкого вида Oxytropis trichophysa bunge (Fabaceae) в Юго-Западной Туве
 в Юго-Западной Туве")
Автор: Селютина Инесса Юрьевна, Кониченко Елена Сергеевна, Зибзеев Евгений Григорьевич, Сенатор Степан Александрович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 5-1 т.20, 2018 года.
Бесплатный доступ
В статье приводятся результаты изучения биологических особенностей и демографической структуры популяций редкого вида Oxytropis trichophysa. Выявлена эколого-ценотическая приуроченность узколокального эндемика O. trichophysa в пределах юго-западной Тывы. На исследованной территории он встречается в качестве доминанта (или содоминанта) исключительно в высокогорных криофитных степях с Artemisia frigida и Festuca tschujensis на высотах от 2000 м над ур. м. Описан онтогенез вида, изучена онтогенетическая структура трех ценопопуляций в юго-западной Тыве. Онтогенез O. trichophysa простой, неполный, включает в себя 4 периода: латентный, прегенеративный, генеративный и постгенеративный и 8 возрастных состояний. Все изученные ценопопуляции вида являются неполночленными, дефинитивными, нормальными, зрелыми с бимодальным онтогенетическим спектром с максимумами на g1- и g3- особях. Высокая реальная семенная продуктивность и длительный генеративный период обеспечивают самоподдержание популяций O. trichophysa в экстремальных условиях высокогорных степей. Нуждается в охране из-за малого размера популяций и узколокальной эколого-ценотической приуроченности. Необходима охрана всех популяций данного вида в юго-западной Тыве.
Редкий вид, узколокальный эндемик, онтогенез, онтогенетическая структура
Короткий адрес: https://sciup.org/148314045
IDR: 148314045 | УДК: 581.522.582.736
Assessment of the status of a rare species Oxytropis trichophysa bunge (Fabaceae) coenopopulations in Southwestern Tuva
The article presents the results of studying the biological features and demographic structure of populations of the rare species Oxytropis trichophysa. The results our studies showed that O. trichophysa is a cryophyte steppe species. It has strict ecological and coenotic habitat conditions, which are confined to the specific cryophyte steppe with dominated Oxytropis trichophysa, Festuca tschujensis, Artemisia frigida at the altitudes of 2000 m above sea level. The ontogenesis and ontogenetic structure of the endemic rare species O. trichophysa were studied in southwestern Tuva. The onthogenesis of this species is simple, incomplete, includes 4 periods: latency, pregenerative, generative and postgenerative and 8 ontogenetic stages. As a result, ontogenetic structure of various cenopopulations was revealed. All studied coenopopulations O. trichophysa are not complete, definitive, normal, mature, have a bimodal age spectrum with peaks at g1- and g3-individuals. High real seed production and a long generative period provide self-maintenance of O. trichophysa populations in extreme conditions of high-altitude steppes. It needs protection because of the small size of the populations and the narrow-local ecologo-cenotic confinement. Protection of Oxytropis trichophysa in Southwestern Tuva is considered to be necessary.
Текст научной статьи Оценка состояния ценопопуляций редкого вида Oxytropis trichophysa bunge (Fabaceae) в Юго-Западной Туве
Oxytropis trichophysa Bunge (остролодочник волосисто-пузырчатый) (рис. 1) – узколокальный эндемик, ареал которого лежит в пределах южной Тувы, Юго-Восточного Алтая и западной части Монгольского Алтая (Пленник, 1976). В России O. trichophysa обитает на северном пре
деле распространения, где его ареал представлен двумя участками. Первый – в юго-восточном Алтае (хр. Чихачёва, Талдуаир, Сайлюгем), второй – в южной и юго-западной части Тувы на хребтах Сангилен (в верховье р. Сольбель-дер), Западный Танну-Ола (в верховье р. Кара-Чадан), Цаган-Шибэту (в верховье р. Барлык) и на хребте Монгун-Тайга (Определитель …, 2007; Красная книга…, 2008). За пределами России встречается в Западной Монголии (Ховд, Гоби-Алтай, Монгольский Алтай) и Китае (Тянь-Шань, Джунгарская Гоби) (Губанов,1996). Вид приурочен исключительно к криофитным степям высокогорного пояса.
-
O. trichophysa относится к секции Polyadena Bunge, представители которой отличаются железистым опушением, мутовчатым расположением листочков и распространены главным образом в степных районах Южной Сибири (Малышев, 2008) (рис. 2).
