Онтогенетическая структура ценопопуляций Iris humilis Georgi в условиях Витимского плоскогорья и Восточного Саяна

Автор: Халтанова Елена Петровна
Журнал: Вестник Бурятского государственного университета. Философия @vestnik-bsu
Рубрика: Биология: ботаника
Статья в выпуске: 4, 2013 года.
Бесплатный доступ
В работе приводятся результаты изучения 6 ценопопуляций Iris humilis Georgi в условиях Витимского плоскогорья и Восточного Саяна. По результатам исследования был выявлен базовый онтогенетический спектр, а также его связь с особенностями развития в онтогенезе.
Онтогенез, онтогенетическая структура, ценопопуляции
Короткий адрес: https://sciup.org/148181881
IDR: 148181881 | УДК: 581.1
Ontogenetic structure of Iris humilis Georgi coenopopulations in the conditions of Vitim plateau and East Sayan
The results of research of ontogenetic structure of Iris humilis Georgi coenopopulations in the conditions of the Vitim Plateau and East Sayan are presented in the article. As results the basic ontogenetic spectrum and its association with a pattern of development of individuals in ontogeny were determined.
Текст научной статьи Онтогенетическая структура ценопопуляций Iris humilis Georgi в условиях Витимского плоскогорья и Восточного Саяна
Iris humilis Georgi представляет собой многолетний летнезеленый дерновинный симподи-ально нарастающий короткокорнсвищный поли-карпик с нарастающим стелющимся побегом [2]. Имеет евроазиатский ареал. Встречается в Европе, на юге советского Дальнего Востока. Монголии. Северо-Западном Китае |13| На территории России произрастает в Западной и Восточной Сибири (рис I). Ареал I. humilis достаточно обширный, но не сплошной, а во многих районах этот вид является редким [1|. Так. например. данный вид занесен в Красные книги Омской и Тюменской областей как редкий, встречающийся в немногих местах. Многие популяции находятся под защитой заповедников Азас. Байкало-Ленский, Байкальский. Баргузинский. Сохондинский, Убсунурский.
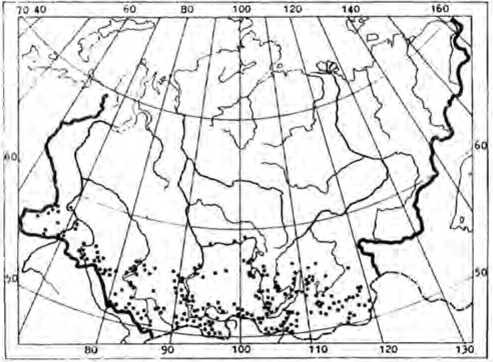
Рис. 1. Apca;i /. humilis в России
/. humilis приурочен к степным, нередко каменистым склонам, произрастает в борах, на лесных полянах и прибрежных лугах. Изученные ценопопуляций /. humilis располагались в основном на хорошо прогреваемых участках южной экспозиции в петрофитных и луговых степях. Исследование онтогенеза и популяционной структуры ириса низкого ранее нс проводилось. и имеются лишь описания генеративных особей [3 ].
Материалы и методы
Материал для изучения онтогенетической структуры ириса низкого был собран на территории Витимского плоскогорья и Восточного Саяна.
Определение возрастных спектров ценопопуляций (ЦП) ирисов проводили по общепринятым методам и принципам, разработанным ТА. Работновым [10], дополненным и модифицированным АА. Урановым [11] и его учениками [6; 7]. Для изучения и определения онтогенетических состояний закладывались учетные площадки 1м2 на каждый третий шаг вдоль и поперек склона. На площадках проводился подсчет особей всех возрастов. При выделении счетной единицы в ЦП учитывали жизненную форму растений. Поскольку I. humilis имеет неявнополицентрическую биоморфу [7], то за счетную единицу на начальных стадиях онтогенеза при нималась отдельная особь, с началом вегетативного размножения - партикула. Для характеристики ценопопуляций использовались классификации А.А. Уранова и О.В. Смирновой [12] и Л.А. Животовского [4]. Для характеристики онтогенетической структуры ценопопуляций I. humilis - различные демографические показатели: экологическая плотность [8], эффективная плотность [4], возрастность [10]. Тип характерного онтогенетического спектра ценопопуляций установлен согласно представлениям Л.Б. За-угольновой [5].
Онтогенетическая структура ценопопуляций I. humilis изучалась в петрофитно-степных сообществах и лугово-степных сообществах, краткая характеристика которых представлена в табл.1.
