Оценка состояния гнездовой орнитофауны пригородных лесов

Автор: Головатюк С.А.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Наземные экосистемы
Статья в выпуске: 2 т.30, 2021 года.
Бесплатный доступ
В статье анализируются особенности формирования видового состава и изменения плотности населения гнездящихся птиц малоизмененных насаждений и пригородных лесов.
Трофическая деятельность, наземногнездящиеся птицы, видовой состав, плотность населения
Короткий адрес: https://sciup.org/148322203
IDR: 148322203 | УДК: 598.2(470.57) | DOI: 10.24412/2073-1035-2021-10397
Assessment of the state of nesting avifauna in suburban forests
In this article analyzed features of species composition and changes in population density of nesting birds, in poorly modified stands and suburban forests.
Текст научной статьи Оценка состояния гнездовой орнитофауны пригородных лесов
Для оценки значения пригородных лесов в сохранении населения птиц, гнездящихся в нижних ярусах леса, использованы данные, полученные при проведении маршрутных учетов в гнездовой период 2016–2020 гг., на территории национального парка «Самарская Лука по стандартной методике [1–9].
В основу условного деления насаждений на «пригородные» и «малоизмененные», положено не только их территориальное расположение и близость к населенным пунктам, но и наличие (отсутствие) на их территории проявлений трофической деятельности копытных и последствий рекреационной деятельности.
Структура и возраст, видовой состав древостоя, подлесочно-кустарникового яруса и травяного покрова насаждений, на территории которых были заложены маршруты, преимущественно сходен. Маршрутный учет проводился на участках широколиственных лесов, в древостое которых доминирует липа мелколистная. Субдоминантом сообщества, в большинстве случаев выступает клен остролистный. В составе древостоя присутствуют также дуб черешчатый, вяз шершавый. Лесотаксационные характеристики насаждений, произрастающих в группе пригородных лесов и группе малоизмененных сообществ, в целом сопоставимы. Возраст преобладающей породы составляет 75–85 лет. В обследованных насаждениях максимальный возраст – до 125 лет, отмечается у деревьев дуба, не выбранного предыдущими рубками и оставленных
ранее на делянках как семенные деревья. Высота насаждений обеих групп составляет, в среднем 21–25 м. Средний диаметр деревьев первого яруса 24–28 см у липы и клена, до 60см у дуба. Сомкнутость крон растений первого яруса не менее 0,7–0,8. В среднем плотность произрастания древесных растений в насаждениях обеих групп составляет до 670 экз./га. Второй ярус в насаждениях выражен не всегда, при его наличии в нем преобладает тонкомерный клен и, в меньшей степени, вяз. Отличия в структуре подлесочнокустарникового яруса и травяном покрове – структурных элементах, определяющих качество защитно-гнездовых условий для наземногнездя-щихся птиц, не выражены. В формировании подлесочного яруса принимают участие лещина обыкновенная и бересклет бородавчатый. Плотность произрастания лещины до 1600 экз./га, при средней высоте растений 3–4,5 м, с толщиной стволов до 5,0 см. Широко представлены в подлеске и другие зоохорные виды – яблоня лесная, рябина, черемуха, боярышники. Травяной покров сложен комплексом преимущественно неморальных видов травянистой растительности.
Интенсивность рекреационного использования территории 57, 61 и 62 кварталов Жигулевского лесничества, составляющих группу «пригородных лесов» в настоящее время незначительна, рекреационная дигрессия лесных сообществ не превышает II–III стадии в 100-метровой полосе от стены леса, снижаясь до минимальных значений в глубине насаждения. В то же время пространственная изоляция полностью устраняет воздействие на современное состояние этих насаждений и их развитие проявлений трофиче- ской активности диких копытных. Насаждения второй группы, произрастающие на территории 38 квартала Жигулевского и 39, 43 и 44 кварталов Больше-Рязанского участковых лесничеств полностью свободны от рекреационного воздействия, но испытывают на себе проявление трофической активности диких копытных. Порои кабана в данных насаждениях занимают иногда (38-й квартал) до 30% площади. Как показали исследования, использование кабаном участков насаждений для добычи кормовых ресурсов, прямо связано с полным отсутствием назем-ногнездящихся птиц на этих участках. Последствия трофической деятельности лося более продолжительны и в полной мере проявляются через значительный интервал времени.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализировались данные, полученные при проведении маршрутных учетов в период 20162020 гг. За указанный период проведено 20 маршрутных учетов, 15 из которых были заложены на территории «пригородных лесов», 5 в «малоизмененных насаждениях». При проведении учетов на территории «пригородных лесов» отмечено 33 вида гнездящихся птиц, в «малоиз-мененных насаждениях» отмечено присутствие 21 вида. Общее количество видов учтенных при проведении маршрутных учетов 34.