В Красную книгу РФ (2008) O. trichophysa внесен в статусе 3д – редкий вид, в Красную книгу Республики Тыва (Красная …, 2002) и Красную книгу Республики Алтай (Красная …, 2007) – в статусе 3 (R) – редкий вид (рис. 3). По данным сводок «Редкие и исчезающие растения Сибири» (Редкие …, 1980) и «Редкие и исчезающие

Рис. 1. Внешний вид O. trichophysa Bunge

Рис. 2. Железисто-опушенные листья и соцветия O. trichophysa
виды флоры Сибири: биология, охрана» (Семенова, 2007), O. trichophysa рекомендован для государственной охраны.
Некоторые локальные популяции охраняются на территории заповедника Убсунурская котловина. В качестве мер охраны рекомендованы организация заказника в долине р. Сыстыг-Хем (Республика Тыва) и мониторинг популяций (Красная …, 2008). По данным Красной книги РФ (Красная …, 2008), численность O. trichophysa не изучена, известные в Туве и Юго-Восточном Алтае популяции, по визуальной оценке полночленные, лимитирующим фактором является узкая экологическая амплитуда вида.
Общеизвестно, что для оценки состояния и сохранения редких видов необходимо изучение их биологических особенностей и структуры их популяций. Ю.А. Злобин (2009) в качестве одной из первоочередных задач популяционной экологии выделяет исследование популяций редких, исчезающих и охраняемых видов растений для разработки эффективных мер поддержания их стабильного существования.
Цель данного исследования – дать оценку состояния популяций редкого эндемичного вида O. trichophysa. . Для достижения этой цели нами были поставлены следующие задачи: выявление эколо-го-ценотической приуроченности O. trichophysa , изучение онтогенеза данного вида и онтогенетической структуры его ценопопуляций.
МАТЕРИАЛЫ И МЕТОДЫ
Изучение онтогенеза и онтогенетической структуры O. trichophysa проводили на территории Монгун-Тайгинского района Республики Тыва в июле 2014г (рис.3). Поскольку O. trichophysa является редким и эндемичным видом, изучение его биоморфологии и структуры ценопопуляции проводили с минимальным изъятием особей (Заугольнова, 1982). При выделении возрастных состояний придерживались общепринятой методики (Ценопопуляции …, 1976, 1988). Диагнозы возрастных состояний составляли на основе морфологических описаний 10-20 особей каждого возрастного состояния.
Изучение онтогенетической структуры це-нопопуляций проводили с применением методов, разработанных Т.А. Работновым (1950), А.А. Урановым (1975) и его школой (Ценопопу-ляции …, 1976, 1988). За счетную единицу принимали особь. Онтогенетическую структуру ЦП анализировали по критерию дельта-омега Л.А. Животовского (2001), а также с использованием таких демографических показателей, как ин-
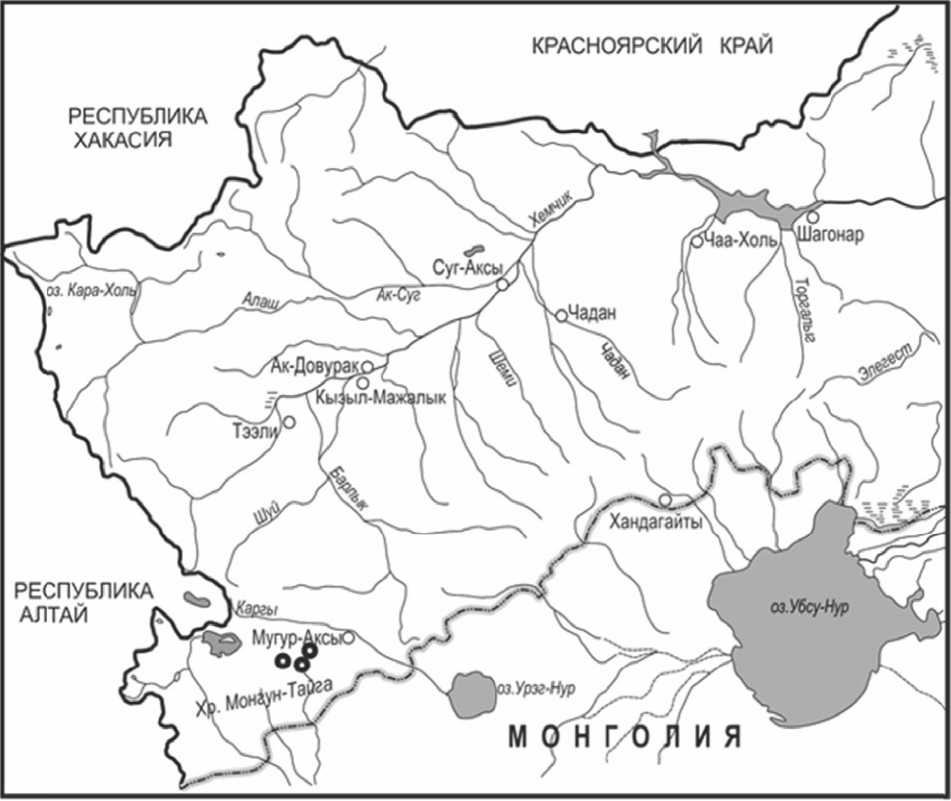
Рис. 3. Карта-схема изученных местообитаний O. trichophysa . Черными окружностями отмечены изученные ценотические популяции
