Оценка состояния экосистем в зоне воздействия горнодобывающих предприятий Среднего Прикамья

Автор: Ефимик В.Е., Митракова Н.В., Ефимик Е.Г., Фарзалиева Г.Ш.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 1, 2026 года.
Бесплатный доступ
Настоящее исследование является продолжением работ, начатых нами в 2021 г., как комплексное изучение почв, растительности и подстилочно-почвенной мезофауны на территории эксплуатации Верхнекамского месторождения калийно-магниевых солей. Целью данной работы стала сравнительная оценка состояния биотических компонентов природных и антропогенно-измененных экосистем на изучаемой территории. Для этого на 10 учетных площадках проводились описания растительности, отбор почвенных проб, морфологический и физико-химический анализ почв, оценивалoсь таксономическое разнообразие и структура населения почвенно-подстилочных беспозвоночных, проведен корреляционный анализ количественных показателей почвенной мезофауны с химическими свойствами почв исследуемых участков в болотных, луговых и лесных экосистемах в пределах зоны добычи и складирования отходов калийно-магниевых солей. Среди изученных верховые болота, суходольный луг и все лесные биогеоценозы не испытывают нагрузки засолением, что нельзя сказать о пойменных лугах, которые подвержены воздействию высокоминерализованных вод. Аллювиальные солончаковые почвы долин малых рек характеризуются засолением средней и высокой степени, сумма токсичных солей в верхних горизонтах варьирует от 0.3 и 0.9%, снижаясь до 0.26 и 0.1% в нижних слоях; доминирующими ионами являются Cl-, Ca2+ и Mg2+. В условиях таких почв происходит частичная трансформация видового состава растительных сообществ. Местные виды частично исчезают, а их место занимают устойчивые к засолению прибрежно-водные и влаголюбивые растения, нередко в том числе факультативно-галофитные и сорно-луговые. Мезофауна пойменных лугов имеет низкое таксономическое разнообразие и бедное население беспозвоночных за счет отсутствия типичных почвенных обитателей (Enchytraeida, Lumbricidae, Elateridae, Chilopoda и др.), при этом специализированных, устойчивых к засолению (галофильных) видов беспозвоночных не обнаружено. Результаты корреляционного анализа количественных характеристик почвенной мезофауны с химическими свойствами почв показали, что беспозвоночные, особенно насекомые, снижают свою численность при увеличении кислотности и концентрации токсичных солей в почвах.
Почвенно-подстилочная мезофауна, болотные, луговые и лесные экосистемы, техногенное засоление, Среднее Прикамье
Короткий адрес: https://sciup.org/147253587
IDR: 147253587 | УДК: 504.05+574.47+592+631.4 | DOI: 10.17072/1994-9952-2026-1-55-72
Assessing ecosystem state in the impact zone of mining enterprises in the Middle Kama region
This research continues the comprehensive study of soils, vegetation, and soil-and-litter mesofauna in typical biotopes initiated in 2021 within the Verkhnekamskoye Potash-Magnesium Deposit. The objective of this study is to comparatively assess biotopes located near industrial facilities and waste disposal areas, as well as in regions experiencing severe soil salinity. To characterize the vegetation and fauna at the survey locations, we conducted geobotanical field descriptions, collected invertebrates through soil sampling, and employed conventional methods to investigate soil properties. The taxonomic diversity and population dynamics of soil-litter invertebrates were examined at ten survey locations. Simultaneously, vegetation descriptions were recorded, and both morphological and physicochemical soil analyses were conducted. Additionally, a correlation analysis was carried out to relate the quantitative parameters of soil mesofauna to the chemical properties of the study site soils, and to relate the quantitative parameters of litter-dwelling invertebrates to those of litter or sod. The floodplain meadows along small rivers, exposed to waters with high mineral content, differed from typical biotopes. The alluvial solonchak soils found in small river valleys exhibit moderate to high levels of salinity. The total concentration of toxic salts in the upper soil horizons ranges from 0.3% to 0.9%, decreasing to a range of 0.26% to 0.1% in the deeper layers. The dominant ions are Cl-, Ca2+, and Mg2+. On such soils, the floristic diversity of plant communities undergoes notable changes. This is manifested in a partial loss of native species and the invasion by salt-tolerant local coastal-aquatic and moisture-loving plants, including facultative halophytes and sometimes ruderal meadow species. The mesofauna present in floodplain meadows exhibits low taxonomic diversity and a sparse population of invertebrates, primarily due to the lack of typical soil-dwelling organisms (such as Enchytraeida, Lumbricidae, Elateridae, Chilopoda, etc.). No obligate salt-tolerant (halophilic) invertebrates were identified. Correlation analysis between the quantitative characteristics of soil mesofauna and soil chemical properties revealed that invertebrates, particularly insects, generally experience a decline in abundance as soil acidity and toxic salt concentrations increase.
Текст научной статьи Оценка состояния экосистем в зоне воздействия горнодобывающих предприятий Среднего Прикамья
Антропогенное и промышленное воздействие на природу изучается давно и активно как за рубежом, так и в нашей стране [Bengtsson, Rundgren, 1984; Воробейчик, Садыков, Фарафонтов, 1994; Блинова, Редькина, 2005; Демаков и др., 2013; Gurov, Gurova, Pet’ko, 2014; Zhang, Van Gestel, Li, 2022 и др.]. Исследования, проведенные в ряде промышленных регионов России, показали, что при загрязнении воздушной среды выбросами металлургических комбинатов, химических предприятий и тепловых электростанций происходит изменение основных структурных параметров почвенной фауны: численности, биомассы, таксономического и трофического составов [Воробейчик, Садыков, Фарафонтов, 1994; Блинова, Редькина, 2005; Автаева, 2008; Танасевич, Рыбалов, Камаев, 2009; Воробейчик и др., 2012; Демаков и др., 2013].
Растительные сообщества на территориях, подвергающихся воздействию промышленных предприятий, также претерпевают определенные изменения (в некоторых случаях – очень существенные). Это отмечается как в других регионах [Zvereva, Kozlov, 2012; Rahmonov et al., 2023; Соколова и др., 2025], так и в Пермском крае [Москвина, Молганова, Овеснов, 2023; Ефимик, 2025].
Для каждой конкретной территории набор поллютантов и виды загрязнений зависят главным образом от специализации работающих здесь промышленных предприятий. На территории Пермского края расположены крупнейшие в России предприятия по добыче калийно-магниевых солей, при функционировании которых одной из основных экологических проблем является техногенное засоление [Хайрулина, 2014; Baklanov et al., 2019; Еремченко, Пахоруков, Шестаков, 2020]. Наиболее подвержены воздействию высокоминерализованных хлоридно-натриевых вод поймы малых рек на территории Верхнекамского месторождения калийно-магниевых солей [Еремченко, Пахоруков, Шестаков, 2020; Khayrulina et al, 2021].
Техногенное засоление оказывает непосредственное влияние на почвы, растительные сообщества и связанные с ними комплексы почвенных беспозвоночных. Но влияние этого фактора, как и степень его проявления, на указанные группы живых организмов детально не изучено. Непонятно, насколько серьезными могут быть последствия длительного засоления, в каких условиях это влияние выраженно сильнее, снижается ли биоразнообразие и, соответственно, устойчивость экосистем. В связи с этим становится актуальным изучение влияния техногенного засоления на растительность и почвенную мезофауну в Прикамье.