В течение всего периода наблюдений плотность населения птиц «пригородных лесов» была выше аналогичных показателей полученных при проведении маршрутных учетов на территории естественных лесных экосистем и составляла в среднем 1014,9 пар/км2 и 748,8 пар/км2 соответственно. В наблюдаемый период превышение колеблется по годам от 21,0% в 2016 году до 50,0% в 2017 году и, в среднем за 5-ти летний период составляет 35,5%. Анализ экологического состава орнитофауны насаждений показывает сохранение ранее выявленных закономерностей.
Наиболее стабильным оказалось участие в составе населения гнездящихся птиц как «пригородных лесов», так и малоизмененных насаждений представителей экологической группы птиц, гнездящихся в убежищах. Долевое участие представителей указанной экологической группы в орнитофауне насаждений обоих типов отличается незначительно и составляет 49,1% в случае «пригородных лесов» и 50,4% в малоизмененных насаждениях. Несколько более разнятся показатели долевого участия в составе населения птиц изучаемых насаждений представителей экологической группы птиц гнездящихся открыто в средних и верхних ярусах насаждения, для «пригородных лесов» этот показатель составляет 41,1% для естественных сообществ 45,7%. Наибольшие отличия в долевом участии в составе населения птиц данных насаждений характерны для группы птиц, гнездящихся в нижних ярусах леса. В первом случае оно оставляет в среднем 9,6%, во втором случае 4,0% (рис. 1, 2).
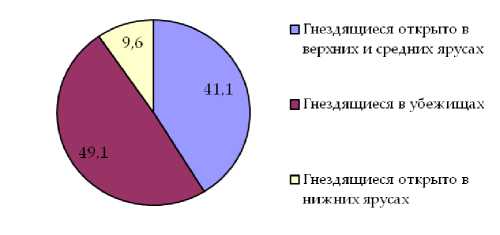
Рис. 1. Долевое участие в составе орнитофауны «пригородных лесов» экологических групп гнездящихся птиц по плотности населения
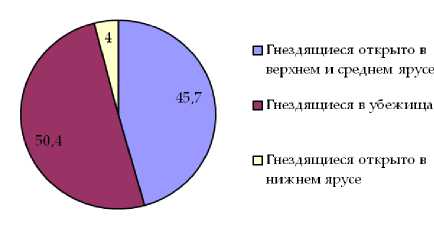
Рис. 2. Долевое участие в составе орнитофауны малоизмененных насаждений экологических групп гнездящихся птиц по плотности населения
Анализ видового состава свидетельствует о преобладании в составе орнитофауны как «пригородных лесов», так и малоизмененных насаждений видов группы птиц, гнездящихся в убежищах. Доля по числу видов птиц этой группы в составе населения птиц «пригородных лесов» составляет 45,0%, в естественных насаждениях 49,1%. Для группы птиц, гнездящихся в верхних и средних ярусах леса, показатель долевого участия по числу видов составляет 34,8% и 44,5% соответственно. Значительные отличия долевого участия в составе населения изучаемых насаждений по числу видов характерны для группы птиц, гнездящихся в нижних ярусах леса. В насаждениях первого типа оно составляет 20,3%, в насаждениях второго типа снижается до 6,5%.
Виды птиц, предпочитающие осветленные, разреженные леса по долевому участию в плотности населения уступают видам птиц, предпочитающим сомкнутые, затененные древостои в насаждениях обоих типов. В «пригородных лесах» данный показатель составляет 28,4% и 71,3% соответственно, в естественных сообществах 20,2% и 79,8%. Долевое участие по числу видов свидетельствует о большей привлекательности для птиц первой группы «пригородных лесов» по сравнению с естественными лесными сообществами, участие птиц данной группы по числу видов уменьшается от 45,1% в «пригород- ных лесах» до 35,3% в малонарушенных насаждениях. Для птиц предпочитающих сомкнутые, затененные древостои изменение долевого участия в составе населения по числу видов имеет обратную динамику, в «пригородных лесах» их долевое участие составляет 54,0% и возрастает до 64,7% в естественных лесных сообществах (рис. 3-6).