дексы восстановления (Iв) и старения (Ic) (Глотов, 1998). Индекс эффективности ( ω ) рассчитывали по методике Л.А. Животовского (2001), индекс возрастности (∆) – А.А. Уранова (1975). Дополнительными критериями также служили плотность популяции и эффективная плотность популяции (число генеративных особей на единицу площади), доля прегенеративных и доля генеративных растений (Одум, 1986; Животовский, 2001). Полученные данные обработаны статистически (Зайцев, 1990) при помощи пакетa прикладных программ MS Excel 2007. Показатели семенной продуктивности определены по методикам И. В. Вайнагий (1974) и Левиной Р.Е. ( 1981).
Геоботанические описания выполнялись по стандартной методике ( Полевая…, 1964). Список сосудистых растений дан по С. К. Черепанову (1995).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Монгун-Тайга – горный массив юго-запада Тувы, расположен в междуречье Мугура, Шара-Харагая и Толайты. Он сложен кристаллическими сланцами и песчаниками, местами прорванными гранитами. Максимальная высота 3994 м над ур. м. Вершины хребта покрыты ледниковыми шапками, общая площадь которых составляет около 44 км². Для Монгун-Тайги характерен аридный горно-тундрово-горностепной тип поясности (Седельников, 1988).
Степной пояс занимает около трети горного массива, по северным склонам он поднимается до 2200 м, по южным до 3000 м. Высокогорные степи приурочены к малоразвитым щебнистым горно-степным почвам. Среди степных сообществ преобладают мелкозлаковые степи с доминированием Festuca tschujensis Reverd. , Festuca lenensis Drobow , Kobresia fi lifolia (Turcz.) C.B. Clarke, Kobresia myosuroides (Vill.) Fiori , Koeleria altaica (Domin) Krylov , Koeleria cristata (L.) Pers. и Poa attenuata Trin . , часто в качестве содоминан-тов выступают Artemisia frigida Willd. и Artemisia depauperata Krasch. В нижней и средней частях высокогорного пояса (2200-2700 м) широко распространены кобрезиевые ( Kobresia myosuroides) , овсяницевые ( Festuca sphagnicola B. Keller) и дри-адовые тундры ( Dryas oxyodonta Juz.).
Oxytropis trichophysa Bunge — вид с узким диапазоном экологической толерантности по отношению к увлажнению и занимаемому субстрату. На исследованной территории он встречается исключительно в высокогорных криофитных степях с содоминированием Artemisia frigida, Festuca tschujensis, Poa attenuata на высотах от 2000 м над ур. м. (рис. 4). Это редкие для Алтае-Саянской горной области сообщества, распространенные исключительно на территории Западной Тувы, Юго-Восточном Алтае и Западной части Монгольского Алтая. Видовой состав этих степей очень своеобразен, в нем в равной степени сочетаются, как типичные гор-

Рис. 4. Криофитная степь с доминированием O. trichophysa Bunge (Тыва, горный массив Монгун-Тайга)
но-степные виды, так и высокогорные виды ( Carex rupestris All. , Eritrichium subrupestre Popov , Festuca sphagnicola, Minuartia verna (L.) Heirn , Poa attenuata и др.). На всей территории ведется интенсивный выпас крупно-рогатого скота, лошадей и овец.