В последнее время в литературе чаще встречаются работы по изучению влияния солености почвы на отдельные модельные группы беспозвоночных, такие как коллемболы, энхитреиды, клещи [Pereira et al., 2015; Bicho, Scott-Fordsmand, Amorim, 2024] дождевые черви [Fang et al., 2025], а также на отдельные виды растений [Inelova, 2024]. Публикации, посвященные комплексному изучению последствий засоления бореальных экосистем, только появляются [Ефимик, Фарзалиева, Есюнин, 2022; Efimik et al., 2024].
Данное исследование является продолжением работ, начатых нами в 2021 г. как комплексное изучение почв, растительности и мезофауны типичных биотопов на территории эксплуатации Верхнекамского месторождения калийно-магниевых солей, которое в Пермском крае проводилось впервые [Ефимик, Фарзалиева, Есюнин, 2022; Efimik et al., 2024]. Целью настоящей работы стала сравнительная оценка состояния биотических компонентов природных и антропогенно-измененных экосистем на изучаемой территории.
Материалы и методика
Краткую характеристику Верхнекамского месторождения калийных солей мы давали ранее [Efimik et al., 2024]. В июле – августе 2024 г. на территории месторождения нами было обследовано 10 биотопов: два типа болот (пл. 1, 2), три типа лугов (пл. 3–5) и пять типов лесов (пл. 6–10) (рис. 1). Материал собирался на 10 учетных площадках в пределах указанных биотопов. На учетных площадках проведены геоботанические описания, взяты пробы беспозвоночных верхнего почвенного и подстилочного ярусов, составлены описания почв с отбором проб для морфологического и физико-химического анализа.
На основе полевых геоботанических описаний была составлена характеристика растительности учетных площадок [Методы изучения ..., 2002; Ипатов, Мирин, 2008]. Видовая принадлежность растений определялась непосредственно на местности, а также в камеральных условиях по собранному гербарному материалу с использованием преимущественно региональных определителей.
Мезофауна подстилки и верхнего почвенного слоя (0–15 см) собиралась по стандартной методике почвенных проб [Гиляров, 1975]. На всех 10 учетных площадках было взято по 10 проб размером 25 x 25 см каждая. Выемка беспозвоночных из проб производилась на месте. Их идентификация осуществлялась в камеральных условиях, где они определялись до отряда и/или семейства, подсчитывались и взвешивались. Модельные группы беспозвоночных животных – пауки, сенокосцы и многоножки – определялись до вида.
Отбор проб почв проведен на каждом исследованном участке в двух горизонтах. В почвенных пробах определены следующие показатели: органическое вещество методом мокрого сжигания по И.В. Тюрину [Теория и практика …, 2006]; актуальная и обменная кислотность (рНвод, рНсол) потенциометрическим методом; содержание подвижных соединений калия методом пламенной фотометрии в солянокислой вытяжке; емкость катионного обмена (ЕКО) по методу Бобко – Аскинази – Алешина1; гидролитическая кислотность – методом Каппена (в вытяжке 1М CH3COONa), основанном на титровании 1 н щелочью в присутствии фенолфталеина [Теория и практика…, 2006]. Количество и качество растворимых солей определяли в водной вытяжке: Na+ и К+ – методом пламенной фотометрии; Cl– – титрованием с азотнокислым серебром; Ca2+, Mg2+ – трилонометрическим методом2; SO42- – турбидиметрическим методом3.
Сумму токсичных солей определяли расчетным методом по Н.И. Базилевич и Е.И. Панкова [1968].
Статистическая обработка данных проводилась алгоритмами программ MS Exell 2013. Корреляционный анализ (Пирсона) количественных показателей почвенной мезофауны с химическими свойствами почв и подстилочных беспозвоночных с параметрами подстилки или дернины реализован в программе Past 4.13 [Hammer, Harper, Ryan, 2001].
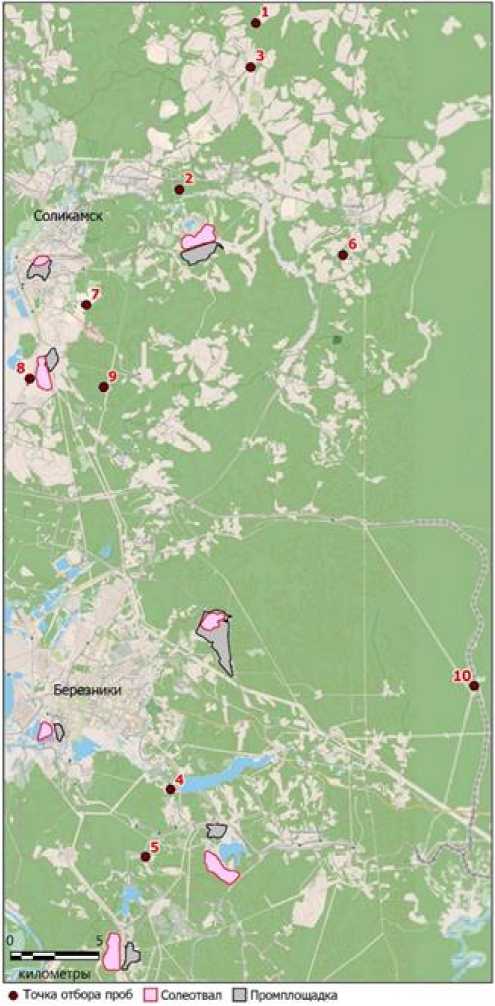
Рис. 1. Места отбора проб на территории Верхнекамского месторождения калийных солей. Обозначения: 1–10 – номера площадок как в тексте
[Sampling sites on the Verkhnekamskoye potash salt deposit territory.
Designations: 1-10 – site numbers as in the text]
Результаты и их обсуждение
Ниже представлен анализ результатов по группам биогеоценозов: болота (верховое пушицевосфагновое (пл. 1) и верховое осоково-сфагновое осушаемое (пл. 2)), луга (суходольный разнотравнозлаковый луг (пл. 3), пойменные луга (пл. 4–5) и леса. Среди последних были изучены: вырубка на месте елово-соснового леса (пл. 6), ельник черничный (пл. 7), елово-березовый заболоченный лес (пл. 8), березово-пихтово-еловый лес (пл. 9) и липово-елово-березовый лес (пл. 10).
В соответствии с ботанико-географическим районированием Пермского края [Овеснов, 2000], учетные площадки расположены в районе среднетаежных Камско-Печорско-Западноуральских пихтовоеловых лесов в подрайоне с преобладанием пихтово-еловых лесов (пл. 1–3, 6–9) и в районе южнотаежных Камско-Печорско-Западноуральских пихтово-еловых лесов с преобладанием осиновых и березовых лесов на месте темнохвойных (пл. 4, 5, 10).
Болота (пл. 1–2)
Болота на территории исследования широко распространены. Обследованы два болота верхового типа с характерным флористическим комплексом.
Верховое олиготрофное пушицево-сфагновое болото (пл. 1) имеет типичный флористический состав. Доминантами мохового яруса являются виды рода Sphagnum . Из трав наиболее обильны представители семейства Cyperaceae ( Eriophorum vaginatum L., Carex globularis L., C. disperma Dew.) и полукустарнички из семейства Ericaceae ( Vaccinium myrtillus L., V. uliginosum L., Oxycoccus palustris Pers., Andromeda polifolia L., Ledum palustre L.) и др. Болото окружено зарастающей вырубкой. Зарастание вырубки происходит сосной обыкновенной, березой повислой ( Betula pendula Roth) и б. пушистой ( B. pubescens Ehrh.), единично встречается ель сибирская ( Picea obovata Ledeb.). На границе собственно болота и вырубки расположено чернично-осоково-сфагновое сосновое редколесье с разреженным древостоем из Pinus sylvestris L. Разнообразие сосудистых растений болота низкое, что характерно для всех олиготрофных верховых болот данного ботанико-географического района.
