Оценка степени генетической дифференциации маточного поголовья красноярского типа черно-пестрой породы в СПК «Алексеевский»

Автор: Еремина И.Ю.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Ветеринария и животноводство
Статья в выпуске: 2, 2014 года.
Бесплатный доступ
Изучены генетические характеристики черно-пестрой породы в СПК «Алексеевский» по ЕАВ-локусу групп крови. Проанализирована структура популяции красноярского типа за последние десятилетия.
Маркеры, группы крови, селекция, молочный скот, полиморфизм, черно-пестрая порода коров
Короткий адрес: https://sciup.org/14083516
IDR: 14083516 | УДК: 575.174.015.3:
The assessment of the genetic differentiation degree of the uterine livestock of the Krasnoyarsk type black and motley breed in the agricultural production cooperative "Alekseevskiy"
The genetic characteristics of black and motley breed in APC "Alekseevskiy" on EAВ-blood group locus are studied. The population structure of the Krasnoyarsk type for the last decade is analyzed.
Текст научной статьи Оценка степени генетической дифференциации маточного поголовья красноярского типа черно-пестрой породы в СПК «Алексеевский»
Введение. Членство России во Всемирной торговой организации влечет за собой неизбежные изменения во многих производственных отраслях, включая сельское хозяйство. Экономические изменения не могут пройти бесследно, в том числе и для молочного скотоводства. Поскольку это весьма чувствительный производственный сегмент, в котором уже сегодня весьма напряженная ситуация: слабая степень дотаций на животноводческую продукцию – с одной стороны, и сложные климатические, социальные и т.п. условия ведения животноводства, повышающие себестоимость продукции, – с другой стороны. Импорт продукции чреват последствиями и другого уровня: продукция длительного срока хранения, произведенная в иных климатических и экологических зонах с применением ГМО кормовых культур, отразится на здоровье населения. А это напрямую затрагивает продовольственную безопасность страны.
В свете вышесказанного, для сохранения и развития конкурентоспособного молочного животноводства, актуальными остаются вопросы состояния кормовой базы, условий содержания, обеспечивающих оптимальное функционирование всех систем организма крупного рогатого скота, и технического оснащения
[1]. Все это относится к условиям реализации генетического потенциала, т.е. внешней среде, определяющей фенотип животного. При этом первичные изменения происходят на уровне генотипа, что обусловливает актуальность контроля его состояния и его целенаправленное изменение.
Выбор оптимальной породы (типа, линии) для осуществления поставленных целей, формирование структуры стада, обеспечивающей наиболее эффективное ведение хозяйства; разработка систем разведения, обеспечивающих продуктивное долголетие животных; обеспечение надежных методов селекции и мониторинга – вот далеко не полный круг вопросов, отражающих состояние селекционной работы, ее уровень.
В современных условиях, когда требования к объёмам производства и срокам столь высоки, нет необходимости объяснять несостоятельность селекционной работы, базирующейся исключительно на традиционных приемах. Для обеспечения наибольшего успеха необходимо привлечение разнообразных методов оценки и коррекции как генотипа, так и фенотипа. Требуются интегральные методы, основанные на современных достижениях генетики, биохимии и других биологических наук для своевременной оценки генофонда и мониторинга протекающих селекционных изменений. Применение селекции при помощи маркеров (Marker Assisted Selection – MAS) дает возможность селекционеру исключить факторы действия среды на признаки из оценки животного и пользоваться лишь генетической составляющей фенотипа [2].
В качестве маркеров используются только природные комплексы генов, характерные для данного вида животных. Эти комплексы прошли через сито естественного отбора у предков домашних животных, поэтому их присутствие в геноме животных является естественным и безопасным как для самого животного, так и для человека, потребляющего от него продукцию. Существует ряд требований к маркерам: 1) фенотипические проявления аллельных вариантов должны быть доступны для идентификации у различных особей; 2) изучаемые локусы должны представлять случайную выборку генов в отношении их физиологических эффектов и степеней изменчивости; 3) маркеры должны обладать лёгкой выявляемостью и воспроизводимостью; 4) получаемые данные должны быть сопоставимы в разных лабораториях; 5) маркеры должны обладать относительной нейтральностью [2, 3].