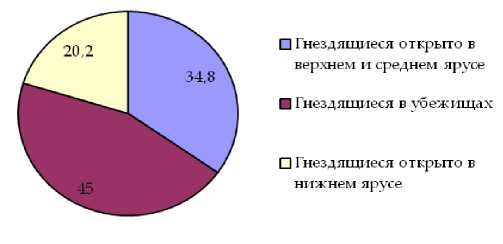
Рис. 3. Долевое участие в составе орнитофауны «пригородных лесов» экологических групп птиц по числу видов
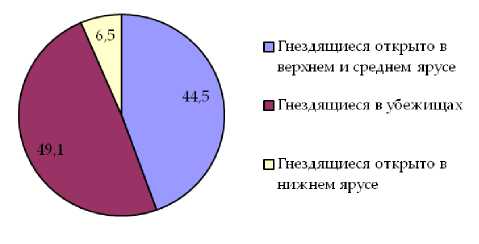
Рис. 4. Долевое участие в составе орнитофауны малоизмененных насаждений экологических групп птиц по числу видов
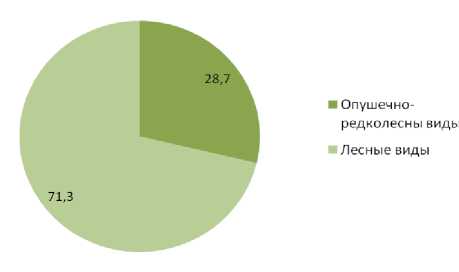
Рис. 5. Соотношение гнездящихся птиц с различной биотопической привязанностью в пригородных лесах
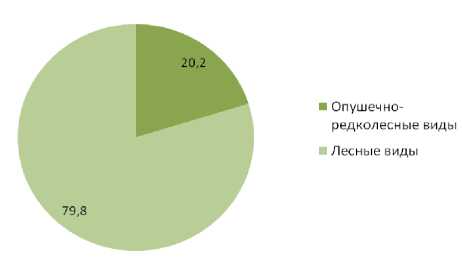
Рис. 6. Соотношение гнездящихся птиц с различной биотопической привязанностью в малоизмененных насаждениях
Коэффициент сходства Серенсена-Чеканов-ского видового состава орнитофауны «пригородных лесов» и «малоизмененных насаждений» составил 0,70, со значительным разбросом по экологическим группам. Максимальное сходство видового состава насаждений указанных типов наблюдается у группы птиц, предпочитающих гнездится в убежищах, коэффициент сходства для данной группы птиц составил 0,92. Минимальное сходство видового состава отмечается для группы птиц, гнездящихся в нижних ярусах леса, отмеченных при проведении маршрутных учетов на территории «пригородных лесов» и «малоизмененных насаждений», коэффициент сходства составил 0,18, что в два раза меньше показателей предшествовавшего пятилетнего периода. Для группы птиц, гнездящихся открыто в средних и верхних ярусах леса изучаемых насаждений, коэффициент сходства составил 0,71.
ВЫВОДЫ
Анализ данных, полученных при проведении маршрутных учетов на территории национального парка «Самарская Лука», в период 2016-2020 гг., свидетельствует о существующих значительных отличиях видового состава и плотности населения гнездящихся птиц малоизмененных насаждений и пригородных лесов. Большее видовое разнообразие орнитофауны пригородных лесов объясняется включением в состав населения птиц помимо типичных лесных видов, также опушечно-редколесных видов. Полученные данные, по предварительной оценке, позволяют утверждать, что пригородные леса имеют боль- шое значение в сохранении наземногнездящихся птиц.
Список литературы Оценка состояния гнездовой орнитофауны пригородных лесов
- Полякова Г.А., Малышева Т.В., Флеров А.А. Антропогенные изменения широколиственных лесов Подмосковья. М.: Наука, 1983. 118 с.
- Зацепина Р.А. Семейство славковые // Птицы Волжско-Камского края. М.: Наука, 1978. С. 94-134.
- Варнаков А.П. Питание кабана и его роль в биоценозах Молого-Шекснинского междуречья // Роль крупных хищников и копытных в биоценозах заповедников. М., 1986. С. 86-94.
- Быков Е.В. Воздействие выпаса на гнездовую орнитофауну пригородных лесов г. Жигулевска // Социально-экологические проблемы Самарской Луки. Тезисы докладов второй научной конференции. Куйбышев, 1990. С. 167-168.
- Головатюк С.А. Изменения состава гнездовой орнитофауны под влиянием трофической деятельности кабана // Известия Самарского научного центра РАН. 2014. С. 156-160.
- Экономов А.В., Кульпин А.А. Оценка качества и емкости среды обитания абана (Sus scrofa L., 1758) // Вестник Удмуртского университета. 2011. Вып. 4. С. 95-96.
- Приедниекс Я.Я., Куресоо А.У., Курлавичус П.И. Рекомендации к орнитологическому мониторингу в Прибалтике. Рига: Зинатне, 1986. 66 с.
- Фальковский П.К. Исследование влияния пастьбы скота в дубравах Тростянецкого лесничества на рост и производительность леса // Тр. по лесному опытному делу Украины. 1929. Вып. 8. С. 155-177.
- Sørensen T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons / Kongelige Danske Videnskabernes Selskab. Biologiske skrifter. 1948. Bind V, Nr. 4. 34 + [7] p.