Профиль почвы данного болота состоит из торфяного горизонта мощностью около 15 см и глеевой подстилающей песчаной породы. Верхняя часть торфяного горизонта состоит из сфагнового очеса, представляющего собой вертикально расположенные стебельки сфагновых мхов соломенно-желтого или буровато-желтого цвета, обычно сильно насыщенного влагой. Торфяно-глеезем характеризуется резко кислой реакцией среды (рН водн. = 3.8–4.2; рН сол. = 2.9–3.4). Содержание каждого из катионов и анионов не превышает 0.5 ммоль/100 г почвы. Содержание органического вещества в торфяном горизонте более 98.5%, в глеевой породе – 1.8%. Емкость катионного обмена выше средней (20–50 мг-экв/100 г почвы); содержание в почве обменного калия по профилю резко убывающее – от очень высокого (>100 мг/100г почвы) на глубине 0–15 см до очень низкого на глубине 15–30 см (1 мг/100 г почвы).
Верховое осоково-сфагновое осушаемое болото (пл. 2). Бо́льшая часть болотного массива осушена в связи с мелиоративными мероприятиями. Дренажная система на болоте привела к зарастанию ранее заболоченных биотопов густым подростом из березы пушистой, некоторые участки более активно зарастают сосной обыкновенной. В травяно-кустарничковом ярусе верховых болотных биотопов обильны вересковые кустарники и кустарнички: Ledum palustre , Vaccinium uliginosum , Oxycoccus palustris , Chamaedaphne calyculata (L.) Moench, также обильны Eriophorum vaginatum , осоки ( Carex disperma , C. limosa L.), встречаются Drosera rotundifolia L., отдельные особи Dryopteris carthusiana (Vill.) H.P. Fuchs., по краю болота – Chamaenerion angustifolium (L.) Scop., Vaccinium vitis-idaea L., Lycopodium annotinum L. Из мхов преобладают виды сфагнума и кукушкин лен обыкновенный ( Polytrichum commune Hedw.). Разнообразие сосудистых растений данного болота довольно низкое, что может быть связано с осушением и постепенным выпадением типичных болотных видов и пока слабым развитием типичных лесных растений.
На территории осушаемого болота диагностирован торфяно-подзол глеевый. Почва характеризуется сильнокислой реакцией среды (рН водн. = 4.7–4.8; рН сол. = 3.5); содержание органического вещества в подстилке 88.6%, в слое 5–20 см 5.42%, на глубине 20–30 см содержание органического вещества 5.25%; емкость катионного обмена средняя (22–24 мг-экв/100 г почвы), содержание в почве обменного калия очень низкое (2.5–2.8 мг/100 г почвы). Содержание водорастворимых ионов также не превышает 0.5 ммоль/100 г почвы, что свидетельствует об отсутствии засоления.
Видовое разнообразие беспозвоночных на верховом болоте (пл. 1) богаче, чем на осушаемом (пл. 2) – 29 против 21 вида соответственно, и складывается оно главным образом из многообразия пауков (12 видов из 5 семейств) и жуков (10 видов из 3 семейств). На осушаемом болоте (пл. 2) пауков было найдено значительно меньше (3 вида из 3 семейств), а разнообразие жуков оказалось таким же (10 видов из 3 семейств).
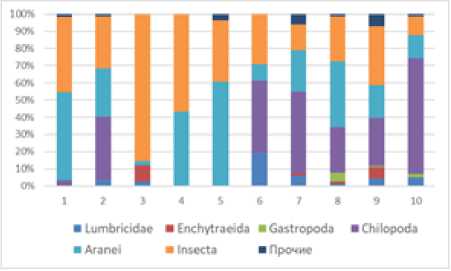
Пауки и жуки были не только разнообразными, но и многочисленными (104.0 экз./м2 или 50% и 68.8 экз./м2 или 33% от общей численности соответственно), составив основу населения верхового болота. На осушаемом верховом болоте ядро мезофауны оказалось несколько иным – его составили многоножки- костянки (52.8 экз./м2 или 35%), жуки (43.2 экз./м2 или 28%) и пауки (41.6 экз./м2 или 27%) (рис. 2А, 3А). Среди жуков наиболее многочисленными оказались на верховом болоте жуки Staphylinidae (40.0 экз./м2) и Carabidae (27.2 экз./м2), а на верховом осушаемом болоте – жуки Staphylinidae (25.0 экз./м2) (табл. 1).
Для обоих болот характерно полное отсутствие червей энхитреид (Enchytraeida) и незначительное присутствие дождевых червей (Lumbricidae – 6.4 экз./м2), но только на осушаемом болоте (табл. 1)
Illlllllll
123456789 10
-
■ lumbrictd*c ■ Enchytracida ■ Gastropoda ■ Chi topoda
-
■ Aranei ■ Insect* ■ Прение
В
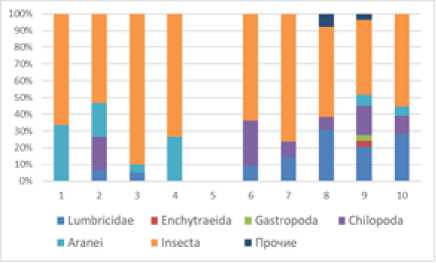
Рис. 3. Соотношение плотности (A) и биомассы (B) основных групп почвенной мезофауны на исследованных учетных площадках. Прочие группы: Diplopoda, Acariformes, Opiliones, Gastropoda [The ratio of density (A) and biomass (B) of the main groups of soil mesofauna at the studied accounting sites. Other groups: Diplopoda, Acariformes, Opiliones, Gastropoda]
Рис. 2. Соотношение плотности (А) и биомассы (B) основных групп мезофауны подстилки на исследованных учетных площадках. Прочие группы: Diplopoda, Acariformes, Opiliones
[The ratio of density (A) and biomass (B) of the main groups of mesofauna litter at the studied accounting sites. Other groups: Diplopoda, Acariformes, Opiliones]
Таблица 1
Таксономический состав, плотность (экз./м2 ± SE) и биомасса (г/м2) почвенно-подстилочной мезофауны на болотах (площадки 1 и 2) и на лугах (площадки 3–5)
[Taxonomic composition, density (specimens/m2 ± SE), and biomass (g/m2) of the soil-bedding mesofauna in swamps (sites 1 and 2) and meadows (sites 3-5)]]
|
Таксон |
Учетные площадки |
||||
|
1 |
2 |
3 |
4 |
5 |
|
|
Lumbricidae |
– |
4.840.5 1.6±0.2 |
6.4 ±0.8 1.6±0.2 |
– |
– |
|
Enchytraeida |
– |
– |
24.0 ± 2.4 0.0 |
– |
– |
|
Chilopoda |
6.4 ±0.7 0,0 |
48.0=4.2 4.8±0.5 |
– |
– |
– |
|
Acariformes: Trombidiidae |
3.2 ±0.3 0.0 |
1.6=0.2 0.0 |
– |
– |
1.6=0.2 0.0 |
|
Aranei |
99.2=8.4 4.8±0.5 |
36.8 ± 3.3 4.8±0.5 |
6.4=0;6 1.6±0.2 |
20.8 ± 1.9 6.4±0.6 |