В результате интенсивного выпаса происходит изменение соотношения доминирующих и содоминирующих видов, при относительно стабильном сохранении видового состава. Уплотнение почвы, а также избирательное поедание животными трав приводит к увеличению проективного покрытия малопоедаемых трав, к которым относятся Artemisia frigida, Artemisia depauperata и Oxytropis trichophysa .
В то же время выпас скота оказывает негативное влияние на выживаемость проростков и особей прегенеративного периода Oxytropis trichophysa. В популяциях, подвергающихся более интенсивному выпасу в первую очередь элиминируются проростки, ювенильные и им-матурные особи.
I ЦП (Мугур) Чуйскотипчаковая степь (описание № 53.14ЗЕГ; 22.07.2014 г; 50°20 ’ 59" с. ш., 90°15 ’ 11" в. д.). Сообщество занимает склон северной экспозиции крутизной 1-2º, 2162 м над ур. м. Почвы горно-степные, щебнистые. Популяция представлена двумя локусами, разделенными грунтовой дорогой и отгонными тропами. Общее проективное покрытие (ОПП) 45%. Видовая насыщенность 24 вида на 100 м2. Вертикальная структура одноярусная, травостой представлен двумя подъярусами. Первый подъярус 10-15 см высотой образован Festuca tschujensis (12-15% от ОПП) и Oxytropis trichophysa (10%) , остальные виды при высоком постоянстве имеют проективное покрытие менее 1% ( Aster alpinus L. , Koeleria altaica, Leontopodium ochroleucum Beauverd , Poa attenuata, Pulsatilla ambigua (Turcz. ex Hayek) Juz.). Основная масса травостоя сосредоточена во втором подъярусе высотой 5-7 см, он представлен такими видами, как Allium pumilum Vved. , Artemisia frigida , Arenaria mongolica Schischk. , Carex duriuscula C.A. Mey ., Ephedra monosperma C.A. Mey. , Eritrichium subrupestre, Oxytropis eriocarpa Bunge , Oxytropis macrosema Bunge , Stellaria petraea Bunge in Ledeb. и др.
Второй локус занимает выровненный участок склона. При сохранении видового состава, изменяется соотношение групп содоминирующих видов. В частности, до 15% возрастает проективное покрытие Artemisia frigida , в травостое появляется Kobresia fi lifolia и Oxytropis tragachanthoides Fisch. Проективное покрытие Oxytropis trichophysa снижается до 7%.
III ЦП (Каргы 2). Остролодочниково-по-лынно-чуйскотипчаковая степь (описание № 55.14ЗЕГ; 22.07.2014 г; 50°23 ’ 03" с. ш., 90°17 ’ 36" в. д.). Сообщество занимает склон юго-юго-восточной экспозиции крутизной 3º, 2080м над ур. м. Почвы горно-степные, щебнистые россыпи занимают до 35%, участки оголенной почвы – до 15%. ОПП – 50%, видовая насыщенность 25 видов на 100 м2. Травостой не дифференцирован на подъяруса, его высота в среднем составляет 15 см. В качестве содоминантов выступают Festuca tschujensis (30%), Artemisia frigida (10%), Oxytropis trichophysa (8%). Проективное покрытие остальных видов не превышает 10%, это Agropyron cristatum (L.) see Palisot , Allium rubens Schrad. ex Willd. , Artemisia depauperata, Aster alpinus, Astragalus multicaulis Ledeb. , Carex duriuscula, Ephedra monosperma, Eremogone mongolica (Schischk.) Ikonn ., Euphorbia tshuiensis, Oxytropis pumila Fisch. ex DC ., Oxytropis tragacanthoides, Peucedanum vaginatum , Poa attenuata , Potentilla sericea L., Stellaria petraea Bunge in Ledeb .
IV ЦП (Мугур 2). Разнотравная степь (описание № 61.14ЗЕГ; 23.07.2014 г; 50°19 ’ 14" с. ш., 90°16 ’ 12" в. д.). Сообщество занимает выровненный верхний участок гряды на высоте 2263м над ур. м. Почвы горно-степные щебнистые слаборазвитые, каменистые россыпи занимают до 60%. ОПП – 40%, видовая насыщенность 24 видов на 100м2. Вертикальная структура одноярусная, высота травостоя 10-12см, его основу составляют криоксерофиты: Artemisia depauperata, Artemisia frigida, Carex rupestris, Eritrichium subrupestre, Festuca sphagnicola, Koeleria cristata, Minuartia verna, Oxytropis tragacanthoides, Poa attenuata, Potentilla sericea и др., являющиеся типичными представителями высокогорных степей Тувы и Юго-Восточного Алтая. Проективное покрытие Oxytropis trichophysa менее 1%. В отличие от выше описанных сообществ, данный вариант степей можно рассматривать как серийное сообщество, приуроченное к неплакорным местообитаниям.
