Односторонняя межсортовая несовместимость отдельных образцов перца сладкого capsicum annuum l

Автор: Бландинская О.А., Козарь Е.Г., Беспалько Л.В., Балашова И.Т.
Журнал: Овощи России @vegetables
Рубрика: Современные направления селекции овощных культур
Статья в выпуске: 4 (21), 2013 года.
Бесплатный доступ
В статье обсуждается явление односторонней перекрестной несовместимости при внутривидовой гибридизации Capsicum annuum L., морфологические особенности ее проявления у сортов перца сладкого Белоснежка и Карлик при опылении пыльцой других образцов.
Перец сладкий, capsicum annuum l. пыльца, несовместимость
Короткий адрес: https://sciup.org/14025040
IDR: 14025040 | УДК: (635.649+635.646):631.523:575.18(06)
Irreciprocal inter-cultivar self-incompatibility of some samples of sweet pepper (capsicum annuum L.)
The irreciprocal self-incompatibility at site-specific hybridization of Capsicum annuum L. and its morphological features in the varieties of sweet pepper Belosnezhka and Karlik during pollination by other samples is described.
Текст научной статьи Односторонняя межсортовая несовместимость отдельных образцов перца сладкого capsicum annuum l
Вопросы опыления – оплодотворения играют важную роль в практической работе по гибридизации. Успех скрещивания во многом зависит от взаимодействия пыльцы отцовского растения с тканями пестика материнской формы и очень часто, несмотря на нормальное их развитие и благоприятные условия, опыление не приводит к завязыванию плодов и получению гибридных семян. Такие комбинации скрещивания относят к несовместимым [1-3]. Перец сладкий (Capsicum annuum L.) является факультативным самоопылителем и относится к самофертильным видам, растения которых, как правило, легко скрещиваются между собой. Обычно у этого вида перца сладкого трудности возникают в основном при получении межвидовых гибридов [4,5], однако, у сортов Белоснежка и Карлик из генофонда лаборатории селекции и семеноводства пасленовых культур ВНИИССОК обнаружено редкое явление односторонней внутривидовой (межсортовой) «нескрещиваемости» при их использовании в качестве материнского компонента. Изучая эти сорта, Е.А. Джос [5] отмечала, что их рыльца не способны воспринимать пыльцу других образцов кроме соб- ственной, что, по мнению автора, может свидетельствовать о наличии у них спорофитной несовместимости. В данной работе продолжено изучение данного явления и приведены результаты сравнительного анализа морфологических изменений в тканях пестика данных образцов в процессе опыления своей и чужеродной пыльцой.
Материалы и методы. Объектами исследования являлись образцы перца сладкого селекции ВНИИССОК – Белоснежка, Карлик, Желтый букет, Л-Фех, Л-70. Агротехника выращивания перца сладкого – общепринятая для ЦНЗР. Кастрацию проводили в фазе белого бутона (за 1-2 суток до раскрытия цветка). Опыление выполняли в день кастрации в утренние часы свежесобранной пыльцой с последующей изоляцией и маркированием опыленных цветков. Морфологическое изучение опыленных бутонов проводили в динамике (в течение 3 суток). Продольные срезы пестиков свеже-сорванных бутонов изучали под бинокулярной лупой CITOVAL 2 с фоторегистрацией Digital Camera for Microscope DCM 300. Полученные фотоматериалы анализировали с помощью специализированной програм-
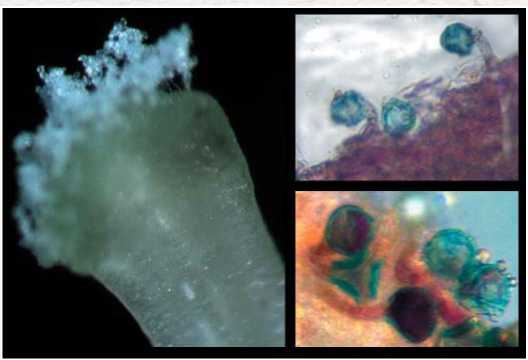
Рис.1. Прорастание пыльцы перца сладкого на рыльце пестика через 3 часа после опыления.
мы Scope Photo 3.0.
Результаты и обсуждение. Инициация процессов межклеточного узнавания и прорастания начинается сразу после адгезии пыльцы на поверхности рыльца и ее гидратации [6-8]. При этом, успех прорастания пыльцы и образование пыльцевой трубки (как in vivo , так и in vitro ) зависит от сбалансированности между водным запасом зрелой пыльцы после дегидратации и степенью оводненности поверхности рыльца пестика или среды [9]. У видов с «влажной» поверхностью рыльца гидратация пыльцы, как правило, начинается сразу же, после ее попадания в жидкий питательный матрикс (водно-липидная эмульсия). В других случаях обводнение наблюдается только после разрыва (лизиса) кутикулы и высвобождения секрета рыльца [10-13].