27.2=2.6 0.0 |
|
Hemiptera: Heteroptera |
8.040.8 0.0 |
– |
17.6±1.6 0.0 |
0.0 11.2±1.3 |
1.6±0.2 0.0 |
Окончание табл. 1
|
Таксон |
Учетные площадки |
||||
|
1 |
2 |
3 |
4 |
5 |
|
|
Hemiptera: Homoptera |
6.4+06 0.0 |
1.6+0.2 0.0 |
1.6+0.2 0.0 |
– |
– |
|
Coleoptera sp. |
– |
– |
1.6 ± 0.2 3.2+0.4 |
1.6+0.2 0.0 |
1.6+0.2 0.0 |
|
Cantharidae, imago |
1.6+0.2 0.0 |
– |
– |
1.6+0.2 0.0 |
– |
|
Staphylinidae sp., imago |
19.2 ± 1.7 6.4±0.7 |
17.6+1.6 1.6+0.2 |
8.0+07 0.0 |
4.8 ±0.5 1.6+02 |
3.2+0.3 0.0 |
|
Staphylinidae sp., larva |
14.4+ 1.3 0.0 |
6.4+06 0.0 |
8.0+0.8 0.0 |
– |
– |
|
Carabidae, imago |
16.0=1.4 3.2+0.3 |
4.8+0.5 1.6+0.2 |
8.0+0.8 3.2+0.4 |
19.2 ± 1.9 4.8+0.5 |
– |
|
Carabidae, larva |
8.0+0.8 0.0 |
– |
– |
– |
1.6+0.2 0.0 |
|
Elateridae, larva |
– |
1.6+0.2 9.6+0.8 |
1.6+0.2 6.4+0.6 |
– |
– |
|
Lepidoptera, larva |
1.6+0.2 0.0 |
– |
1.6+0.2 0.0 |
– |
– |
|
Diptera, длинноусые, imago |
1.6=0.2 0.0 |
4.8+0.5 1.6+0.2 |
– |
– |
4.8+0.6 0.0 |
|
Diptera, pupa |
1.6=0.2 0.0 |
4.8+0.5 1.6+0.2 |
– |
– |
– |
|
Rhagionidae, larva |
1.6=0.2 0.0 |
– |
3.2+0.4 1.6+0.2 |
– |
– |
|
Bibionidae, larvae |
– |
– |
158.4+ 19.3 0.0 |
– |
– |
|
Прочие группы |
4.8+05 0.0 |
4.8+05 0.0 |
6.4 + 0.7 14.4+1.3 |
– |
3.2+03 0.0 |
|
Плотность по всем группам |
193.6+16.1 14.4+1.4 |
131.2+11.0 24.0+2.0 |
252.8+27.1 32.0+2.9 |
48.0+4.3 24.0+2.2 |
44.8+3.9 0.0 |
|
Суммарная плотность по двум слоям |
208.0±17.3 |
153.6+12.8 |
284.8±28.9 |
72.0±6.2 |
44.8±3.9 |
|
Биомасса по всем группам |
0.21/0.03 |
0.47/0.16 |
0.37/1.99 |
0.15/0.04 |
0.07/0.0 |
|
Суммарная биомасса по двум слоям |
0.24 |
0.63 |
2.36 |
0.19 |
0.07 |
Примечание: в числителе – плотность в подстилке, в знаменателе – плотность в почвенном слое; прочерк – отсутствие таксона. Прочие группы: Curculionidae, Scarabaeidae, Hymenoptera: Parasitica, Tipulidae, Psocoptera, Blattodea.
Луга (пл. 3-5)
Луга на территории обследования (за исключением пойменных) сформированы на залежах и на прочих участках, лишенных древесной растительности в результате деятельности человека.
Суходольный разнотравно-злаковый луг (пл. 3) характеризуется наличием широко распространенных луговых и сорно-рудеральных видов и достаточно богатым флористическим составом. Довольно обычны различные виды клеверов ( Trifolium Tourn. ex L.), Leucanthemum vulgare Lam., Hypericum perforatum L., Gnaphalium sylvaticum L., Rumex acetosella L., Anthoxanthum odoratum L., Achillea millefolium L., Hieracium caespitosum Dumort., Hieracium umbellatum L., Vicia cracca L, Agrostis tenuis Sibth. и A. gigantea Roth, Stellaria graminea L. и др.
На суходольном разнотравно-злаковом лугу (пл. 3) диагностирована серогумусовая (дерновая) почва. Почва характеризуется сильнокислой реакцией среды (рН водн = 4.9–5.5; рН сол = 3.6–4.1), содержание органического вещества в подстилке 63.8%, в серогумусовом горизонте – 1.12%, снижается с глубиной до 0.58%. Емкость катионного обмена низкая (14–16 мг-экв/100 г почвы), содержание в почве обменного калия очень низкое (1.5–1.8 мг/100 г почвы). Засоление отсутствует, сумма ионов не превышает 0.5 ммоль/100 г почвы.
Пойменные луга малых рек (пл. 4, 5) , подверженные воздействию высокоминерализованных сточных вод Верхнекамского месторождения калийно-магниевых солей, резко выделяются среди изученных биотопов. Именно в таких условиях техногенное засоление проявляется особенно сильно и непосредственно влияет на все живые организмы сообществ. Что касается растительных группировок, здесь наблюдается частичное выпадение аборигенных видов и зарастание устойчивыми к засолению местными прибрежно-водными и влаголюбивыми видами растений, в том числе факультативно-галофитными. На молодых техногенно-засоленных почвах наиболее распространенными видами, устойчивыми к засолению, являются: Phragmites australis (Cav.) Trin. ex Steud., Typha latifolia L., Phalaroides arundinacea (L.) Rauschert, Lactuca tatarica (L.) C.A. Mey., Juncus compressus Jacq., Triglochin palustris L., Puccinellia distans (Jacq.) Parl., Deschampsia cespitosa (L.) P. Beauv., Elytrigia repens (L.) Nevski и некоторые другие. Большинство из них – типичные прибрежно-водные и влаголюбивые солеустойчивые виды с широкими (или даже космополитными) ареалами, а также сорно-луговые. Наряду с вышеотмеченными видами встречаются факультативно-галофитные виды, в естественном ареале часто встречающиеся по морским побережьям или в условиях природного засоления: Spergularia marina (L.) Griseb. ( S. salina J. Presl & C. Presl), Atriplex patens (Litv.) Iljin, Triglochin maritima L., Puccinellia hauptiana V.I. Krecz. Наиболее засоленные участи часто полностью лишены какой-либо растительности.
Почва в пойме р. Зырянки (пл. 4) диагностирована как аллювиальная перегнойно-глеевая глинистая солончаковатая. С глубины 35 см сочится вода, отмечается заметный запах сероводорода, почва бесструктурная, в профиле отмечаются пятна ржавой и сизой окраски, что свидетельствует о наличии подвижного железа в окисной и закисной формах. Почва нейтральная рНвод = 6.9 и 7.3 в верхнем и нижнем слоях соответственно, гидролитическая кислотность низкая 1.74–1.18 ммоль/100 г.
Отмечено, что долговременное влияние высокоминерализованных вод на аллювиальные почвы приводит к изменению кислой реакции на нейтральную [Пахоруков, Еремченко, 2021; Khayrulina et al., 2021].