Oxytropis trichophysa – стержнекорневой по-ликарпический травянистый гемикриптофит с многоглавым погруженным каудексом. По-беговая система представлена надземными и подземными побегами. Надземные побеги у взрослых особей двух типов: розеточные вегетативные (главный и боковые) и пазушные удлиненные генеративные. Вегетативные розеточные побеги полициклические с неопределенно долго функционирующей верхушечной почкой и моноподиальным нарастанием. Генеративные побеги монокарпические, безлистные, пазушные, сохраняются в течение некоторого времени после опадания цветков или плодов.
Подземная часть побеговой системы взрослых особей представляет собой ветвистый погруженный каудекс. На главах каудекса находятся спящие почки, а также остатки отмерших листьев. Корневая система представлена мощным, деревянистым, слабо ветвящимся главным корнем. В базальной части корня видны поперечные складки поверхностных тканей, свидетельствующие о его контрактильной деятельности, в результате которой каудекс постоянно оказывается погруженным в почву.
Стержнекорневые многоглавые травянистые многолетники с поликарпическими побегами розеточного типа являются биоморфами моноцентрического типа, не способными к вегетативному размножению. Размножение осуществляется только семенным путем (Лебедев, 1998).
Онтогенез O. trichophisa включает в себя 4 периода: латентный, прегенеративный, генеративный и постгенеративный и 8 онтогенетических состояний (рис. 5., табл. 1).
Латентный период. Бобы шаровидные, перепончатые, слабо бело- и черно-опушенные, мелкобугорчато- железистые, почти двухгнездные. Семена мелкие, округло-почковидные, зеленовато-коричневые или оливковые, длиной 2,20 ± 0,01 мм и 1,95± 0,01 мм шириной, созревают в конце июля – начале августа. Размножение осуществляется только семенным путем.
Прегенеративный период. В изученных природных популяциях проростки нами не были найдены, поэтому описание проростка даём по Р.Я. Пленник (1976).
Проросток состоит из крупных, мясистых, округло-яйцевидных семядолей, 7-8 мм длиной и 5-6 мм ширины. Первые листочки (проростка тройчатые, их 2, реже 1, причём второй – пятерной, с редкими волосками и железками. К концу вегетации оформляется лист с 16 мутовками листочков. Взрослый лист несет 12-14 мутовки листочков, расположенных односторонне по 2-3 и сближенных в 4-6-листные мутовочки. Первичный побег ортотропный, розеточный. Корень стержневой достигает 1,5- 3 см.
Ювенильные растения 1–3 года жизни представляют собой моноподиально-розеточно-стержнекорневые растения. Они имеют один розеточный побег, высота растения составляет 3,5 - 7,7 см. Розеточный побег с 2 - 3 листьями, на которых листочки располагаются 8–18 мутовками. Длина листьев от 2,8 до 5,5см.