Таблица 1
|
№ сообщества |
Географическое положение |
Ассоциация |
ОПП (%) |
Обилие I. humilis |
|
Окинский район |
||||
|
3 |
Местность Синцын-Тала, левобережье, межтеррасный склон южной экспозиции |
Плотно дерновинная злаковая ирисовая степь (Avenula schelliana, Agrostis trinii) |
50 |
3 |
|
4 |
Е1адпойменная терраса р. Синца |
Овсецово-ирисовая степь (Avenula schelliana, Iris humilis) |
40 |
3 |
|
5 |
Лев. берег р. Синца, нижняя часть склона южной экспо зиции |
Злаково-полынная степь (Artemisia frigida, Artemisia commutata, Achnatherum sibiricum) |
20-30 |
+ |
|
Тункинский район |
||||
|
6 |
Окрестности с. Монды |
Разнотравно-стоповидноосоково-кобрезиевая степь (Роа botryoides, Cleistogenes squarrose, Carex duriuscula) |
50 |
2 |
|
Еравнинский район |
||||
|
1 |
Окрестности с. Гарам, к зап. от п. Сосново-Озерское, близ озера |
Остепненный луг (Carex obtusata, Poa botryoides, Myosotis imitate) |
70 |
1 |
|
2 |
К востоку от и. СосновоОзерское |
Леймусово-мятликовая степь (Leymus chinensis, Poa botryoides) |
70 |
1 |
Характеристика сообществ
Результаты и их обсуждение
Прегенеративный период онтогенеза I. humilis представлен проростками, ювенильными, имматурными и виргинильными особями.
Проростки представляют собой особи с 1-2 листьями длиной 2-2,7 см и шириной 0,1см, корневая система смешанная. Имеется главный корень и 1-2 придаточных, длиной 2-4 и соответственно 1-3. Ювенильные особи имеют 3-4 листья, развиты придаточные корни, которые расходятся от зоны кущения веером.
Имматурные особи несут на себе 1-2 мутовки с 2-Злистьями, длина которых составляет 4,5-9
см, ширина - 0,2 см. Корневище начинает ветвиться за счет образования боковых почек, из которых впоследствии образуются новые побеги.
Виргинильные особи за счет моноподиально-го нарастания корневищ образуют куст, состоящий из 3-5 вегетативных побегов, окутанных влагалищами прошлогодних листьев. На корневище закладываются почки возобновления, из которых в последующем формируются новые побеги.
Молодые генеративные растения несут как генеративные (1-2), так и вегетативные (5-10) побеги, которые состоят из 3-4 листьев длиной
-
3,5- 14.5 см. собранных в мутовки. На генеративном побеге формируется цветонос 5.5-13см длиной, на вершине которого супротивно отходят цветоножки, несущие каждый по одному цветку. После цветения и плодоношения генера
тивный побег отмирает и на корневище остается заметный рубец. Возобновление становится симподиальным в результате образования молодых корневищ в виде белых выростов с чешуевидными листьями.
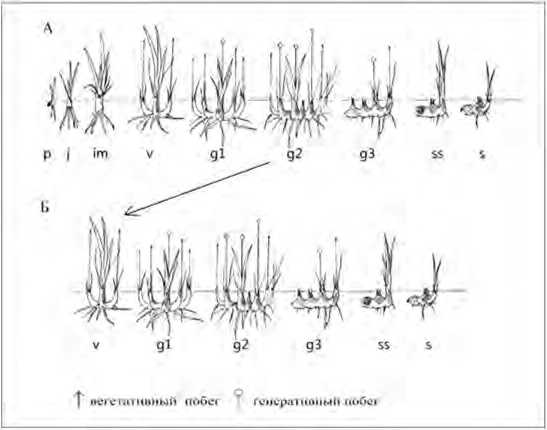
Рис.2 Онтогенез /ня humilis Georgi
А- полный онтогенез: Б - неполный онтогенез, р - проростки: j - ювенильные: im - иммап рное: v - виргинильные: g 1 - молодое генеративное: g2 - взрослое генеративное: g3 - старое генеративное; ss - субсешиплюе: s - сенильное.
У взрослых генеративных растений наблюдается максимальное количество генеративных (4-10) и вегетативных побегов (17-40). При этом границы особи становятся размытыми в результате годичного увеличения и разрастания корневища. В средневозрастном генеративном состоянии также наблюдается партикуляция особи (рис. 2). Отделение клона от материнской особи происходит в результате постепенного отмирания корневищ. Клон представляет собой парти-кулы, находящиеся в виргинильном или молодом генеративном состояниях. Старые генеративные особи представляют собой отдельные партикулы, состоящие из 1 генеративного и 2-3 вегетативных побегов. После потери способности к цветению растения переходят в постгснс-ративный период. Особи субсенильного и сенильного возраста состоят из 2-3 вегетативных побегов. несущих по 2-3 листа имматурного вида, корневище их полое и почти полностью разрушено.
На основе биологии вида, типа онтогенеза, особенностей размножения, длительности возрастных состояний характерный спектр ценопо-пуляций /. humilis можно определить как левосторонний. Данный тип спектра обусловлен тем. что ирисы преимущественно размножаются с помощью корневищ, нежели чем семенами |1|. в результате чего происходит омоложение особей генеративного состояния до виргинильных и молодых генеративных.