В слое 0–10 см содержание органического вещества 28.3%, что обусловлено перегнойным горизонтом, количество органического вещества резко снижается с глубиной до 2.68%. Емкость катионного обмена очень высокая в верхнем слое – 99 мг-экв/100 г почвы, что связано с высоким содержанием органического вещества и глинистым гранулометрическим составом, с глубиной ЕКО снижается почти в 2.5 раза. Количество обменного калия уменьшается с 15.3 мг/100 г до 6.8, что свидетельствует о среднем и низком уровне обеспеченности по градации [Вальков и др., 2004]. Количество хлорид ионов в верхнем слое почвы 5.51 ммоль/100 г, в слое 10–20 – 1.27; содержание ионов кальция, магния и натрия в слое 0– 10 см составило 2.63, 1.25 и 2.18 ммоль/100 г соответственно, с глубиной содержание ионов уменьшается в 3–5 раз. Сумма токсичных солей для слоя 0–10 см составила 0.31%, что характеризует верхний слой почвы как среднезасоленный; для слоя 10–20 см сумма токсичных солей составила 0.1%, что свидетельствует о слабом засолении.
Почва на заболоченном пойменном лугу (пл. 5) – аллювиальная перегнойно-глеевая глинистая солончаковая. Почва бесструктурная, плотная, пластичная, отмечены пятна ржавого и сизого цветов, что свидетельствует о наличии подвижного железа в формах Fe3+ и Fe2+.и протекающих процессах оглеения. Почва кислая, рН водное варьирует с 3.6 до 5.1 с поверхности до глубины 35 см, гидролитическая кислотность в верхнем слое 4–10 см составила 44.9 ммоль/100 г, с глубиной снижается до 6.69 и 3.48 ммоль/100 г. Содержание органического вещества в верхнем слое очень высокое – 35%, что связано с наличием слаборазложившегося органического вещества, с глубиной его количество резко снижается более чем в 10 раз. Засоление и обводненность снижают скорость минерализации. ЕКО свидетельствует о высокой емкости катионного обмена. Уровень содержания обменного калия низкий – 7.6–3.0 мг/100 г. По всему исследованному профилю (до 35 см) отмечается высокое количество ионов Cl-, Ca2+ и Mg2+. Так, в слое 4–10 см количество хлорид ионов составляет 17.9 ммоль/100 г, ионов кальция и магния 11.75 и 5.0 ммоль/100 г соответственно. Содержание гидрокарбонат ионов по профилю не превышает 0.2 ммоль/100 г, натрия – 0.4 ммоль/100 г, калия – 0.1 ммоль/100 г. В слое 15–25 количество ионов Cl-, Ca2+ и Mg2+ составило 5.04; 3.25 и 2.0 соответственно; а в слое 25–35 – 4.82, 3.13 и 1.88 ммоль/100 г. Сумма токсичных солей по горизонтам сверху вниз составляет 0.9, 0.26 и 0.25%. Количество токсичных солей в размере 0.9% свидетельствует об очень сильном засолении, 0.25–0.25% – о среднем засолении.
Почвенно-подстилочная мезофауна изученных лугов демонстрирует яркие различия между фаунами суходольного и пойменных засоленных лугов. На суходольном лугу выше видовое и таксономическое разнообразие (25 видов из 7 отрядов и 3 классов беспозвоночных). На пойменном засоленном лугу (пл. 4) обнаружено 18 видов из 3 отрядов и 2 классов, а на пойменном засоленном лугу (пл. 5) – 12 видов из 7 отрядов и 2 классов. Также на пойменных лугах не были найдены земляные черви (Enchytraeida и Lumbricidae) и личинки жуков-щелкунов (Elateridae), отмеченные на суходольном лугу; заметно низкое разнообразие жуков (особенно на пойменном лугу (пл. 5)).
Оригинальность фауны беспозвоночных на пойменном засоленном лугу (пл. 5) составила 16.7%, на засоленном лугу (пл. 4) – 22.2%. При этом никаких специфичных, тем более галофильных, видов беспозвоночных обнаружено не было.
Особенно показательна бедность населения беспозвоночных на обоих засоленных пойменных лугах (72 экз./м2 на первом и всего 44.8 экз./м2 на втором) и полное отсутствие почвенных беспозвоночных на засоленном лугу (пл. 5) (табл. 1). Примечательно, что численность беспозвоночных на разнотравном суходольном лугу в 4–6 раз больше, чем на засоленных лугах. Высокий показатель плотности населения на этом лугу (284.8 экз./м2) объясняется тем, что в одной из 10 проб попалась кладка личинок комаров-толстоножек (Bibionidae) с показателем в 158.4 экз./м2. Но, даже несмотря на это, численность беспозвоночных на суходольном лугу будет в 3 раза выше таковой обоих пойменных засоленных лугов.
Леса (пл. 6–10)
Все изученные лесные биогеоценозы – вторичны и сформированы на бывших вырубках и на зарастающих лугах.
Вырубка на месте сосново-елового леса (пл. 6) находится на кустарниковой стадии сукцессии. Из кустарников доминируют Sorbus aucuparia L. и Rubus idaeus L. Отмечен подрост мелколиственных пород. В травяном ярусе преобладает Chamaenerion angustifolium , также отмечены Stellaria holostea L., Rumex acetosella L., Calamagrostis epigeios (L.) Roth, C. arundinacea (L.) Roth, Solidago virgaurea L., Melica nutans L., Urtica dioica L. и др. Среди единичных особей хвойных пород, уцелевших при вырубке, встречаются угнетенные повышенным освещением Trientalis europaea L., Dryopteris carthusiana (Vill.) H.P. Fuchs, Maianthemum bifolium (L.) F.W. Schmidt, Luzula pilosa (L.) Willd.
Почва на вырубке относится к подзолу грубогумусированному песчаному. Подстилочно-торфяный горизонт мощностью около 3–4 см, в нижней части которого обнаружен грубогумусовый материал. Подзолистый горизонт маломощный, перетекает в альфегумусовый. Подзолистый горизонт характеризуется сильнокислой реакцией среды (рН водн = 4.8–5.2; рН сол = 3.7–4.1). Содержание органического вещества низкое, с глубиной снижается, что обусловлено природой и гранулометрическим составом подзола. Так, подзолистый горизонт содержит 1.02% органического вещества в слое 3–10 см; альфегумусовый – 0.66%. Емкость катионного обмена низкая (12–16 мг-экв/100 г) по критериям Валькова [Вальков и др., 2004], содержание в почве обменного калия очень низкое (1.1–1.2 мг/100 г). Количество водорастворимых катионов (Ca2+, Mg2+, Na+, K+) и анионов (HCO2-, Cl-, SO 4 2-) не превышает 0.5 ммоль/100 г., что свидетельствует об отсутствии засоления.
Древесный ярус ельника черничного (пл. 7) образован елью сибирской, береза повислая единична. Под пологом хорошо развит разновозрастный подрост ели, пихты немногочисленны. Из кустарников отмечены Sorbus aucuparia и Amelanchier spicata (Lam.) C. Koch. В травяно-кустарничковом ярусе доминирует Vaccinium myrtillus, обычны папоротники ( Dryopteris carthusiana , Gymnocarpium dryopteris ), с ними соседствуют Luzula pilosa , Oxalis acetosella L., Trientalis europaea , Rubus saxatilis L., Maianthemum bifolium , Melampyrum pratense L. и др. Зеленые мхи формируют выраженный ярус.