Имматурные растения – растения 2–6 года жизни с 1-2 розеточным побегами. В начале этого состояния трогаются в рост пазушные почки, формирующие боковые розеточные побеги, так-

Рис. 5. Схема онтогенеза O. trichophysa
Se – cемя, p – проросток, j – ювенильная особь, im – имматурная, v – виргинильная, g1 – молодая генеративная, g2 – средневозрастная генеративная, g3 – стареющая генеративная, ss – субсенильная
Таблица 1. Морфометрические параметры особей разных онтогенетических состояний Oxytropis trichophysa
|
Онтогенетическое состояние |
Высота растения |
Диаметр кау-декса |
Число генеративных побегов |
Число вегетативных побегов |
Длина листа |
Длина листочка |
Ширина листочка |
Число мутовок в листе |
Число листьев на побег |
|
j |
5.6±0.9 |
0.5±0.0 |
- |
1.3±0.2 |
4.2±0.3 |
0.2 |
0.05±0.01 |
13.3±1.2 |
2.7±0.2 |
|
im |
6.4±0.3 |
1.4±0.2 |
- |
2.2±0.1 |
4.8±0.2 |
0.2±0.01 |
0.05±0.01 |
14.0±0.5 |
2.5±0.1 |
|
v |
7.9±0.5 |
4.3±0.6 |
- |
7.9±1.1 |
6.3±0.2 |
0.3±0.01 |
0.06±0.01 |
15.7±0.4 |
2.7±0.1 |
|
g 1 |
16.1±1.1 |
11.2±0.9 |
7.1±1.2 |
29.6±2.8 |
10.5±0.4 |
0.4±0.01 |
0.10±0.01 |
15.8±0.6 |
3.7±0.1 |
|
g 2 |
19.4±1.0 |
17.0±1.2 |
36.4±2.8 |
45.8±3.3 |
11.5±0.4 |
0.4±0.01 |
0.10±0.01 |
17.7±0.5 |
3.5±0.1 |
|
g 3 |
14.7±1.2 |
18.1±0.6 |
15.6±2.5 |
41.7±2.5 |
9.9±0.5 |
0.4±0.01 |
0.10±0.01 |
16.2±0.5 |
3.7±0.2 |
|
ss |
6.4±0.2 |
7.2±1.2 |
- |
15.6±1.2 |
5.1±0.2 |
0.2±0.01 |
0.10±0.01 |
12.2±0.7 |
2.8±0.2 |
же моноподиально нарастающие. Повреждение открытой почки возобновления моноподиально нарастающих побегов в прегенеративном периоде развития приводит к гибели побега и, часто, особи. Листьев на побеге 2–5, длиной 4-6см, листья с 9–23 мутовками. Формируется и становится заметным каудекс (диаметр его 0,8-3,5 см).
Виргинильные растения имеют хорошо развитый каудекс, его диаметр достигает 11см. Число моноподиально нарастающих розеточных побегов увеличивается до 7–16, на каждом побеге 3–4 листьев взрослого типа с 13–23 мутовками. Длина листа составляет 5–9,5 см.
Генеративный период. У молодых генеративных растений каудекс крепкий и плотный, диаметром до 17 см. Генеративные побеги образуются в пазухах листьев, их число достигает 2 - 8. На вегетативном розеточном побеге формируется 3–5 листьев взрослого типа с 18–22 мутовками. Многоглавый каудекс средневозрастных генеративных растений достигает в диаметре 27 см, на нем моноподиально нарастают боковые розеточные побеги, в розетке 3–5 листьев взрослого типа. Число генеративных побегов составляет 22–62. У старых генеративных растений каудекс многоглавый, диаметром до 23 см с обособленными, часто отмершими, главами. На каудексе появляются мертвые участки и полости. Вегетативных розеточных побегов – 29-54. На растениях формируется 4–26 генеративных побегов. Листья длиной 9-13 см, с 13-23 мутовками.
Постгенеративный период O. trichophysa проходит быстро. Характерными для субсенильных особей являются следующие признаки: побеги только вегетативные розеточные в числе 9-17 с листьями взрослого и переходного к сенильному типов, на побеге 2-4 листа, число мутовок на них сокращается до 9-14. Каудекс частично разрушенный, партикулирует без интеграции. Особи сенильного возрастного состояния не были обнаружены в изученных ценопопуляциях. Отсутствие особей этого онтогенетического состояния связано, скорее всего, с быстрой элимина- цией растений в постгенеративном периоде, о чём свидетельствует малая доля особей субсенильного состояния в ценопопуляциях.
Таким образом, онтогенез O. trichophysa простой, неполный (p-ss), остаётся неизменным в различных местообитаниях.
Все изученные ценопопуляции являются неполночленными, дефинитивными, нормальными, зрелыми с бимодальным онтогенетическим спектром с абсолютным максимумом на g3-особях и локальным – на g1-растениях. ЦП 4 (Мугур 2) - инвазионная, представлена двумя g2-особями и одной g1 и может считаться парцеллярной популяцией в понимании Т.А. Ра-ботнова (Работнов, 1969) или микропопуляцией (Жиляев, 2013).