Изученные ЦП характеризуются левосторонним спектром, который совпадает с характерным. Абсолютный максимум приходится на виргинильные особи (33.9-55.4%) (табл. 2). что связано с омоложением до виргинильного состояния в результате партикуляции генеративных особей. Также достаточно значительна фракция молодых генеративных особей (13.2-35,4%).
ЦП /. humilis являются нормальными и в основном неполночленными. Так. в ЦП 5 и 6 отсутствует стадия проростка. Это связано с высокой степенью задернованности сообществ, что затрудняет прорастание семян ирисов. Также в этих ЦП отсутствуют взрослые генеративные особи. В ЦП 1 и 3 нс представлены особи постгенеративного периода (сенильные и субсенильные). Полночленную структуру имеют ЦП 3 и 4.
По классификации «дельта-омега» |4] восточно-саянские ценопопуляции, располагающиеся в Окинском (ЦП 1, 2, 3) и Тункинском (ЦП 4) районах, относятся к молодым за счет значительной фракции особей виргинильного и молодого генеративного возрастных состояний, а также отсутствия или незначительного количества (до 5,1%) особей постгенеративных возрастов. Ценопопуляций, расположенные на юге Витимского плоскогорья (ЦП 5,6), - переходные. Хотя доля виргинильных особей и превалирует, но также значительно количество особей субсенильных (20,7%) и сенильных (11,3%) воз растов. Экологическая плотность во всех ЦП колеблется от 5,3- 7,7 шт./м2, исключение составляет ЦП 4, где на 1м2 приходится 18,6 особи. Эффективная плотность изученных популяций значительно превышает экологическую плотность, что характерно для молодых популяций, где значительное количество приходится на молодую фракцию.
Таблица 2
|
№ ЦП |
онтогенетическое состояние особей, % от общего числа |
Демографические показатели |
тип ЦП |
|||||||||||
|
Р |
j |
im |
V |
G1 |
G2 |
G3 |
SS |
S |
E | 8 g |
к £ 1 E ■^ в В m |
A |
CD |
||
|
1 |
12,3 |
0,46 |
15 |
55,4 |
26,1 |
1,5 |
4,6 |
0 |
0 |
7,7 |
1,3 |
0,46 |
0,15 |
молодая |
|
2 |
3 |
6,15 |
1,5 |
55,3 |
20 |
4,6 |
1,5 |
4,6 |
3 |
6,5 |
2,5 |
0,48 |
0,22 |
молодая |
|
3 |
9,2 |
4,4 |
8,9 |
36,9 |
35,4 |
18,4 |
3,07 |
0 |
0 |
7,6 |
1,6 |
0,59 |
0,22 |
молодая |
|
4 |
3,4 |
15,5 |
15,5 |
31 |
17,2 |
1,7 |
8,6 |
5,1 |
1,7 |
18,6 |
3,7 |
0,42 |
0,22 |
молодая |
|
5 |
0 |
1,8 |
15 |
33,9 |
13,2 |
0 |
3,7 |
20,7 |
11,3 |
5,3 |
1,2 |
0,41 |
0,4 |
пере ходная |
|
6 |
0 |
1,8 |
13 |
45 |
13,2 |
0 |
3,7 |
20,7 |
11,3 |
5,8 |
1,4 |
0,45 |
0,41 |
пере ходная |
Примечание: А - индекс возрастности, со - индекс эффективности.
Онтогенетическая и демографическая характеристика Iris humilis
Заключение
В условиях Витимского плоскогорья и Восточного Саяна были изучены онтогенез и популяционная структура 6 ценопопуляций I. humilis.
Согласно концепции дискретного описания онтогенез I. humilis можно отнести ко II надтипу, где полный онтогенез осуществляется в ряду поколений вегетативно возникших особей [9]. В результате онтоморфогенеза I. humilis может проходить следующие стадии: отдельная особь —> первичный побег, первичный куст —> система парциальных побегов и кустов —> парциальный куст —> парциальный побег. Хотя для I. humilis и возможно семенное размножение, что подтверждается наличием в изученных популяциях особей ранних этапов онтогенеза, но преобладает вегетативное размножение за счет распадения материнских особей до омоложенных партикул. Преобладание вегетативного размножения определяет характер базового спектра, который имеет левосторонний характер за счет значительной фракции особей виргинильного состояния вегетативного происхождения. Небольшое влияние на соотношение возрастных групп в изученных ценопопуляциях, во-первых, оказывает задернованность сообществ, что отрицательно сказывается на прорастании семян: в еравнинских ценопопуляциях полностью отсутствуют проростки, а доля имматурных особей незначительна в отличие от восточно-саянских. Во-вторых, выпас скота способствует более быстрому распадению особей на раметы, вследствие чего в восточно-саянских популяциях отсутствуют особи постгенеративного периода, тогда как фракция сенильных и субсенильных особей составляет значительную долю в еравнинских. В целом экотопические условия и антропогенная нагрузка не сильно влияют на характер онтоген-тического спектра.