Серогумусовая легкосуглинистая почва ельника черничного состоит из подстилки, мощностью 3 см, серогумусового горизонта мощностью 9 см, который постепенно переходит в материнскую породу облегченного гранулометрического состава. Почва резко кислая (рН водн. = 4.3–4.9; рН сол. = 3.3–3.8). Содержание органического вещества в серогумусовом горизонте 3.71%, снижается с глубиной до 1.73%; в подстилочном горизонте составило 26.2%. Емкость катионного обмена средняя (17–26 мг-экв/100 г почвы), обеспеченность почвы подвижным калием по критериям [Вальков и др., 2004] средняя (9.5–12.7 мг/100 г почвы). Содержание каждого из катионов и анионов не превышает 0.5 ммоль/100 г почвы.
Смешанный заболоченный елово-березовый лес (пл. 8) в древесном ярусе образован елью сибирской, березой пушистой и б. повислой. В кустарниковом ярусе отмечена Frangula alnus Mill., Padus avium Mill. В травяно-кустарничковом ярусе представлены Geum rivale L., Calla palustris L., Equisetum fluviatile L., Lycopus europaeus L., Filipendula ulmaria (L.) Maxim., Deschampsia cespitosa (L.) Beauv., Viola epipsila Ledeb., Cirsium palustre (L.) Scop. и ряд других. Из мхов преобладают виды рода Sphagnum .
На исследованном участке заболоченного леса диагностирована торфяная почва. Почва сильнокислая (рН водн. = 4.8–6.1; рН сол. = 5.0–5.3); содержание органического вещества в слое 0–20 см 80.1%, в слое 20– 40 см – 85.9%, что в целом характерно для торфяных почв; емкость катионного обмена очень высокая – >50 мг-экв/100 г почвы, содержание в почве обменного калия среднее 32.0–56.3 мг/100 г почвы, Наличие каждого из катионов и анионов не превышает 0.5 ммоль/100 г почвы, что свидетельствует об отсутствии засоления.
Березово-пихтово-еловый крупнопапоротниковый лес (пл. 9) характеризуется хорошо выраженной ярусностью. В древесном ярусе преобладают Picea obovata , Abies sibirica Ledeb. и Betula pendula . В подлеске представлены немногочисленные кустарники: Padus avium , Rubus idaeus , Sorbus aucuparia , Lonicera xylosteum L. и др. В травяном ярусе доминируют крупнолистные виды папоротников: Dryopteris filix-mas (L.) Schott., D. carthusiana , Athyrium filix-femina (L.) Roth. Также отмечены Aegopodium podagrar-ia L., Lathyrus vernus (L.) Bernh., Stellaria holostea L., Pulmonaria obscura Dumort, Asarum europaeum L., Milium effusum L. и др. Моховой ярус развит слабо. Хорошо развита подстилка из хвойно-лиственного опада.
В березово-пихтово-еловом лесу диагностирована серогумусовая супесчаная почва. Профиль характеризуется наличием подстилки мощностью около 3 см, серогумусовым горизонтом, переходящим в песчаную породу. Почва резко кислая (рН водн. = 4.2–5.1; рН сол. = 3.3–3.8). Содержание органического вещества в серогумусовом горизонте варьирует от 3 до 5%, снижается с глубиной до 1.5–2%; в подстилочном горизонте около 30%. Емкость катионного обмена средняя (17–28 мг-экв/100 г почвы), обеспеченность почвы подвижным калием средняя (9.5–17 мг/100 г почвы). Содержание водорастворимых ионов менее 0.5 ммоль/100 г почвы.
Липово-елово-березовый крупнопапоротниковый лес (пл. 10) характеризуется древесным ярусом из хвойных и лиственных пород – Tilia cordata Mill., Picea obovata , Betula pendula . Подрост представлен активным возобновлением ели сибирской и липы сердцелистной, пихта в подросте немногочисленна. В подлеске малина, рябина и черемуха обыкновенная. В травяном ярусе преобладают крупные папоротники – Dryopteris filix-mas , D. expansa (C. Presl) Fraser-Jenk. & Jermy, Athyrium filix-femina , встречается Matteuccia struthiopteris (L.) Tod. Совместно с ними отмечаются преимущественно неморальные виды трав – Stellaria holostea , Pulmonaria obscura Dumort, Aegopodium podagraria , Equisetum sylvaticum , Asper-ula odorata L., Aconitum septentrionale Koelle, Asarum europaeum , Calamagrostis arundinacea (L.) Roth и др. Моховой ярус не выражен. Хорошо развита подстилка.
В липово-елово-березовом лесу диагностирована серогумусовая суглинистая почва. Профиль почвы включает подстилку мощностью 2 см, серогумусовый горизонт, залегающий на суглинистой породе. Мощность гумусового горизонта 12 см, ниже гумусового горизонта залегает суглинистая материнская порода. Почва сильнокислая (рН водн. = 5.0–5.1; рН сол. = 3.8). Содержание органического вещества в гумусовом горизонте 5.08%, с глубиной снижается до 1.53%. Емкость катионного обмена средняя (21–28 мг-экв/100 г почвы), содержание в почве обменного калия среднее (7.7–17 мг/100 г почвы). Содержание каждого из катинов и анионов в водной вытяжке почвы не превышает 0.5 ммоль/100 г почвы.
Мезофауна лесных биотопов демонстрирует широкий спектр качественных и количественных параметров. Наибольшим таксономическим разнообразием беспозвоночных выделяются елово-березовый заболоченный лес (43 вида из 10 отрядов и 6 классов животных) и березово-пихтово-еловый лес (31 вид из 11 отрядов и 6 классов). При этом более высокое разнообразие видов в этих лесах показали отряды Aranei (11 и 8 видов соответственно) и Coleoptera (9 и 14 видов соответственно). Низкое таксономическое разнообразие беспозвоночных было отмечено на вырубке елово-соснового леса – 14 видов из 7 отрядов и 4 классов (табл. 2).