Большинство исследований свидетельствуют о том, что полиморфные белки отвечают перечисленным требованиям и могут использоваться в качестве эффективных молекулярно-генетических маркеров в селекции организмов. В качестве молекулярных маркеров животных могут использоваться системы групп крови [4–7].
Цель исследования. Использовать полиморфные белки ЕАВ-локуса в качестве маркеров для оценки степени генетической дифференциации маточного поголовья черно-пестрой породы в СПК «Алексеевский».
В соответствии с этим были поставлены следующие задачи : 1) изучить генетические характеристики черно-пестрой породы в СПК «Алексеевский»; 2) проанализировать структуру аллелофонда по ЕАВ-системе групп крови животных красноярского типа на примере стада СПК «Алексеевский»; 3) провести генетический мониторинг аллелофонда породы за последние десятилетия.
Мониторинг генетической структуры, уровня полиморфности типов черно-пёстрого скота Сибири, включая красноярский, приобский и кемеровский, проводили в 1993 и 2010 годах раздельно по маточным группам и быкам-производителям.
Характерная картина распределения частот встречаемости антигенов маточного поголовья СПК «Алексеевский» представлена на рисунке 1.
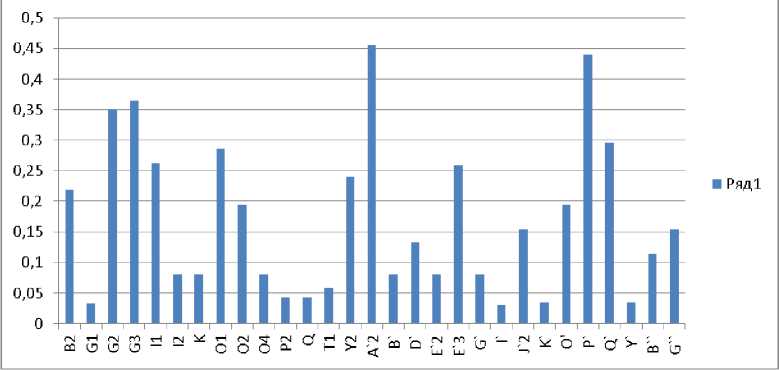
Рис. 1. Антигенный спектр ЕВА-локуса маточного поголовья стада СПК «Алексеевский»
Следует отметить невысокую насыщаемость ЕАВ-локуса антигенами, наивысшие показатели частоты менее 50 %, 13 антигенов имеют частоту менее 10 %.
Сопоставляя частотный спектр маточного поголовья в ретроспективе (с 1993 г.), выявлили 7 общих антигенов с частотой менее 10 %: I', G1, У, K', T1, B', K. У животных, исследованных в 1993 году, также невысокие значения частот антигенов (до 50 %). Значительно отличается антигенофонд по ЕАВ-локусу у быков-отцов (2010 г.) (рис. 2).
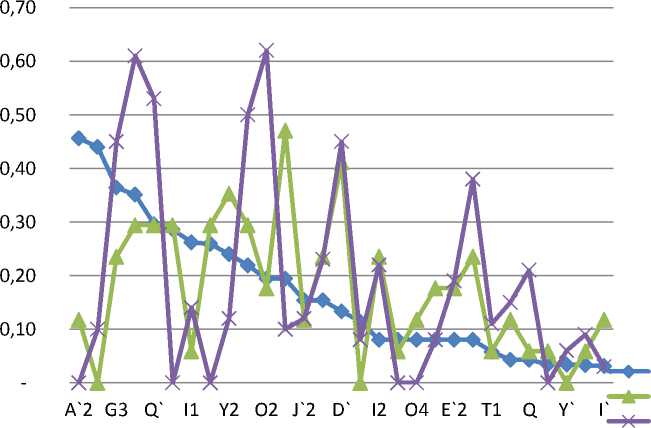
Красноярский 2010 маточное
Красноярский 2010 быки
Красноярский 1993 маточное
Рис. 2. Сравнительная частотная характеристика ЕВА-локуса групп крови черно-пестрого скота
Для анализа в общем спектре выделяли три группы антигенов: часто встречающиеся, накапливающиеся (40-100 %); редко встречающиеся (21-39 %); очень редко встречающиеся, элиминирующиеся (0-20 %) (табл.1).