Исследованные ценопопуляции отличалась низкими средними показателями плотности 0,3-0,8 особей/м2 и неполночленностью онтогенетического спектра (во всех популяциях отсутствуют сенильные особи, в ЦП 1 – j и im-особи). Максимальные значения экологической плотности (0,8 ос/м2 при варьировании числа особей от 0 до 7 на 1 кв. м) отмечены в ЦП1 (в чуйско-типчаковой степи) (табл. 2). В условиях полын-но-остролодочниково-чуйскотипчаковой степи (ЦП 2) и остролодочниково-полынно-чуйско-типчаковой степи (ЦП 3) плотность особей была меньше в 2 раза – 0,4 ос/м2 и 0,3 ос/м2 соответственно (при варьировании особей от 0 до 8 и 0 - 5 на 1 кв.м.). При этом показатели эффективной плотности популяции варьируют соответственно от 0,6 особи на 1м2 (ЦП 1) до 0,2 особи/ м2 (ЦП 2 и ЦП 3) , что свидетельствует о наличии во всех изученных ценотических популяциях стабильной генеративной фракции.
Среди особей прегенеративного периода наблюдается преобладание виргинильных (2,915%) и имматурных особей (0-12%) над ювенильными (0-3,8%). Отсутствие проростков и малая численность ювенильных растений в популяциях свидетельствует о слабом семенном возобновлении.
Таблица 2. Некоторые демографические показатели ценопопуляций Oxytropis trichophysa
|
№ ЦП |
Плотность |
Max: абсолютный , локальный |
Онтогенетическая структура (%), группа |
Д |
to |
1в |
1ст |
Тип ЦП |
|||
|
Экологическая, особей на м 2 |
эффективная, |
||||||||||
|
j, im, v |
g 1 - g 3 |
ss, s |
|||||||||
|
I |
0,8 |
0,64 |
g 1 , g 3 |
2,94 |
92,15 |
4,90 |
0,52 |
0,80 |
0,03 |
0,04 |
зрелая |
|
II |
0,4 |
0,24 |
g 1, g 3 |
31,25 |
62,5 |
6,25 |
0,41 |
0,61 |
0,33 |
0,06 |
переходная |
|
III |
0,3 |
0,20 |
g 1, g 3 |
22,22 |
73,33 |
4,44 |
0,48 |
0,67 |
0,23 |
0,04 |
переходная |
|
ценопопуляционные локусы в ЦП 1 |
|||||||||||
|
I.1 |
1,1 |
0,92 |
g 1, g 3 |
0 |
100 |
0 |
0,52 |
0,84 |
0 |
0 |
переходная |
|
I.2 |
0,5 |
0,36 |
g 1 , g 3 |
7,89 |
78,94 |
13,15 |
0,53 |
0,73 |
0,09 |
0,13 |
зрелая |
1.1 – склон северной экспозиции 1-2º крутизной, проективное покрытие (ПП) Oxytropis trichophysa – 10%, ПП Artemisia frigida до 2%;
1.2 – выровненный участок, ПП Oxytropis trichophysa – 7%, ПП Artemisia frigida – 15%
Онтогенетический спектр характеризуется значительным преобладанием генеративной группы особей (от 62,8% до 100% от общего числа особей). Во всех изученных популяциях преобладают особи старого генеративного состояния (g3) доля которых составляет от 35% в ЦП 2 до 46,7% в ЦП 3. Высокая жизнеспособность особей генеративного состояния позволяет им успешно существовать в экстремальных условиях высокогорных криофитных степей.
Оценка возрастности ∆ (дельта) и эффективности ω (омега) показала, что ЦП 1 (Мугур) относится к зрелым, а ЦП 2 (Каргы 1) и ЦП 3 (Каргы 2) - переходные (табл. 2). Важными популяционными параметрами, характеризующими интенсивность самоподдержания популяций и степень их старения, являются индекс возобновления и индекс старения. В целом, во всех изученных ЦП индекс старения низкий (от 0,04 до 0,06). В зрелой ЦП 1 наряду с низким индексом восстановления (Iв=0,03) отмечается и низкий индекс старения (Ic=0,04). В ЦП переходного типа индекс восстановления достаточно высок (Iв 0,23 – 0,33).
В ценопопуляции 1 (Мугур) выделяются два ценопопуляционных локуса (табл. 2). В локусе 1.1, который находится на склоне, экологическая плотность O. trichophysa составляет 1,1 ос/м2, в то время как в локусе 1.2 (на выполо-женном участке) плотность особей в два раза меньше – 0,5 ос/м2 . При этом онтогенетический спектр в первом локусе был представлен только генеративными особями (100%), в онтогенетическом спектре локуса 1.2 также есть виргинильные и субсенильные растения, доля генеративных особей – 79%.