Таблица 2 Таксономический состав, плотность (экз./м2 ± SE) и биомасса (г/м2)
почвенно-подстилочной мезофауны лесов
[Taxonomic composition, density (specimens/m2 ± SE) and biomass (g/m2) of the soil-litter mesofauna of forests]
|
Таксон |
Учетная площадка |
||||
|
6 |
7 |
8 |
9 |
10 |
|
|
Lumbricidae |
19.2 ± 1.7 1.6±0.2 |
8.0 ±0.8 43=0.5 |
1.6 ±0.2 6.4=03 |
8.0 ±0.7 9.64=0.9 |
8.0=03 8.0=03 |
|
Enchytraeida |
– |
1.6 ± 0.2 0.0 |
13±0.2 0.0 |
11.2=1.2 1.64=0.2 |
– |
|
Gastropoda |
– |
– |
6.4±03 0.0 |
1.6=0.2 1.64=0.2 |
3.2=0.4 0.0 |
|
Diplopoda |
– |
– |
0.0 13=0.2 |
1.64=0.2 1.64=0.2 |
– |
|
Chilopoda |
416=5 6 4.84=0.5 |
62.4=5.4 3.2±0.3 |
32.0=23 13=0.2 |
46.4=4.0 8.04=0.7 |
104.4=83 3.24=0.4 |
|
Trombidiformes |
– |
1.6=10.2 0.0 |
13=0.2 0.0 |
– |
– |
|
Opiliones |
– |
6.4=03 0.0 |
– |
8.0±03 0.0 |
1.64=0.2 0.0 |
|
Aranei |
9.6 ±0.8 0.0 |
32.0 ± 2.8 0.0 |
46.44=3.9 0.0 |
333=23 0.0 |
20.84=22 13±0.2 |
|
Hemiptera: Heteroptera |
3.2=03 0.0 |
1.6±0.2 0.0 |
– |
83±03 3.2±03 |
0.0 3.2=0.4 |
|
Hemiptera: Auchenorrhyncha |
13=0.2 0.0 |
– |
– |
1.64=0.2 0.0 |
– |
|
Coleoptera sp. |
0.0 43=0.6 |
0.0 3.2=03 |
43±0.5 0.0 |
1.64=0.2 0.0 |
– |
Окончание табл. 2
|
Таксон |
Учетная площадка |
||||
|
6 |
7 |
8 |
9 |
10 |
|
|
Chrysomelidae |
– |
0.0 3.240.4 |
1.640.2 3.240.3 |
1.6402 1.6402 |
– |
|
Staphylinidae |
3.240.3 0.0 |
3.040.8 3.240.3 |
4.840.5 1.6402 |
19.241.6 3.240.35 |
12.841.1 0.0 |
|
Carabidae |
– |
3.2 ± 0.3 0.0 |
14.441.3 1.6402 |
12.8=12 1.640.2 |
3.240.3 0.0 |
|
Elateridae. larva |
– |
3.240.3 14.441.3 |
1.640.2 3.240.3 |
14.440.9 6.440.6 |
1.6402 8.040.7 |
|
Lepidoptera. larva & pupa |
3.2 ±0.4 0.0 |
– |
– |
4.840.5 0.0 |
– |
|
Diptera . короткоусые. larva |
14.4=1.6 0.0 |
1.6402 0.0 |
1.640.2 0.0 |
1.6402 1.6402 |
– |
|
Diptera . Bibionidae. larva |
3.240.4 0.0 |
– |
– |
– |
– |
|
Tabanidae. pupa & larva |
– |
0.0 1.640.2 |
– |
– |
0.0 1.640.2 |
|
Rhagionidae. larva |
– |
- |
3.240.3 0.0 |
1.640.2 3.240.4 |
– |
|
Hymenoptera (Parasitica). imago |
– |
1.6402 0.0 |
– |
– |
0.0 1.6402 |
|
Прочие группы |
0.0 6.440.8 |
– |
0.0 - |
3.240.4 0.0 |
– |
|
Плотность по всем группам |
99.2=8.4 17.641.8 |
131.2410.9 33.642.9 |
121.2410.3 20.841.8 |
171.2414.3 46.443.9 |
155.2413.3 28.842.4 |
|
Суммарная плотность по двум слоям |
116.8±9.8 |
164.8±13.7 |
142.4±12.1 |
217.6±18.0 |
184.0±15.5 |
|
Биомасса по всем группам |
0.68/0.14 |
0.61/1.44 |
0.26/0.97 |
0.91/0.71 |
0.80/0.57 |
|
Суммарная биомасса по двум слоям |
0.82 |
2.05 |
1.23 |
1.62 |
1.37 |
Примечание: в числителе – плотность в подстилке, в знаменателе – плотность в почвенном слое; прочерк – отсутствие таксона. Прочие группы: Cantharidae, Curculionidae, Hymenoptera: Symphyta.
березовом лесу до 34.2% – в березово-пихтово-еловом лесу (рис. 2 А–Б). В почвенном слое доля насекомых оказалась значительной и составила от 63.6% на вырубке до 76.2% в ельнике (рис. 2 А–Б). Наиболее многочисленными насекомыми стали жуки коротконадкрылые (Staphilinidae), жужелицы (Carabidae) и жуки-щелкуны (Elateridae) (табл. 2).
Обсуждение
Нами были изучены почвы, растительность и почвенно-подстилочная мезофауна типичных для бореальной зоны болотных, луговых и лесных экосистем в пределах зоны добычи и складирования отходов калийно-магниевых солей. Среди изученных болотные сообщества, суходольный луг и все лесные биогеоценозы не испытывают нагрузки засолением, чего нельзя сказать о пойменных лугах. Необходимо подчеркнуть, что засоление почв выражено в долинах малых рек, а источниками засоления являются места складирования твердых и жидких отходов калийных предприятий.
Пойменные луга, подверженные техногенному засолению, выделяются среди изученных сообществ и отличаются от пойменных лугов Пермского края, не испытывающих такого воздействия. Прежде всего, пойменные засоленные луга зарастают устойчивыми к засолению местными прибрежно-водными и влаголюбивыми видами растений, в том числе факультативно-галофитными. В то же время здесь нередки обычные сорно-луговые виды. Суходольные луга не подвержены засолению.
Мезофауны пойменных лугов, имеющих ту или иную степень засоления (пл. 4–5), в отличие от мезофауны суходольного разнотравного луга (пл. 3) и от остальных изученных стаций, демонстрируют низкое таксономическое разнообразие и наименьшие показания плотности населения и биомассы беспозвоночных. Структура мезофауны данных лугов оказалась крайне упрощена. В ней отсутствуют почвенные черви Enchytraeida и Lumbricidae, личинки жуков-щелкунов (Elateridae), многоножки (Chilopoda) и мн. др. беспозвоночные (см. табл. 1). Особенно обращает на себя внимание отсутствие каких-либо представителей мезофауны в почвенном слое пойменного засоленного луга (пл. 5), что говорит о наличии здесь неподходящих условий для почвенных обитателей (см. табл. 1).
Похожие результаты были получены при изучении влияния естественного засоления на почвенную фауну в степной зоне Центральной Азии [Брагина, 2004], Южного Зауралья [Нагуманова, 2007] и Забайкалья [Хобракова и др., 2015], которые показали, что засоленные почвы отличаются значительным обеднением видового состава и сокращением численности обитающих в них беспозвоночных животных.
Как мы отмечали ранее [Efimik et al., 2024], изученные верховые болота типичны для региона по своему флористическому составу и почвенно-подстилочной мезофауне. При этом осушаемое верховое болото находится на стадии трансформации растительности, что выражается в зарастании подростом березы и в меньшей выраженности моховой подстилки (0.85 ± 0.1 см по сравнению с 10.3 ± 0.8 см на верховом осоково-пушицево-сфагновом болоте). Мезофауна осушаемого болота беднее таковой верхового болота в целом, а население малочисленнее. На изученных болотах также отличается структура населения беспозвоночных подстилки и верхнего почвенного слоя (рис. 2–3). Засоление в данных типах болот не выявлено.
Все обследованные леса типичны для нашей зоны и на территории исследования вторичны. Различия во флористическом составе связаны с возрастом древостоя, составом древесного яруса, типом почвы и характером увлажнения. Учетные площадки в лесах выбирались на разном удалении от мест добычи и хранения солевых отходов, тем не менее, в связи с промывным водным режимом бореальной зоны, засоление почв в лесных экосистемах отсутствует.
По структуре мезофауна лесов отличается от луговой и болотной более сложной структурой, в которой ядром являются в подстилке многоножки-костянки, пауки и насекомые, в почвенном слое – дождевые черви, многоножки-костянки и насекомые. При этом численность лесных беспозвоночных не очень высокая и даже уступает таковой на верховом болоте (пл. 1) и на суходольном лугу (пл. 3).