Сравнительная характеристика частот антигенных факторов крови ЕАВ-локуса у коров черно-пестрой породы разных типов
Таблица 1
|
Группа |
Антигены, % |
||
|
Часто встречающиеся |
Элиминирующиеся |
Редко встречающиеся |
|
|
40-100 |
0-20 |
21-39 |
|
|
Красноярский 2010 |
A'2, P' |
O', O2, G'', J'2, D', B'', G', I2,E'2, B', O4, K, T1, P2, Q, K',Y', G1, I' |
G3, G2, Q', O1, I1, E'3, Y2, B2 |
|
Красноярский 1993 |
O2, G2, Q', B2, G3, D' |
E'2, P2, I1, Y2, J'2, T1, O', P', G1, B', B'', Y', I' |
G', G'', I2, Q |
|
Новосибирский |
Q', Y2, B2, G3 |
O1, K, D', Q, I1, J'2, T1, Y', B', P2, B'', I', G1 |
E'2, G2, G'', I2, O2, G', O', P' |
|
Кемеровский |
G2, B2, Y2, Q' |
J'2, A'2, O2, O1, K', B', B'' |
Q', E'2, O' |
При сопоставлении полученных результатов с литературными данными [8] были выявлены особенности у представителей разных типов черно-пестрого скота. Группы часто встречающихся и редко встречающихся антигенов существенно различаются у разных типов черно-пестрого скота. Наименьшее число распространённых антигенов - у животных красноярского типа (A'2, P'), при том, что изначально (1993) этот тип был насыщен пятью антигенами (O2, G2, Q', B2, G3, D) и с большей частотой встречаемости. Группа элиминирующихся антигенов у исследуемых групп обнаружила сходство по восьми антигенам: O', J`2, B``, E`2, B', T1, G1, I' (т.е. сходство на 28 % по антигенам ЕАВ-локуса), при этом данная группа наиболее наполнена антигенами, поэтому названные антигены можно считать маркерными. У животных красноярского типа группа редко встречающихся антигенов - наибольшая по числу факторов, прирост количества антигенов группы составил 20 % к 2010 г. по сравнению с 1993 г.
В дальнейшем изучен аллелофонд маточного поголовья красноярского типа черно-пёстрого скота, n=324 (табл. 2 ).
Аллелофонд ЕАВ-локуса животных Красноярского типа (n=324)
Таблица 2
|
Аллели |
Частота |
Аллели |
Частота |
Аллели |
Частота |
|
Y 2 |
0,1990 |
G 3 O i O 2 |
0,0062 |
B 2 G‘ G" |
0,0015 |
|
Q′ |
0,1296 |
G‘ G 3 O i O 2 Y‘ G" |
0,0062 |
E 2 ’ J 2 ’ O’ Y‘ |
0,0015 |
|
A 2 ‘ |
0,0803 |
A 2 ’ G" |
0,0154 |
B 2 D‘ B"Q |
0,0015 |
|
G″ |
0,0787 |
G 2 G 3 E 2' |
0,0124 |
B 2 O i y 2 O’ |
0,0015 |
|
g 2 g 3 |
0,0771 |
G 2 G 3 E 2' Q′ |
0,0108 |
Y 2 G‘ Y‘ G" |
0,0015 |
|
I 2 |
0,0401 |
A 2 ’ B 2 E 3 ’ O 3 |
0,0108 |
G 3 T i |
0,0015 |
|
A 2 ‘ O i |
0,0355 |
G 2 G 3 Y 2 E 2' Q′ |
0,0062 |
O i O 2 Q |
0,0015 |
|
G 2 G 3 Y 2 E 3' Q′ |
0,0340 |
A 2 ’ G" |
0,0046 |
P 2 O 2 G‘ Y‘ |
0,0015 |
|
O i O 2 J 2 ’ O’ |
0,0247 |
B 2 O i O 2 |
0,0046 |
A 2 ‘G 3 K O i Q Q‘ Е з ‘ P‘ J 2 ’ O’ |
0,0015 |
|
B 2 B" D‘ |
0,0247 |
Y 2 G‘ G" |
0,0031 |
I i Q‘ G’G" |
0,0015 |
|
A 2 ’ B 2 E 3 ’ |
0,0244 |
B 2 B‘ |
0,0031 |
I i G 3 Q’ Q Е з ‘ |
0,0015 |
|
B 2 B" B’O i |
0,0244 |
B 2 O i J 2 ’ O’ |
0,0031 |
G 3 I i K O Q‘ G’G" |
0,0015 |
|
b |
0,0185 |
G i |
0,0015 |
||
|
I′ |
0,0170 |
D′ |
0,0015 |
Выявлены сорок феногрупп, встречающихся у этих животных. Наивысшая частота выявлена у моно-и дифакторных феногрупп: Y2, Q`, A`2, G``, G2 G3 – и составляет 0,199–0,077. Условно маркерными для красноярского типа можно считать следующие: Y2, Q′, A2′, G″, G2 G3, I2, A2′ O1 , G2G3Y2E3' Q′, O1 O2 J2′ O′, B2 B″ D′, A2′ B2E3′, B2 B″ B′O1, b, I′, A2′ G″.