Растения семейства Пасленовые имеют влажные рыльца, секретирующие сахара, фенольные вещества, полисахариды, белки и липиды, что обеспечивает хорошую адгезию, стимулирует быстрое набухание и прорастание пыльцы. Далее пыльцевые трубки растут между клетками столбика, путем энзиматического растворения пектинового межклеточного матрикса с образованием канала внутри столбика, по которому пыльцевые трубки достигают завязи [5,7,8,14-16]. Начало прорастания пыльцевых трубок у сортов перца сладкого на поверхности рыльца наблюдали через 2-3 часа после нанесения пыльцы (рис.1).
У образцов Желтый букет, Л-Фех, Л-70, которые легко вступают в скрещивания с другими сортами, к этому времени происходит лизис проводниковой ткани у основания рыльца (рис.2) и формирование широкого канала (полости) по всей длине столбика, по которому пыльцевые трубки легко достигают завязи (на вторые сутки после опыления). При этом не отмечено существенных различий в изменении морфологии пестика у данных сортов при опылении своей или чужеродной пыльцой.
У сорта Белоснежка при опылении собственной пыльцой наблюдали аналогичные изменения в тканях пестика (рис.3), однако образование канала отмечали преимущественно в нижней части столбика (около 1/2 от общей длины). При опылении чужеродной пыльцой чаще всего наблюдали отсутствие образования проводящего канала, хотя пыльца на рыльце пестика прорастала. Это же отмечала в своих исследованиях и Е.А. Джос [5], которая показала, что в ряде комбинаций большинство чужеродных пыльцевых трубок, в отличие от опыления собственной пыльцой, было ориентировано в разные стороны, только не вовнутрь столбика (через 2 часа после опыления). При этом вместо лизиса проводниковой ткани столбика наблюдалось даже ее уплотнение, что затрудняло дальнейший рост трубок. Однако, как показали наши исследования, при опылении чужеродной пыльцой у сорта Белоснежка в отдельных случаях происходил частичный лизис проводниковой ткани (без образования четкого канала) и на вторые сутки в завязи отдельных пестиков отмечалось появление единичных пыльцевых трубок (рис.3). При этом успех оплодотворения и завязывания гибридных семян в данном случае в значительной степени зависел от соотношения на растении цветков, опыленных собственной (самоопыление) и чужеродной пыльцой. Наличие на растении даже единичных самоопыленных цветков в период формирования гибридных плодов, как правило, приводило к опадению последних на ранних стадиях развития, по-видимому, ввиду низкой их осемененности. На осыпание плодов вследствие ограниченного количества семян, как одну из причин неудач при межвидовых скрещиваниях, указывали и другие авторы [17, 18]. Полное удаление на материнском растении цветов и бутонов, не участвующих в скрещивании, у сорта Белоснежка в ряде комбинаций способствовало завязыванию и созреванию части гибридных плодов, даже при небольшом количестве семян (отсутствует конкуренция, возникающая между «самоопы-
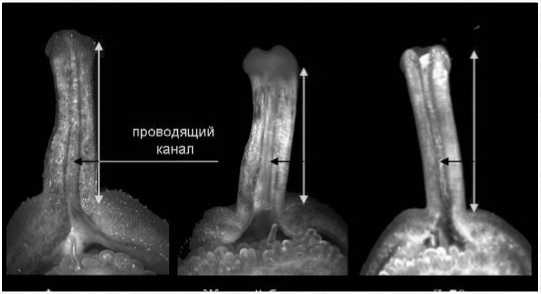
Рис.2. Образование проводящего канала в столбике пестика у сортов перца сладкого Фехерезон, Желтый букет и Л-70 через 3 часа после опыления.
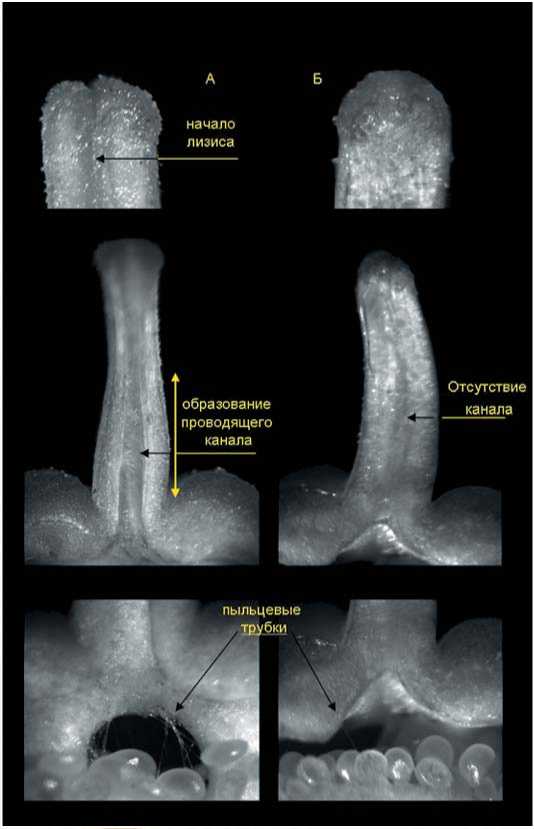
Рис. 3. Продольный срез пестика цветков сорта Белоснежка после опыления собственной (А) и чужеродной пыльцой (Б): разрез верхней части столбика в первые сутки опыления, образование канала прорастание пыльцевых трубок в завязь на 2 сутки после опыления.