Корреляционный анализ количественных показателей почвенной мезофауны с химическими свойствами почв показал следующее. Высокая отрицательная корреляция по шкале Р.Е. Чеддока [Котеров и др., 2019] между значениями Cl-, Ca2+, Mg2+ почв и значениями плотности (численности) всех беспозвоночных, и заметная отрицательная корреляция между значениями тех же ионов и плотности насекомых верхнего почвенного слоя (табл. 3) свидетельствует о чувствительности почвенных беспозвоночных и, особенно, насекомых к увеличению концентрации токсичных солей, формирующих хлоридное и магние-во-кальциевое засоление почвы.
Также была отмечена заметная отрицательная корреляция между плотностью (численностью) всех беспозвоночных с такими почвенными показателями, как гидролитическая кислотность (ГК), количество органического вещества, EKO (табл. 3), что свидетельствует о чувствительности беспозвоночных к увеличению кислотности почвы и содержанию органического вещества. Такую же чувствительность к почвенной органике демонстрируют многоножки (Chilopoda) (табл. 3).
Таблица 3
Коэффициенты корреляции плотности (N) и биомассы (B) почвенной мезофауны с химическими свойствами почв
[Correlation coefficients of density (N) and biomass (B) of soil mesofauna with chemical properties of soils]
|
Таксон |
EX О ca К |
о К ft |
о к |
б |
+ к! |
+ ад |
+ св |
й и « о а я О |
О W |
||
|
Lumbricidae (N) |
0.03 |
-0.13 |
0.33 |
-0.43 |
-0.38 |
-0.46 |
-0.29 |
0.14 |
-0.23 |
-0.22 |
-0.44 |
|
Lumbricidae (B) |
0.03 |
-0.06 |
0.16 |
-0.34 |
-0.28 |
-0.36 |
-0.17 |
0.37 |
-0.11 |
-0.05 |
-0.26 |
|
Enchytraeida (N) |
-0.23 |
-0.22 |
0.42 |
-0.14 |
-0.15 |
-0.15 |
-0.16 |
-0.18 |
-0.11 |
-0.22 |
-0.23 |
|
Enchytraeida (B) |
-0.23 |
-0.22 |
0.42 |
-0.14 |
-0.15 |
-0.15 |
-0.16 |
-0.18 |
-0.11 |
-0.22 |
-0.23 |
|
Gastropoda (N) |
-0.23 |
-0.22 |
0.42 |
-0.14 |
-0.15 |
-0.15 |
-0.16 |
-0.18 |
-0.11 |
-0.22 |
-0.23 |
|
Gastropoda (B) |
-0.23 |
-0.22 |
0.42 |
-0.14 |
-0.15 |
-0.15 |
-0.16 |
-0.18 |
-0.11 |
-0.22 |
-0.23 |
|
Chilopoda (N) |
-0.19 |
-0.36 |
0.15 |
-0.43 |
-0.42 |
-0.45 |
-0.41 |
-0.32 |
-0.41 |
-0.52 |
-0.62 |
|
Chilopoda (B) |
-0.14 |
-0.36 |
0.04 |
-0.46 |
-0.45 |
-0.48 |
-0.43 |
-0.34 |
-0.44 |
-0.54 |
-0.66 |
|
Diplopoda (N) |
0.16 |
0.14 |
0.35 |
-0.25 |
-0.19 |
-0.26 |
-0.03 |
0.52 |
0.01 |
0.21 |
-0.10 |
|
Dipljpjda (B) |
-0.03 |
-0.03 |
0.43 |
-0.20 |
-0.17 |
-0.21 |
-0.08 |
0.21 |
-0.03 |
0.02 |
-0.14 |
|
Aranei (N) |
0.28 |
0.24 |
0.31 |
-0.13 |
-0.23 |
-0.13 |
0.46 |
-0.17 |
0.07 |
0.14 |
0.48 |
|
Aranei (B) |
0.54 |
0.53 |
0.58 |
0.00 |
-0.09 |
-0.02 |
0.69 |
-0.09 |
-0.03 |
0.12 |
0.54 |
|
Insecta (N) |
0.22 |
-0.07 |
0.04 |
-0.62 |
-0.65 |
-0.66 |
-0.07 |
-0.29 |
-0.53 |
-0.49 |
-0.42 |
|
Insecta (B) |
0.02 |
-0.15 |
-0.39 |
-0.20 |
-0.20 |
-0.21 |
-0.22 |
-0.23 |
-0.27 |
-0.29 |
-0.36 |
|
Общее количество |
0.16 |
-0.14 |
0.24 |
-0.71 |
-0.74 |
-0.75 |
-0.17 |
-0.28 |
-0.54 |
-0.53 |
-0.52 |
|
Общая биомасса |
0.04 |
-0.16 |
-0.23 |
-0.40 |
-0.37 |
-0.42 |
-0.30 |
0.03 |
-0.32 |
-0.29 |
-0.48 |
Примечание: 0.5–0.7 – заметное значение, 0.7–0.9 – высокое значение корреляции по шкале Чеддока [Котеров и др., 2019].
Есть данные китайских ученых [Wu et al., 2015], которые также показали, что плодородие почвы положительно влияет на почвенные организмы как в засоленных, так и в незасоленных почвах, но стоит отметить, что они изучали засоление морскими солеными водами на морском побережье.
С изменением значений биомассы пауков была отмечена заметная положительная корреляция со многими почвенными показателями – pHвод, pHсол, ионами HCO 3 -, Na+, а также EKO (табл. 3), что труднообъяснимо, т. к. пауки не относятся к почвенным обитателям, попадание в почвенные пробы чаще всего происходит при сборе подстилки.
Заключение
Большинство изученных нами экосистем не испытывают нагрузки техногенным засолением в связи с промывным водным режимом бореальной зоны. Засоление выражено лишь в поймах малых рек, где формируются техногенные солончаковые почвы, которые оказывают непосредственное влияние как на флористический состав, так и на состав и структуру почвенной мезофауны.
В настоящее время солеотвалы и шламохранилища на территории Верхнекамского месторождения солей занимают более 1 000 га, а с учетом перспективного развития калийного производства их площади могут достигнуть 2–3 тыс. га [Бобошко, Бачурин, 2004]. Следовательно, при сохранении современных технологий солевая нагрузка на экосистемы Прикамья возрастет.
Пойменные луга малых рек подвержены воздействию высокоминерализованных сточных вод калий-но-магниевых предприятий, они резко выделяются по типу почв, флористическому составу, а также по структуре фауны и населения беспозвоночных подстилки и верхнего слоя почвы. Формирование солончаковых почв в данных биотопах обусловлено близким расположением мест хранения отходов и площадной разгрузкой засоленных подземных вод.
Именно в таких условиях техногенное засоление проявляется особенно сильно и непосредственно влияет на все живые организмы сообществ. В условиях таких почв заметно изменяется видовой состав фитоценозов – наблюдается сокращение количества видов, типичных для пойменных лугов бореальной зоны. Их замещают растения, толерантные к засолению: прибрежно‑водные, влаголюбивые (в том числе факультативно‑галофитные) и сорно‑луговые.
Мезофауна пойменных лугов имеет низкое таксономическое разнообразие и бедное население беспозвоночных. Мезофауна солончаковых почв пойменных лугов упрощена и обеднена за счет отсутствия типичных почвенных обитателей (Enchytraeida, Lumbricidae, Elateridae, Chilopoda и др.). Следует отме- тить, что специализированных, устойчивых к засолению (галофильных) видов беспозвоночных не обнаружено.
Результаты корреляционного анализа количественных характеристик почвенной мезофауны с химическими свойствами почв показали, что беспозвоночные, особенно насекомые, снижают свою численность при увеличении концентрации токсичных солей и кислотности в почвах.