Сравнительный анализ полиморфизма у животных красноярского и приобского типов черно-пестрого скота выявил ряд особенностей (табл.3).
Частоты аллелей групп крови черно-пестрой породы по ЕАВ-локусу
Таблица 3
|
Аллели |
Красноярский тип |
Приобский тип |
Черно-пестрый голшт.1/2 |
|
|
1993 (7300) |
2010 (324) |
2010 (621) |
2000 (333) |
|
|
G 2 G 3 Y 2 E 2 ' Q′ |
0,3340 |
0,0062 |
0,1739 |
0,2650 |
|
O 2 T. 2 D‘ |
0,0833 |
|||
|
G 2 O 2 Q Q‘ G" |
0,3160 |
|||
|
G″ |
0,0700 |
0,0787 |
0,0161 |
0,2192 |
|
y 2 |
0,0800 |
0,1990 |
0,0701 |
|
|
O′ |
0,0600 |
0,0300 |
||
|
B 2 |
0,0600 |
0,0080 |
||
|
G 2 G 3 |
0,0370 |
0,0771 |
||
|
B 2 O’ |
0,0550 |
|||
|
Р 2 |
0,0200 |
0,0032 |
||
|
I 2 |
0,0800 |
0,0401 |
0,1450 |
0,3513 |
|
D′ |
0,0660 |
0,0015 |
0,0090 |
|
|
«b» |
0,0200 |
0,0185 |
0,0370 |
0,0240 |
Современный аллелофонд по изучаемой системе имеет пять общих феногрупп, т.е. около 40 % идентичности у сравниваемых типов; по негативному «b» аллелю частота одинаково низкая. Частота аллеля G2G3Y2E2'Q' у красноярского типа еще ниже (0,006), что в 30 раз ниже, чем у приобского типа (0, 174), при этом изначально эта аллель встречалась с частотой в 50 раз выше (0,334). По аллелю G″ – частота у красноярского типа остается стабильно невысокой (0,070), для приобского типа она еще ниже (0,016). Аллель Y2, при сравнительно низкой концентрации у животных приобского типа (0,070), чаще встречается у живот- ных красноярского типа (0,199), с накоплением по сравнению с состоянием на 1993 г. (0,080). Противоположная картина наблюдается по аллелю I2.
Выводы. Проведенный анализ полиморфизма групп крови коров черно-пестрой породы красноярского типа по антигенным факторам и аллелофонду ЕАВ-локуса подтвердил процесс консолидации генетического материала у представителей нового типа. Можно считать маркерными: невысокую насыщаемость ЕАВ-локуса антигенами; элиминируемость антигенов O', J`2, B``,E`2, B`, T1, G1, I'; присутствие аллелей Y2, Q′, A2′, G″, G2 G3, I2, A2′ O1 , G2G3Y2E3' Q′, O1 O2 J2′ O′, B2 B″ D′, A2′ B2E3′, B2 B″ B′O1, b, I′, A2′ G″. При стабильно низкой частоте «b» и G″ аллеля, снижается частота G2G3Y2E2'Q', I2 и D′ аллелей, повышается Y2 и G2G3 аллелей.