ленными» и гибридными плодами).
Изучение перекрестной несовместимости сорта перца сладкого Карлик, показало, что одной из причин этого явления также является трудность прорастания пыльцевых трубок в тканях его столбика. В отличие от Белоснежки и других сортов, у него практически не происходило образования открытого канала, даже при опылении собственной пыльцой (рис.4). Пыльцевые трубки росли внутри плотного матрикса проводящей ткани столбика, которая при самоопылении постепенно лизировалась и частично разрыхлялась. При этом основная масса пыльцевых трубок достигала завязи преимущественно на 3 сутки после опыления.
В случае нанесения чужеродной пыльцы этого не происходило, хотя и наблюдалось прорастание части пыльцевых зерен на рыльце пестика. При этом в большинстве изученных комбинаций скрещивания предотвращение самоопыления путем удаления неопыленных цветков и бутонов материнского растения, как правило, не приводило к завязыванию гибридных плодов (исключение составила лишь комбинация Карлик х Белоснежка). Торможение роста пыльцевых трубок в тканях столбика сорта Карлик, у которого совместимость пыльцы и пестика, по-видимому, строго детерминируется генотипом конкретного материнского растения, также может быть одной из причин сложности получения семян при внутрисортовых скрещиваниях в рамках проведения сортоулучшающей селекции.
Заключение
Считается, что перекрёстная несовместимость, как и самонесовместимость, генетически детерминированы. У гомоморфных растений процессы, препятствующие оплодотворению, происходят между пыльцой и тканями пестика, причем реакция при гаметофитной несовместимости определяется гаплоидным генотипом пыльцы (состоянием множества аллелей генов несовместимости). Пыльца прорастает, но пыльцевые трубки, начавшие путь по столбику, через определенное время прекращают свой рост. Этот тип характерен для семейств Solanaceae (томат, паслен), Rosaceae (яблоня, роза), Fabaceae (бобы) и др. Реакция пыльцы со спорофитным контролем несовместимости определяется генотипом диплоидного материнского растения
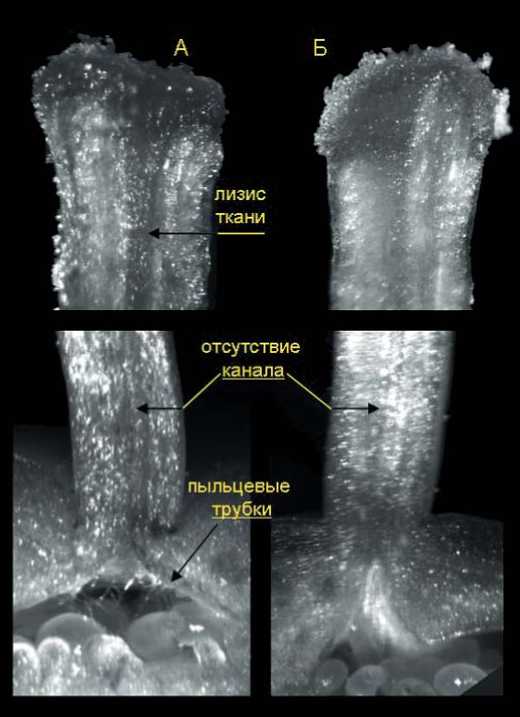
Рис.1. Прорастание пыльцы перца сладкого на рыльце пестика через 3 часа после опыления.
и зависит от взаимодействия аллелей генов несовместимости. Она протекает обычно в форме подавления прорастания пыльцевых зерен на рыльце столбика, которые не прорастают совсем или в малом количестве лишь частично проникают в ткани столбика. Этот тип несовместимости характерен для семейств Brassicaceae (капуста, редька), Asteraceae (скерда) и другие [14].