В целом, в изученных популяциях особенно мала численность ювенильных растений, что свидетельствует о слабом семенном возобновлении. Многолетние наблюдения показали, что возобновление в популяциях в отдельные неблагоприятные годы не наблюдается или оно слабое, в годы с более благоприятными условиями увлажнения – активное (Лебедев, 1989; Санданов и др., 2014; Селютина, Санданов, 2015). Неблагоприятные условия (засушливые периоды, антропогенное влияние: выпас скота, рекреационная нагрузка) приводят к снижению активности возобновления и к элиминации особей в прегенеративном периоде онтогенеза. На затрудненное семенное возобновление указывает также низкая плотность ценопопуляций.
O. trichophysa – перекрестноопыляемый вид, цветет в июне - начале июля, плодоносит в июле – августе. Семенная продуктивность была изучена только в ЦП 1, поскольку у растений других ЦП семена еще не успели сформироваться. Потенциальная семенная продуктивность (ПСП) составила в среднем 13619 семязачатков на особь, реальная семенная продуктивность (РСП) – 6207 семян на особь. Процент семенификации (ПС) – 45%. Средние значения этого параметра свидетельствуют о неблагоприятных погодных условиях в период формирования семян, также средние значения ПС могут быть обусловлены низкой фертильностью растений. Одним из важных показателей семенной продуктивности служит число завязавшихся плодов, его можно рассматривать как первое производное от ПСП (Levina, 1981). Процент завязавшихся плодов является важным показателем, как критерий достаточности опыления. Для растений из разных популяций число цветков в соцветии варьирует незначительно - от 10,9±0,6 до 14,5±0,8, при этом 7-92% из них формируют бобы, то есть достаточность опыления у растений O. trichophysa очень сильно колеблется. По нашему мнению, это связано большей частью с экстремальными погодными условиями в период цветения, которые нередки в аридных высокогорьях (дождь, снег, сильный ветер, резкое понижение температуры), когда резко снижается посещаемость растений опылителями.
Тем не менее, формирование значительного числа генеративных побегов на растениях (в среднем 36,4±2,8) обеспечивает высокую потенциальную и достаточно высокую реальную семенную продуктивность особей и в совокупности с большой продолжительностью генеративного периода дает возможность устойчивого существования O. trichophysa в условиях высокогорных степей.
Таким образом, в результате проведенных исследований, выявлена эколого-ценотиче-ская приуроченность узколокального эндемика Oxytropis trichophysa в пределах юго-западной Тывы. Он встречается в качестве доминанта (или содоминанта) в своеобразных злаковополынных высокогорных криофитных степях ( Artemisia frigida Willd , Festuca tschujensis Reverd. и Poa attenuata Trin.) на высотах от 2000 м над ур. м. и выше. Онтогенез O. trichophysa простой, неполный, включает в себя 4 периода: латентный, прегенеративный, генеративный и постгенеративный и 8 возрастных состояний. Все изученные ценопопуляции этого вида являются неполночленными, дефинитивными, нормальными, зрелыми с бимодальным онтогенетическим спектром с максимумами на g1- и g3- особях. Высокая реальная семенная продуктивность и длительный генеративный период обеспечивают самоподдержание популяций O. trichophysa в экстремальных условиях высокогорных степей.
Список литературы Оценка состояния ценопопуляций редкого вида Oxytropis trichophysa bunge (Fabaceae) в Юго-Западной Туве
- Вайнагий И.В. О методике изучения семенной продуктивности растений // Ботанический журнал. 1974. Т. 59. №6. С. 826-831.
- Глотов Н.В. Об оценке параметров возрастной структуры популяций растений // Жизнь популяций в гетерогенной среде. Йошкар-Ола: Периодика Марий Эл, 1998. С. 146-149
- Губанов И.А. Конспект флоры Внешней Монголии (сосудистые растения). М.: Валанг, 1996. 136 с. [
- Животовский Л.А. Онтогенетические состояния, эффективная плотность и классификация популяций растений // Экология. 2001. №1. С. 3-7.
- Жиляев Г.Г. Роль микропопуляций в авторегуляции природных популяций Homogyne alpina (L.) Cass. (Asteraceae) на субальпийском лугу Черногоры (Украинские Карпаты) // Науковi основи збереження бiотичноï рiзноманiтностi. 2013. Т.4. №1. С. 29-42.