Однако разделение видов на гаметофитные и споро-фитные в отношении контроля несовместимости является весьма относительным. У некоторых видов представлено сочетание отдельных характеристик этих систем [14,19], что было отмечено также и в наших исследованиях при изучении явления внутривидовой односторонней несовместимости у сортов перца сладко- го Белоснежка и Карлик. Хотя механизм этого явления пока еще до конца не ясен, но можно предположить, что избирательность оплодотворения у сортов Белоснежка и Карлик во многом обусловлена морфофизиологическими особенностями строения их пестика, характером взаимной стимуляции между пыльцевыми зернами, клетками рыльца и столбика на ранних этапах прорастания, особенностями метаболизма материнского растения (внутренний отбор), что было отмечено рядом исследователей и на других культурах [2,3]. Анализ литературы и приведенные в данной работе экспериментальные данные свидетельствуют о том, что односторонняя несовместимость сортов Белоснежка и Карлик не является абсолютной и ее можно преодолеть различными способами [5, 20, 21].
Список литературы Односторонняя межсортовая несовместимость отдельных образцов перца сладкого capsicum annuum l
- Johri B.M., Ambegaokar K.B., Srivastava P.S. Comparative Embryology of Angiosperms, n.1. SpringerVerlag, 1992.-1221 р.
- Методические указания по гаметной селекции сельскохозяйственных растений. -Москва, 2001. -391с.
- Ковалева Л.В., Захарова Е.В. Гаметофитно-спорофитные взаимодействия в системе пыльца-пестик. Гормональный статус и механизм самонесовместимости. Физиология растений. 2004.-Т. 51.-№ 3.-С. 446-451.
- Бухарова А.Р. Получение исходного материала перца на основе межвидовых скрещиваний: Автореф. дис. канд.с.-х. наук. М., 1995. -22 с.
- Джос Е.А. Создание исходного материала перца с использованием отдаленной межвидовой гибридизации: Автореф. дис. канд.с.-х. наук. М., 2009. -24 с.
- Суриков И.М. Морфологические, генетические и эволюционные аспекты теории внутривидовой и межвидовой половой несовместимости у цветковых растений/Генетико-селекц. аспекты систем размножения энтомофил. видов растений, 1987. -С. 3
- Konar R.N, Linskens H.F. The morphology and anatomy of the stigma of Petunia hybrida.Planta. 1966a; 71: 356371.
- Zinkl, G.M., Preuss, D. Dissecting Arabidopsis pollenstigma interactions reveals novel mechanisms that confer mating specificity. Ann. Bot.85, 2000. -Р. 15-21.
- Kranz, E., L?rz, H. Micromanipulation and in vitro fertilization with single pollen grains of maize. Sex. Plant Reprod. 3, 1990. -Р. 160-169.
- Цингер Н.В., Петровская-Баранова Т.П. Оболочка пыльцевого зерна -живая физиологически активная структура. -ДАН СССР, 1961.-138 с.
- Heslop-Harrison J. Pollen walls as adaptive systems. Annals of the Missouri Botanic Garden 66: 813-829.
- Hoekstra F.A., Bruinsma J. Protein synthesis of binucleate and trinucleate pollen and its relationship to tube emergence and growth. Planta. -1979. -559-566 р.
- Heslop-Harrison Y., Heslop-Harrison J. The digestive glands of Pinguicula: structure and cytochemistry. Ann.Bot. 1981. -47: 293-319.
- Суриков И.М. Несовместимость и эмбриональная стерильность растений/И. М. Суриков. ВАСНИЛ. -М.: Агропромиздат, 1991. -220 с
- Cochran H. A morphological study of flower and seed development in pepper. Journal of Agricultural Research. 1938. -№56. -Р. 395-419.
- Hu Shi-yi; Xu Li-yun Transmitting tissue and pollen tube growth in sweet pepper Capsicum frutescens var. grossum//Acta bot. sinica. -1985. -Т. 27. -N 5. -P. 449454.
- Бриггс Ф., Ноулз П. Научные основы селекции растений. -М.: Колос, 1972. -397 с.
- Бухарова А.Р., Бухаров А.Ф. Отдаленная гибридизация овощных пасленовых культур. -Мичуринск: МичГАУ. -2008. -274 с.
- Lawrence M.J., Marshall D.F., Curtis V.E., Fearon C.H. Gametophytic self -incompatibility re -examined: a repli. Biological Reviews, 1945.-V. 54-№I.-P.131-138.
- Козарь Е.Г., Бландинская О.А., Балашова И.Т., и др. Использование стероидных гликозидов и термической обработки пыльцы при гибридизации перца сладкого//Межд. научно-практ. конф. Состояние и перспективы научных исследований по картофелеводству, овощеводству и бахчеводству. -Алма-Ата, 2011. -С. 43-50.
- Козарь Е.Г., Беспалько А.В., Бландинская О.А., Балашова Н.Н. Применение холодового стресса в селекции перца сладкого по микрогаметофиту. Мат. межд. конф. (24-25 октября)/Кишинев, 2013. -С. 41.
